ZmWRKY104–ZmCCaMK interaction enhances brassinosteroid-promoted salt tolerance in maize (Zea mays L.) via antioxidant defense
Li-li Zhao

TL;DR
This study identifies a new gene interaction in maize that improves salt tolerance through antioxidant defenses, which could help breed salt-resistant crops.
Contribution
A novel non-phosphorylation-dependent WRKY-CCaMK regulatory module is identified in the BR signaling pathway for salt tolerance in maize.
Findings
ZmWRKY104 interacts with ZmCCaMK in the nucleus to form a regulatory module.
Overexpression of ZmWRKY104 enhances antioxidant enzyme activities and salt tolerance in maize.
Transgenic maize with ZmWRKY104 shows reduced leaf yellowing and improved growth under salt stress.
Abstract
Brassinosteroid (BR)-mediated salt tolerance is a crucial mechanism for maize (Zea mays L.) adaptation to saline-alkaline environments. This study aimed to elucidate the molecular mechanism underlying BR-induced salt tolerance in maize, focusing on the regulatory roles of ZmWRKY104 and ZmCCaMK. Key results showed that ZmWRKY104 directly interacts with ZmCCaMK in the nucleus in a non-phosphorylation-dependent manner, forming a novel regulatory module. BR treatment upregulates ZmWRKY104 expression, and overexpression of ZmWRKY104 significantly enhances the activities of antioxidant enzymes (APX and SOD). Co-expression of ZmWRKY104 and ZmCCaMK synergistically promotes the antioxidant defense system in maize. Transgenic maize overexpressing ZmWRKY104 exhibits obvious salt tolerance advantages under 100 mM NaCl stress compared to wild-type plants, including reduced leaf yellowing, increased…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
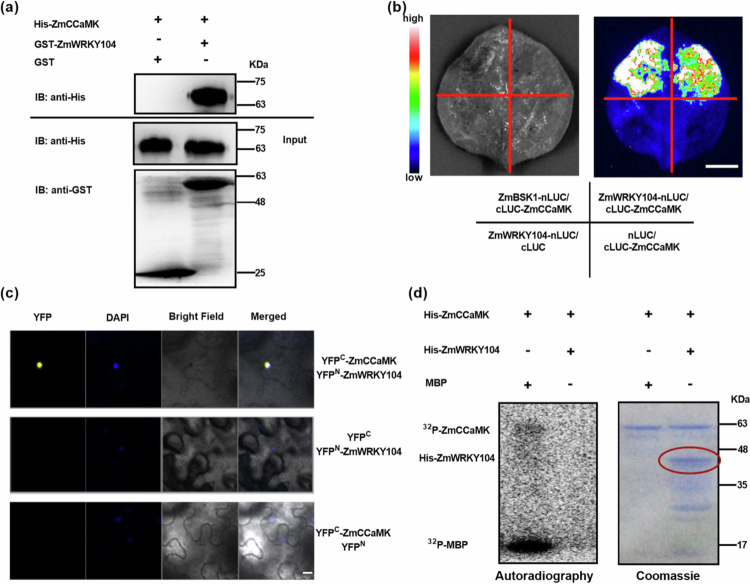 Figure 1
Figure 1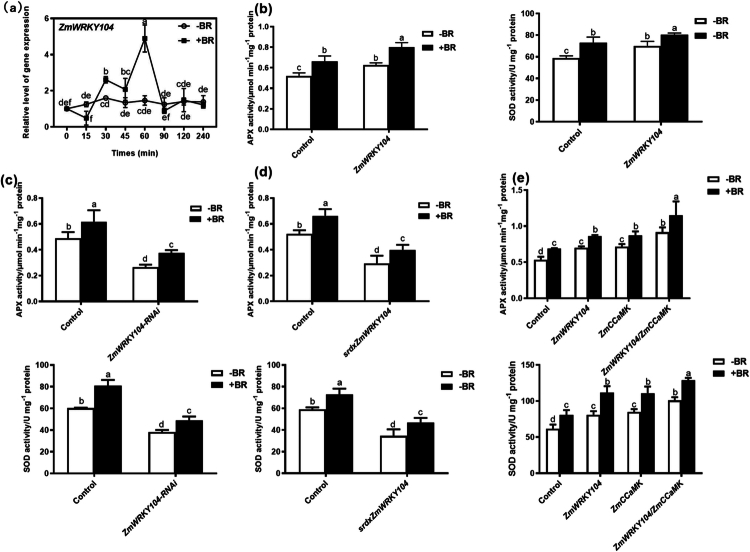 Figure 2
Figure 2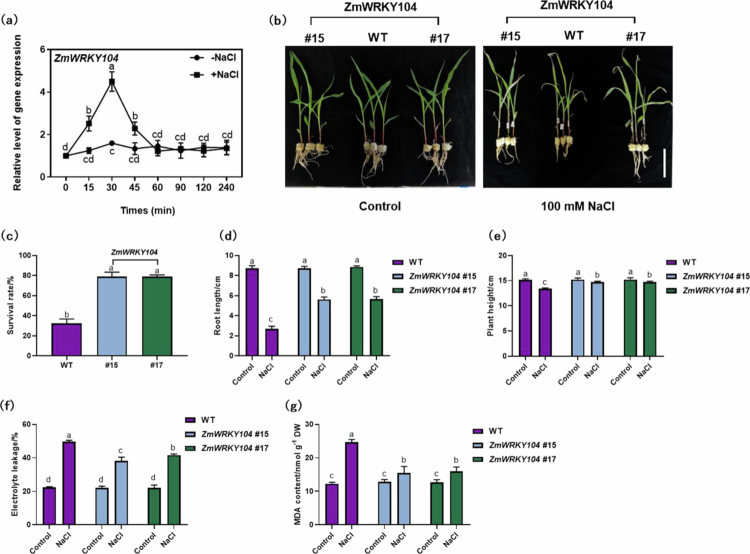 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Plant Stress Responses and Tolerance · Plant Molecular Biology Research
Introduction
1
Saline-alkaline land covers ~100 million hectares in China, with 7.6 million hectares of cultivated land affected by salinization, primarily in arid/semi-arid regions north of the Huaihe River, northwest China, and Xinjiang.1^,^2 Salt stress induces oxidative damage in maize via osmotic pressure and ion toxicity, reducing yields by 20–30%.3^,^4 Cultivating salt-tolerant varieties is critical for sustainable maize production in these regions.
Brassinosteroids (BRs) are key hormones regulating plant stress responses, improving abiotic tolerance by activating antioxidant systems.1^,^5 Calcium/calmodulin-dependent protein kinase (CCaMK) acts as a central signaling hub in abiotic stress,6^,^7 with synergistic regulation with BR emerging as a research focus.1^,^5 WRKY transcription factors are well-documented in plant stress defense,8‐16 but their interaction with CCaMK in BR-induced salt tolerance remains unreported in maize.
