Phylogenomic analysis and genetic mechanisms of antifungal resistance in clinical isolates of Candida glabrata (Nakaseomyces glabratus) from across Canada, 2013–2020
Domenica G. De Luca, David C. Alexander, Tanis C. Dingle, Philippe J. Dufresne, Jeff Fuller, Greg J. German, David Haldane, Linda M. Hoang, Lei Jiao, Julianne V. Kus, Lisa Li, Kathy Malejczyk, Caroline Sheitoyan-Pesant, Markus Stein, Morag Graham, Gary Van Domselaar

TL;DR
This study uses whole-genome sequencing to explore how Candida glabrata strains in Canada develop resistance to antifungal drugs and how they are related genetically.
Contribution
The study is the largest national genomic survey of clinical C. glabrata isolates in Canada and identifies novel genetic variants linked to antifungal resistance.
Findings
Phylogenomic analysis revealed 15 genetically related clusters of C. glabrata, with one cluster significantly associated with antifungal resistance.
Thirty-six unique PDR1 variants were found in fluconazole-resistant isolates, with over half being novel resistance variants.
Most micafungin-resistant isolates had well-characterized FKS gene mutations, indicating a known resistance mechanism.
Abstract
Candida glabrata (Nakaseomyces glabratus) is an important cause of invasive fungal infections and may exhibit reduced susceptibility toward antifungal drugs. Here, we used whole-genome sequencing to investigate the genomic phylogeny and identify genetic determinants of antifungal resistance in a collection of 142 clinical C. glabrata isolates obtained from the 10 provinces of Canada between 2013 and 2020. Our study prioritized resistant isolates (n = 62, 43.7%) from invasive infections to better understand antifungal resistance and represents the largest national genomic survey of clinical C. glabrata isolates performed to date in Canada. Phylogenomic analysis based on single-nucleotide variants in the core genome revealed 15 genetically related clusters of C. glabrata. The largest cluster (cluster I, n = 29) was significantly associated with antifungal resistance (P value = 0.0112).…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
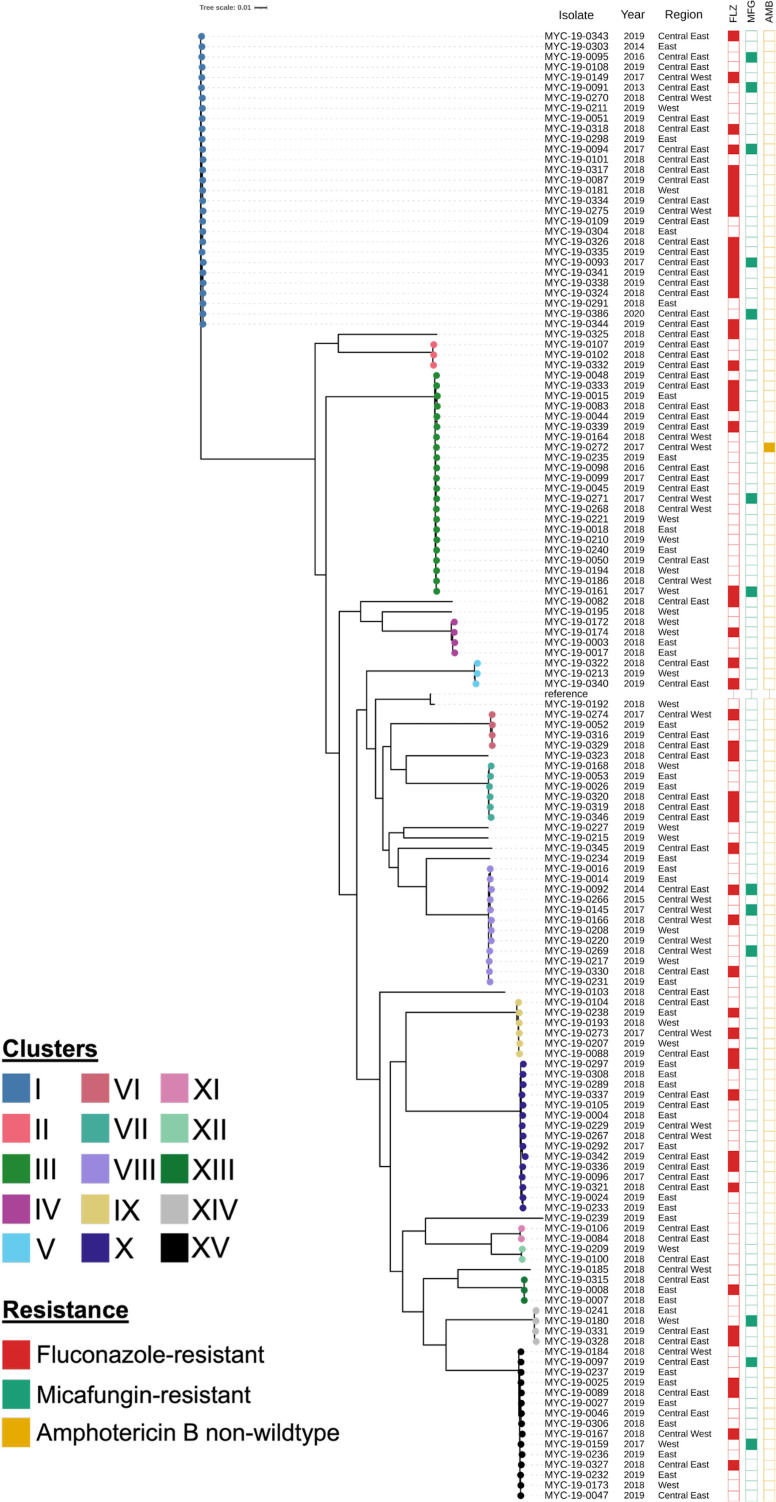 Fig 1
Fig 1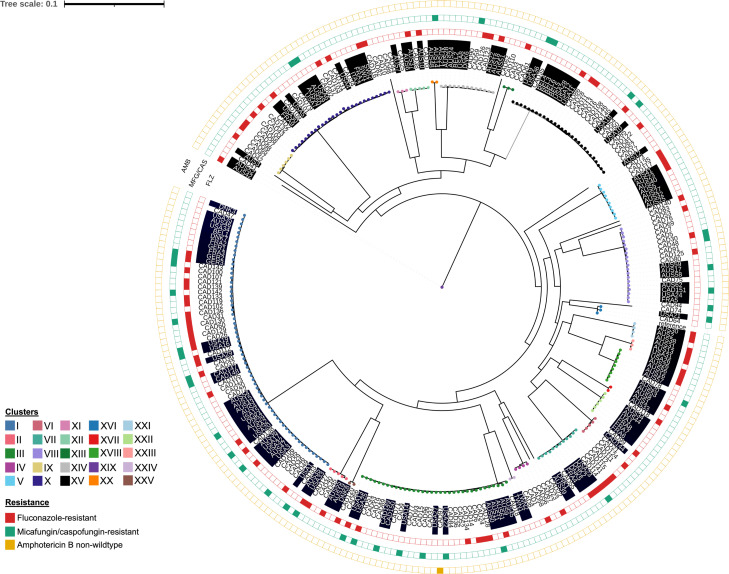 Fig 2
Fig 2| Antifungal drug | MIC (µg/mL) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤0.008 | 0.016 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | >128 | % S/SDD | % I | % R/NWT | |
| FLZ | NA | NA | NA | NA | NA | NA | 1 | 2 | 4 | 24 | 42 | 13 | 4 |
|
|
| 63.4 | NA | 36.6 |
| MFG | 49 | 56 | 13 | 7 | 4 |
|
|
|
|
|
| NA | NA | NA | NA | NA | 88.0 | 2.8 | 9.2 |
| AMB | NA | NA | NA | NA | 4 | 19 | 59 | 46 | 13 |
| NA | NA | NA | NA | NA | NA | 99.3 | NA | 0.7 |
| Cluster ( | S/SDD | R/NWT | |
|---|---|---|---|
| I (29) | 10 (34.5) | 19 (65.5) |
|
| II (3) | 2 (66.7) | 1 (33.3) | 1.0000 |
| III (22) | 15 (68.2) | 7 (31.8) | 0.2509 |
| IV (4) | 3 (75.0) | 1 (25.0) | 0.6319 |
| V (3) | 1 (33.3) | 2 (66.7) | 0.5806 |
| VI (4) | 2 (50.0) | 2 (50.0) | 1.0000 |
| VII (6) | 3 (50.0) | 3 (50.0) | 1.0000 |
| VIII (12) | 7 (58.3) | 5 (41.7) | 1.0000 |
| IX (6) | 3 (50.0) | 3 (50.0) | 1.0000 |
| X (15) | 10 (66.7) | 5 (33.3) | 0.4263 |
| XI (2) | 2 (100.0) | 0 (0.0) | 0.5045 |
| XII (2) | 2 (100.0) | 0 (0.0) | 0.5045 |
| XIII (3) | 2 (66.7) | 1 (33.3) | 1.0000 |
| XIV (4) | 1 (25.0) | 3 (75.0) | 0.3182 |
| XV (15) | 9 (60.0) | 6 (40.0) | 0.7916 |
| Cluster | Isolate | Region | FLZ MIC (µg/mL) | Confirmed | Suspected | Potential novel |
|---|---|---|---|---|---|---|
| I | MYC-19-0087 | Central East | 128 | G583S ( | ||
| MYC-19-0093 | Central East | 128 | K1087E | |||
| MYC-19-0181 | West | 128 | G583S ( | |||
| MYC-19-0275 | Central West | 128 | S343P | |||
| MYC-19-0317 | Central East | 64 | G583S ( | |||
| MYC-19-0318 | Central East | 64 | E340K ( | |||
| MYC-19-0324 | Central East | 64 | D1089Y ( | |||
| MYC-19-0326 | Central East | 64 | G1099D ( | |||
| MYC-19-0334 | Central East | 128 | G583S ( | |||
| MYC-19-0335 | Central East | >128 | A693E | |||
| MYC-19-0338 | Central East | 128 | L291F | |||
| MYC-19-0341 | Central East | >128 | L931F | |||
| MYC-19-0343 | Central East | 128 | C350R | |||
| II | MYC-19-0332 | Central East | 128 | S942F ( | ||
| III | MYC-19-0015 | East | 64 | N764I | ||
| MYC-19-0083 | Central East | 128 | N764I | |||
| MYC-19-0161 | West | >128 | E1083G | |||
| MYC-19-0339 | Central East | 128 | G346D ( | |||
| IV | MYC-19-0174 | West | 64 | S316I ( | ||
| V | MYC-19-0340 | Central East | 64 | T360I ( | ||
| VI | MYC-19-0274 | Central West | 64 | P235L | ||
| VII | MYC-19-0319 | Central East | 128 | D876Y ( | ||
| MYC-19-0320 | Central East | 128 | D876N ( | |||
| MYC-19-0346 | Central East | 64 | R761S ( | |||
| VIII | MYC-19-0092 | Central East | ≥256 | I616F | ||
| MYC-19-0166 | Central West | 128 | W256R | |||
| MYC-19-0330 | Central East | 64 | Y124S | |||
| IX | MYC-19-0088 | Central East | ≥256 | R592S ( | ||
| MYC-19-0238 | East | 64 | I207T | |||
| MYC-19-0273 | Central West | 128 | D876N ( | G611V | ||
| X | MYC-19-0297 | East | 128 | D876N ( | ||
| MYC-19-0336 | Central East | 128 | K274E ( | F613del | ||
| MYC-19-0342 | Central East | 64 | T1080A ( | |||
| XV | MYC-19-0025 | East | 64 | R761G | ||
| MYC-19-0089 | Central East | 256 | V849L | |||
| – | MYC-19-0082 | Central East | 128 | R772K ( | I971V | |
| – | MYC-19-0325 | Central East | 64 | L1081F | ||
| – | MYC-19-0345 | Central East | 64 | G1079R ( | G189V |