This study identified ZmWRKY104 as a ZmCCaMK-interacting protein, investigating its role in BR-mediated antioxidant defense and salt tolerance. The findings provide new insights into plant salt stress responses and valuable genetic resources for maize improvement in saline-alkaline regions.
Materials and methods
2
Plant materials and treatments
2.1
Maize (Zea mays L. cv. B73) and Nicotiana benthamiana seeds were maintained by the laboratory. Two-week-old maize seedlings were treated with 50 nmol/L BR (oleuropein lactone, Sigma-Aldrich, USA) or 100 mM NaCl (Solarbio, China) for 0, 15, 30, 45, 60, 90, 120, or 240 min; controls were treated with distilled water. The second leaf was harvested, frozen in liquid nitrogen, and stored at −80 °C.17
Protein interaction assays
2.2
GST-pull down**:** Purified GST-ZmWRKY104 and His-ZmCCaMK proteins (expressed in E. coli BL21) were incubated with GST affinity resin (GE Healthcare, USA) at 4 °C for 2 h. Bound proteins were detected by Western blotting using anti-His (Abcam, ab18184) and anti-GST (Abcam, ab19256) antibodies.17
LCI assay**:** ZmWRKY104 was cloned into pC1300-nLUC, and ZmCCaMK into pC1300-cLUC. The constructs were co-transformed into N. benthamiana leaves via Agrobacterium GV3101. Luciferase signals were detected using an in vivo imaging system (Tanon-5200, China) after 48 h.18
BiFC assay**:** ZmWRKY104 (CDS without terminator) was cloned into pSPYNE (YFP^N^-ZmWRKY104), and ZmCCaMK into pSPYCE (YFP^C^-ZmCCaMK). The vectors were co-infiltrated into 4-week-old N. benthamiana leaves, and YFP signals were observed via confocal microscope (Zeiss LSM710, Germany).17^,^19
In Vitro kinase assay
2.3
The reaction system (50 μL) contained 25 mM Tris-HCl (pH 7.5), 5 mM MgCl₂, 1 mM DTT, 2.5 mM CaCl₂, 2 μM calmodulin (CaM), 200 nmol/L ATP, 10 μCi [γ- ³²P] ATP, 0.01 mg/mL ZmCCaMK, and 1 μg His-ZmWRKY104 (or MBP as positive control). After incubation at 30 °C for 30 min, products were separated by 12% SDS-PAGE and detected by radioautography.17
Vector construction
2.4
The pXZP008 vector, which harbors the mCherry gene driven by the ubiquitin promoter, was used for protoplast transient expression assays as described previously.17 The coding region of ZmWRKY104 was amplified by PCR and cloned into the pXZP008 vector using BamHI and KpnI restriction enzymes, generating the ubi:ZmWRKY104-mCherry plasmid. To construct the dominant-negative ubi:ZmWRKY104-SRDX-mCherry plasmid, the exogenous EAR motif repression domain SRDX (5’-CTGGATCTAGAACTCCGTTTG-3’) was introduced into the C-terminus of ZmWRKY104 via PCR, following the method described by.20
Antioxidant enzyme activity detection
2.5