| Cluster | Isolate | Region | MFG MIC (µg/mL) | |||
|---|---|---|---|---|---|---|
| I | MYC-19-0091 | Central East | 0.25 | WT | WT | WT |
| MYC-19-0093 | Central East | 2 | WT |
| WT | |
| MYC-19-0094 | Central East | 8 | WT |
| WT | |
| MYC-19-0095 | Central East | 4 | WT |
| WT | |
| MYC-19-0386 | Central East | 1 | WT |
| WT | |
| III | MYC-19-0161 | West | 0.25 | D632V | WT | WT |
| MYC-19-0271 | Central West | 1 | WT |
| WT | |
| MYC-19-0272 | Central West | 0.12 (I) | WT | WT | WT | |
| VIII | MYC-19-0092 | Central East | 4 |
| WT | WT |
| MYC-19-0145 | Central West | 1 | WT |
| WT | |
| MYC-19-0266 | Central West | 0.12 (I) | WT | WT | WT | |
| MYC-19-0269 | Central West | 0.5 | WT |
| WT | |
| XIII | MYC-19-0315 | Central East | 0.12 (I) | WT | WT | R1378S |
| MYC-19-0008 | East | 0.12 (I) | WT | WT | WT | |
| XIV | MYC-19-0180 | West | 2 | WT |
| WT |
| XV | MYC-19-0097 | Central East | 2 | WT |
| WT |
| MYC-19-0159 | West | 2 | WT |
| WT |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Fungal Infections and Studies · Nail Diseases and Treatments
INTRODUCTION
The yeast Candida glabrata, also known as Nakaseomyces glabratus, is an opportunistic human fungal pathogen that may cause potentially life-threatening invasive infections. As a commensal member of the human gastrointestinal microbiome, systemic infections of C. glabrata most often originate from the intestinal microflora of the host (1, 2), rather than dissemination of clones (3–5). C. glabrata has become a recognized nosocomial pathogen with increasing incidence of invasive disease (3), which is of particular concern since fungal infections by this species are amongst those most often associated with antifungal resistance (6, 7). In many countries worldwide, C. glabrata is a leading cause of invasive candidiasis, typically ranking second to Candida albicans (7–12). Unlike most Candida species, C. glabrata displays reduced susceptibility toward azole antifungals, especially fluconazole, and can acquire azole and echinocandin resistance following antifungal therapy (6, 7, 13). Resistance has been reported within all three drug classes in C. glabrata and some strains exhibit multidrug resistance (i.e., resistance to two or more drug classes) (7). With only three classes of antifungal drugs (azoles, echinocandins, polyenes) available for the management of invasive infections, resistance to even a single class may severely limit treatment options.
Azole resistance in C. glabrata (e.g., fluconazole, itraconazole, posaconazole, and voriconazole) is primarily mediated by amino acid substitutions in PDR1, the central transcriptional regulator of efflux pump expression. Activated forms of PDR1 induce high levels of azole resistance by overexpressing the ATP-binding cassette (ABC) transporters CDR1, PDH1 (CDR2), and SNQ2 (14, 15). Specific regions of PDR1, referred to as the inhibitory domain (ID; 312–382 aa), middle homology region (MHR; 539–632 aa), and transcriptional activation domain (TAD; 800–1107 aa), are predictive of azole resistance (16). Alterations in ERG11, mitochondrial dysfunction, chromosomal duplications, and ploidy variations have also been reported as minor potential contributors to azole resistance in C. glabrata (17–20). Echinocandin resistance (e.g., anidulafungin, caspofungin, and micafungin) in C. glabrata is typically mediated by alterations in FKS subunits of glucan synthase. Specific positions and amino acid variations in highly conserved hot-spot (HS) regions of FKS1 and FKS2 genes are correlated with reduced echinocandin susceptibility (21). The mechanisms of polyene (e.g., amphotericin B) resistance in C. glabrata are not well understood; however, genes involved in ergosterol biosynthesis (e.g., ERG2, ERG3, and ERG6) have been implicated (22–24).
Based on a large Canadian multicenter study conducted between 2011 and 2016, C. glabrata was the second most common Candida species isolated from invasive infections, with resistance rates of 1.0% and 2.5% for fluconazole and micafungin, respectively (8). A study from a large reference laboratory in Ontario, Canada, reported a higher rate of fluconazole resistance (9.0%) during a similar period (25). Although the rates of antifungal resistance in Canada are relatively low, limited information is known beyond the species distribution and phenotypic antifungal susceptibilities of these infections. Several molecular typing methods, such as multi-locus sequence typing (MLST) and microsatellite length polymorphism typing, are widely used to analyze genetic variation in C. glabrata; however, whole-genome sequencing provides higher resolution and superior discriminatory power compared to these methods (26).
In this study, we sequenced 142 clinical strains of C. glabrata isolated between 2013 and 2020 from the 10 provinces of Canada, representing the largest and most comprehensive genomic study of C. glabrata conducted to date in Canada. Invasive infections resistant to at least one antifungal drug from the three major classes (azoles, echinocandins, polyenes) were prioritized to improve insights into antifungal resistance. As such, this collection was not intended to reflect the current rates of antifungal resistance observed in Canada, but rather to investigate antifungal-resistant strains of C. glabrata in terms of the genomic phylogeny and associated genetic mechanisms of antifungal resistance.