Maize protoplasts were isolated from B73 leaves (Sheen laboratory protocol, modified). Protoplasts were transfected with ZmWRKY104 overexpression (pXZP008-ZmWRKY104)/SRDX vector (pXZP008-ZmWRKY104-SRDX) or dsRNA (for silencing) via PEG-mediated method. After 24 h, APX and SOD activities were measured using kits (Solarbio, China).5,19‐24
Transgenic maize salt tolerance evaluation
2.6
ZmWRKY104 overexpression lines (#15, #17) and wild-type (WT) maize were grown in 1/2 Hoagland solution. At the 3-leaf stage, seedlings were treated with 100 mM NaCl for 4 days. Phenotypes (leaf yellowing, plant height, root length) were recorded, and electrolyte leakage (EL) and malondialdehyde (MDA) content were measured.17
Statistical analysis
2.7
Data were analyzed using GraphPad Prism 8.2.1. Significant differences were determined by Duncan’s multiple comparisons (p < 0.05). All experiments had ≥3 biological replicates.
Results and discussion
3
ZmWRKY104 directly interacts with ZmCCaMK in
Vitro and in Vivo
3.1
GST-pull down assay showed GST-ZmWRKY104 bound to His-ZmCCaMK, while the GST control did not (Figure 1a). LCI assay revealed strong luciferase signals in N. benthamiana leaves co-transformed with ZmWRKY104-nLUC +cLUC-ZmCCaMK, significantly higher than negative controls (Figure 1b). BiFC assay detected YFP signals in the nucleus of leaf cells co-expressing YFP^C^-ZmCCaMK and YFP^N^-ZmWRKY104 (Figure 1c). These results confirm a direct nuclear interaction between ZmWRKY104 and ZmCCaMK— the first report of WRKY-CCaMK crosstalk in maize BR signaling. This nuclear localization aligns with the transcriptional regulatory role of WRKY proteins and the signaling function of CCaMK, suggesting they may co-regulate downstream gene expression.7^,^25
ZmWRKY104 directly interacts with ZmCCaMK in a non-phosphorylation-dependent manner. (a) GST-pull down assay showing binding between GST-ZmWRKY104 and His-ZmCCaMK. GST/His-ZmCCaMK was used as a negative control. (b) LCI assay showing strong luciferase signals in co-transformed N. benthamiana leaves. ZmBSK1-nLUC/cLUC-ZmCCaMK was used as a positive control, and ZmWRKY104-nLUC/cLUC and nLUC/cLUC-ZmCCaMK were used as negative controls. Scale bar, 1 cm. (c) BiFC assay showing nuclear YFP signals in co-expressing leaf cells (DAPI = nuclear stain). Scale bar, 20 µm. (d) In vitro kinase assay showing no ³²P-labeled ZmWRKY104 (MBP = positive control). The ZmWRKY104 band is located within the red ellipse. Data are means ± SE (n = 3, p < 0.05).
ZmCCaMK does not phosphorylate ZmWRKY104
3.2
CCaMK was initially identified as a key factor in root nodule symbiosis of legumes and arbuscular mycorrhizal symbiosis of mycorrhizal plants, and its role in stress response was subsequently confirmed. Previous studies have reported that CCaMK can phosphorylate its target factor CYCLOPS; however, the present study found a novel interaction between ZmCCaMK and ZmWRKY104 that is independent of phosphorylation.