MATERIALS AND METHODS
Isolate collection
The 10 provincial public health laboratories in Canada provided 142 clinical C. glabrata isolates collected between 2013 and 2020. We prioritized isolates from invasive infections that were resistant to at least one antifungal drug from the three major drug classes (azoles, echinocandins, and polyenes) to enrich our collection with antifungal-resistant isolates. Twenty-two isolates were from Western Canada (British Columbia or Alberta), 20 from Central West (Saskatchewan or Manitoba), 65 from Central East (Ontario or Québec), and 35 from Eastern Canada (New Brunswick, Newfoundland, Nova Scotia, or Prince Edward Island). Isolates were recovered from sterile and non-sterile body sites, including blood (n = 85), urine (n = 11), intra-abdominal (n = 7), pus (n = 7), kidney (n = 2), abscess fluid (n = 2), bile (n = 2), pancreas (n = 2), and 24 isolates from various other sites. The line list of isolates used in this study, geographic region of isolation, antifungal minimum inhibitory concentrations (MICs), year, and site of sampling are presented in Table S1.
For the global analysis, we included an additional 139 C. glabrata strains with antifungal susceptibility data from the National Center for Biotechnology Information (NCBI). Isolates were from Australia (n = 50, BioProject PRJNA480138; n = 12, PRJNA310957), Belgium (n = 6, PRJNA361477), Canada (n = 9, PRJNA610214), Denmark (n = 8, PRJEB40738), France (n = 9, PRJNA361477), Germany (n = 1, PRJNA361477; n = 2, PRJNA483064), Italy (n = 2, PRJNA361477), Switzerland (n = 2, PRJNA374542), Taiwan (n = 2, PRJNA361477), and the United States (n = 20, PRJNA592373; n = 12, PRJNA361477; n = 3, PRJNA524686; n = 1, PRJNA329124). Sequence Read Archive accessions with corresponding BioProject numbers are listed in Table S2.
Antifungal susceptibility testing
Antifungal susceptibility testing was carried out by broth microdilution either following the Clinical and Laboratory Standards Institute (CLSI) M27-Ed4 guidelines (27) with custom YCML1FCFA panels (Thermo Fisher Oxoid, Oakwood Village, OH) or the manufacturer’s instructions for Sensititre YeastOne Y09 panels (Thermo Oxoid, Oakwood Village, OH). Results for fluconazole, micafungin, and amphotericin B were reported in this study. MICs were evaluated at 24 h and interpreted either using clinical breakpoints established in the CLSI M27M44S Ed3 document (28) (fluconazole and micafungin) or epidemiological cutoff values established in the CLSI M57S Ed4 document (29) (amphotericin B). Resistance breakpoints for fluconazole and micafungin resistance were MICs of ≥64 µg/mL and ≥0.25 µg/mL, respectively. The breakpoint for an intermediate MIC to micafungin was 0.12 µg/mL. The epidemiological cutoff value used for amphotericin B non-wild type was >2 µg/mL. Candida krusei ATCC 6258 and Candida parapsilosis ATCC 22019 were used as quality control strains.
Whole-genome sequencing
Genomic DNA (gDNA) was extracted using the MasterPure Complete DNA and RNA Purification Kit (Mandel, Guelph, ON). Libraries were prepared using the Nextera XT DNA Library Preparation kit (Illumina, San Diego, CA) and sequencing was carried out on the NextSeq 500/550 platform (Illumina, San Diego, CA) with the NextSeq 500/550 v2.5 kit (Illumina, San Diego, CA). Isolates with coverage below 60× were re-sequenced.