In vitro kinase assays showed a ³²P-labeled band for MBP (positive control) but no ³²P-labeled band for ZmWRKY104 (Figure 1d), indicating that ZmWRKY104 is not a phosphorylation substrate of ZmCCaMK.
To highlight the innovation of this finding, we compared the interaction mode of CCaMK/CYCLOPS with that of ZmCCaMK/ZmWRKY104 based on relevant literature26^,^ 27), with the following differences: 1) Binding mode: The interaction between CCaMK and CYCLOPS is phosphorylation-dependent (CCaMK phosphorylates the Ser/Thr sites of CYCLOPS), which is a classic mechanism of root nodule symbiotic signal transduction in legumes. In contrast, the interaction between ZmCCaMK and ZmWRKY104 in this study is phosphorylation-independent (no phosphorylation band of ZmWRKY104 was detected in in vitro kinase assay), representing a novel non-phosphorylation-dependent interaction. 2) Functional localization: CCaMK/CYCLOPS mainly regulates the development of symbiotic organs, while ZmCCaMK/ZmWRKY104 focuses on the regulation of antioxidant defense under abiotic stress (salt stress). 3) Action mode: CCaMK activates the transcriptional function of CYCLOPS through phosphorylation, whereas ZmCCaMK may act as a “scaffold protein” to recruit ZmWRKY104 to the promoter regions of APX and SOD genes, and can activate downstream genes without phosphorylation. This comparison clearly highlights the innovative discovery of this study in the functional diversity of CCaMK.
Additionally,1 focused on drought stress and found that the autophosphorylation sites (Thr420 and Ser454) of ZmCCaMK are the key to BR-induced antioxidant defense. In contrast, the present study targets salt stress and reveals that ZmCCaMK regulates antioxidant defense through non-phosphorylation interaction with ZmWRKY104. This indicates that ZmCCaMK adopts a differentiated regulatory mechanism under different stress conditions: it directly activates the downstream pathway through autophosphorylation under drought stress, and regulates the pathway by forming a complex through protein interaction under salt stress, reflecting the diversity and specificity of the plant stress signal network.
ZmWRKY104 positively regulates BR-induced antioxidant defense
3.3
qRT-PCR showed ZmWRKY104 expression was upregulated by BR treatment, peaking at 60 min (4.8-fold vs. 0 h, Figure 2a), indicating ZmWRKY104 is a BR-responsive gene. Transient overexpression of ZmWRKY104 in protoplasts increased APX (20%) and SOD (19%) activities, with BR treatment further enhancing these activities (APX: +28%, SOD: +15%, Figure 2b). Silencing ZmWRKY104 via dsRNA or srdxZmWRKY104 reduced basal APX/SOD activities by 36–45%, and BR could not restore these activities (Figure 2c-d). These results demonstrate ZmWRKY104 is essential for BR-induced antioxidant defense— consistent with BR’s role in activating antioxidant systems to mitigate salt-induced oxidative damage.1^,^5
ZmWRKY104 regulates BR-induced antioxidant defense, synergistically enhanced by ZmCCaMK. (a) qRT-PCR showing ZmWRKY104 expression under BR treatment. White circles represent distilled water treatment, and black squares represent 50 nmol/L BR treatment. (b) Transient overexpression of ZmWRKY104 increases APX/SOD activities. (c-d) Silencing/SRDX ZmWRKY104 reduces basal antioxidant enzyme activities, which BR cannot restore. (e) Co-expression of ZmWRKY104 and ZmCCaMK synergistically enhances APX/SOD activities. Different lowercase letters indicate significant differences (n = 3, p < 0.05).
ZmWRKY104 and ZmCCaMK synergistically enhance BR-induced antioxidant defense
3.4
Co-expression of ZmWRKY104 and ZmCCaMK in protoplasts increased APX (71%) and SOD (63%) activities— higher than single-gene expression (ZmWRKY104 alone: +31% APX, +31% SOD; ZmCCaMK alone: +34% APX, +37% SOD, Figure 2e). BR treatment further elevated enzyme activities in the co-expression group (APX: +25%, SOD: +38%). This synergism suggests ZmCCaMK interacts with ZmWRKY104 to amplify BR-mediated antioxidant responses, potentially by stabilizing ZmWRKY104 or facilitating its binding to target gene promoters. The findings highlight the importance of the ZmWRKY104-ZmCCaMK module in fine-tuning BR-induced stress defense, which is critical for maize adapting to salt stress.