Genomic analysis
Raw reads were pre-processed using fastp v.0.23.2 with default settings (30). Phylogenomic analyses based on single-nucleotide variants (SNVs) in the core genome were carried out using the Single Nucleotide Variant Phylogenomic v.1.2.3 pipeline (31). Parameters used were minimum coverage 10, minimum mapping quality 30, SNV abundance ratio 0.75, and SNV density filter >2 SNVs per 20-base window. C. glabrata CBS 138 (ATCC 2001) was used as the reference genome (32). Briefly, repeat regions along the reference genome were masked using MUMmer, processed paired-end reads were aligned to the reference genome using SMALT, and SNVs were called by two independent variant callers (FreeBayes and SAMtools/BCFtools). Variant and non-variant calls were merged into a single file and flagged for mismatches between variant callers. Base calls that did not meet the defined thresholds for coverage, mapping quality, and SNV abundance were removed. High-density regions were identified to mask potential recombinant regions. PhyML was used to construct a maximum-likelihood phylogeny based on the core genome SNV alignment using the GTR + γ substitution model. The SNV alignment was also used to generate a pairwise SNV distance matrix. Maximum-likelihood dendrograms were visualized and annotated using the Interactive Tree of Life v.7.2 (33).
For the variant analysis, the processed reads were aligned using BWA-mem v.0.7.17.1 (34) and sorted using SAMtools v.1.18 (35). Variants were called using freebayes v.1.3.7 (minimum mapping quality: 30, minimum coverage: 10, minimum alternate allele fraction: 0.75, minimum base quality: 20) (36). snpEff v.5.2 (37) was used to annotate variants and snpSIFT v.5.2 (38) was applied to extract the resistance genes of interest.
Copy number variants (CNVs) were identified using a read-depth approach. GC bias was corrected using deepTools computeGCbias v.3.3.2.0.0 and correctGCbias v.3.3.2.0.0 (39). The reference genome was divided into 1,000 base pair windows using BEDTools makewindows v.2.31.1 (40). Read coverage across the genome was calculated from aligned and sorted BAM files using BEDTools genomecov v.2.31.1 (40). The average read depth per window was then computed using bedtools map v.2.31.1 (40). Chromosome-end bias was corrected based on the average read depth versus distance to the nearest chromosome end using a custom R script for locally weighted scatterplot smoothing regression (41). Coverage values were normalized by dividing by the genome-wide median coverage. Genes with normalized coverage values ≥2 or 0 were considered to have increased or decreased copy numbers, respectively. CNVs were subsequently identified, annotated, and visualized using a custom R script (41).
Statistical analysis
The Fisher’s exact test was used to determine if there were statistically significant associations between clusters and categorical variables, such as region of isolation, body site of isolation, and antifungal resistance. P values of <0.05 were considered statistically significant.
RESULTS
Antifungal susceptibility testing
MICs for fluconazole, micafungin, and amphotericin B were determined for all isolates in the collection by broth microdilution (Table 1). Fifty-two isolates (36.6%) were resistant to fluconazole (MIC ≥ 64 µg/mL), 13 (9.2%) were resistant to micafungin (MIC ≥ 0.25 µg/mL), and a single isolate (0.7%) was non-wild type to amphotericin B (MIC > 2 µg/mL). In total, 62 (43.7%) C. glabrata isolates were resistant to at least one antifungal drug (fluconazole, micafungin, or amphotericin B). Four isolates were multidrug resistant to two drug classes (fluconazole + micafungin). Four isolates had intermediate MICs to micafungin (MIC = 0.12 µg/mL).
Phylogenomic analysis
Phylogenomic analysis was carried out based on SNVs in the core genome (Fig. 1). Overall, the phylogeny represented 75.5% of the C. glabrata reference strain CBS 138 (ATCC 2001) genome. Most isolates (130/142, 91.5%) grouped into one of 15 genetically related clusters. A cluster was defined as two or more isolates that were less than 1,000 SNVs apart. Isolates within clusters differed by <10 to hundreds of SNVs (Table S3). Isolates between clusters differed by >13,500 SNVs, except for clusters XI and XII, which each differed by approximately 7,000 SNVs from the next most related isolate (Table S3). The five largest clusters (cluster I, n = 29; cluster III, n = 22; cluster VIII, n = 12; cluster X, n = 15; and cluster XV, n = 15) consisted of more than half (65.5%) of the isolates in our collection.
Maximum-likelihood phylogeny of C. glabrata isolates from Canada (n = 142) based on SNVs in the core genome. This phylogeny represents 75.5% of the core genome of reference strain CBS 138 (ATCC 2001). The 15 clusters (defined as two or more isolates <1,000 SNVs apart) are represented in various colors. Metadata includes the year of isolation, region of isolation, and resistance/non-wild type to fluconazole (red), micafungin (green), and/or amphotericin B (yellow). Antifungal-resistant/non-wild-type isolates are indicated by colored boxes.