ZmWRKY104 overexpression enhances maize salt tolerance
3.5
NaCl treatment upregulated ZmWRKY104 expression (2.8-fold at 30 min, Figure 3a), indicating its involvement in salt stress responses. Under 100 mM NaCl, WT maize showed severe leaf yellowing (60–70% yellow leaves), reduced plant height (11% vs. control), and low survival rate (32%); overexpression lines (#15, #17) had less yellowing (20–25%), higher plant height (97% vs. control), and survival rate (79%, Figure 3b–e). Additionally, EL (40% vs. 50% in WT) and MDA (16 nmol/g FW vs. 25 nmol/g FW in WT) were lower in overexpression lines (Figure 3f-g). EL and MDA are indicators of membrane damage caused by oxidative stress, so their reduction confirms ZmWRKY104 enhances salt tolerance by mitigating oxidative damage via antioxidant defense activation.
ZmWRKY104 overexpression enhances maize salt tolerance. (a) qRT-PCR showing ZmWRKY104 expression under NaCl treatment. (b) Phenotypes of WT and overexpression lines (#15, #17) under salt stress. Maize seedlings were treated with 100 mM NaCl for 4 days, with ~30 transgenic plants per replicate; representative images were captured at 4 days post-treatment. Scale bar, 5 cm. (c–e) Survival rate (Maize seedlings were treated with 100 mM NaCl for 7 d, ~30 plants per replicate), root length, and plant height of maize seedlings. (f-g) EL and MDA content in leaves under salt stress. Different lowercase letters indicate significant differences (n = 3, p < 0.05).
This practical significance is particularly relevant for northern China’s saline-alkaline regions, where maize yield is constrained by 100–150 mM NaCl. Introducing ZmWRKY104 into local varieties (e.g., “Zhengdan 958”) via CRISPR/Cas9 could increase yield, addressing the regional challenge of saline-alkaline land utilization.
Conclusions
4
- ZmWRKY104 directly interacts with ZmCCaMK in the nucleus via a non-phosphorylation-dependent mechanism.
- ZmWRKY104 positively regulates BR-induced antioxidant defense (APX/SOD activities), with synergistic enhancement by ZmCCaMK.
- Overexpression of ZmWRKY104 improves maize salt tolerance by reducing oxidative damage (lower EL and MDA).
ZmWRKY104 is a promising candidate gene for salt-tolerant maize breeding, providing a practical strategy for sustainable agriculture in saline-alkaline regions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Liu L, Han T, Liu W, Di P, Yu X, Yan J, Zhang A. Thr 420 and Ser 454 of Zm C Ca MK play a crucial role in brassinosteroid-induced antioxidant defense in maize. Biochem Biophys Res Commun. 2020;525(3):537–542. doi: 10.1016/j.bbrc.2020.03.025.32113680 · doi ↗ · pubmed ↗
- 2Zhou N. Saline-alkaline land resources in China and their utilization. J Soil Water Conserv. 2012;26(4):1–6.
- 3Sun Y, Wang KJ, Zhang JW, Liu P, Dong ST. Effects of salt stress on maize growth and yield. J Maize Sci. 2016;24(3):123–128.
- 4Zhang H, Dong S. Salt stress tolerance mechanisms in maize. Chin J Appl Ecol. 2011;22(5):1345–1352.
- 5Yan J, Guan L, Sun Y, Nakano T, Kato H, Shima Y, Ito Y. Calcium and Zm C Ca MK are involved in brassinosteroid-induced antioxidant defense in maize leaves. Plant Cell Physiol. 2015;56(5):883–896. doi: 10.1093/pcp/pcv 034.25647327 · doi ↗ · pubmed ↗
- 6Defalco TA, Bender KW, Snedden WA. Breaking the code: Ca²⁺ sensors in plant signalling. Biochem J. 2009;425(1):27–40. doi: 10.1042/BJ 20081838.20001960 · doi ↗ · pubmed ↗
- 7Poovaiah BW, Du L, Wang H, Yang T. Recent advances in calcium/calmodulin-mediated signaling with an emphasis on plant-microbe interactions. Plant Physiol. 2013;163(2):531–542. doi: 10.1104/pp.113.219778.24014576 PMC 3793035 · doi ↗ · pubmed ↗
- 8Cai R, Dai W, Zhang C, Wang Y, Wu M, Zhao Y, Ma Q, Xiang Y, Cheng B. The maize WRKY transcription factor Zm WRKY 17 negatively regulates salt stress tolerance in transgenic Arabidopsis plants. Planta. 2017;246(6):1215–1231. doi: 10.1007/s 00425-017-2744-6.28861611 · doi ↗ · pubmed ↗