There were several clusters that contained closely genetically related isolates (defined in this study as ≤20 SNVs). In cluster I (n = 29, 2-526 SNVs), there were three fluconazole-resistant isolates (MYC-19-0324, MYC-19-0341, and MYC-19-0344) and one micafungin-resistant isolate (MYC-19-0386) from the same province in the Central East region of Canada that differed by 2-16 SNVs (Fig. 1). Two of these isolates (MYC-19-0344 and MYC-19-0386) were collected from the same patient with differing susceptibility profiles and were separated by only two SNVs. The remaining isolates (MYC-19-0324 and MYC-19-0341) were recovered from separate patients at different healthcare facilities. A susceptible isolate (MYC-19-0291) from the Eastern region of Canada was also closely related to this group (8-14 SNVs). Within cluster VIII (n = 12, 2-681 SNVs), there were two susceptible isolates (MYC-19-0208 and MYC-19-0220) from the West and Central West regions of Canada separated by 3 SNVs (Fig. 1). There were several other instances of closely genetically related isolate pairs from the same patient (cluster III, MYC-19-0044 and MYC-19-0339, 3 SNVs; cluster VIII: MYC-19-0014 and MYC-19-0016, 2 SNVs; cluster X: MYC-19-0024 and MYC-19-0233, 13 SNVs; and cluster XIII, MYC-19-0007 and MYC-19-0008, 2 SNVs) observed in our study (Fig. 1).
Clusters did not group based on region of isolation within Canada (Fig. 1). Two potential minor exceptions were determined in clusters II and XI (Fig. 1). Isolates in cluster II (n = 3, 179-211 SNVs) were from the Central East region of Canada but from different provinces, while the two strains in cluster XI (n = 2, 297 SNVs) were from the same province in the Central East region of Canada; however, these associations were not significant (cluster III, P value = 0.0935; cluster XI, P value = 0.2078).
Some clusters appeared to be enriched for bloodstream or sterile site isolates. All strains in clusters IX (n = 6, 181-554 SNVs), XI (n = 2, 297 SNVs), and XII (n = 2, 200 SNVs) were isolated from blood (i.e., sterile site) (Table S1). Isolates in clusters II (n = 3, 179-211 SNVs) and XIII (n = 3, 2-37 SNVs) were collected from various sterile body sites (cluster II, bile = 1, blood = 2; cluster XIII, abdomen fluid = 1, blood = 2) (Table S1). These associations, however, were not statistically significant.
The majority of antifungal-resistant C. glabrata isolates (58 of 62, 93.5%) were present in 13 of 15 (86.7%) clusters (Fig. 1). Clusters XI (n = 2) and XII (n = 2) were the only clusters that did not contain at least one resistant isolate. Cluster I had the greatest number of antifungal-resistant isolates with 19/29 (65.5%) resistant to at least one antifungal drug (fluconazole, n = 14; micafungin, n = 3; and fluconazole + micafungin, n = 2) (Table 2). The associations of cluster I with antifungal resistance (P value = 0.0112) and fluconazole resistance (P value = 0.0298) were statistically significant, but not micafungin resistance (P value = 0.1403). Cluster III had the second largest number of antifungal-resistant isolates with 7/22 (31.8%) resistant to at least one antifungal drug (fluconazole, n = 4; micafungin, n = 1; amphotericin B, n = 1; and fluconazole + micafungin, n = 1); however, this cluster was not significantly associated with antifungal resistance (P value = 0.2509) or fluconazole resistance (P value = 0.1578) or amphotericin B resistance (P value = 0.1549) (Fig. 1). Fluconazole-resistant isolates (n = 52) were found in all clusters except for clusters XI and XII (Fig. 1). All micafungin-resistant isolates (n = 13) fell into one of five clusters (cluster I, n = 5; cluster III, n = 2; cluster VIII, n = 3; cluster XIV, n = 1, and cluster XV, n = 2) (Fig. 1). The sole amphotericin B-resistant isolate was found in cluster III (Fig. 1). There were four fluconazole-resistant C. glabrata isolates that did not cluster, highlighting that resistance was widely distributed within the phylogeny (Fig. 1).
Global phylogenomic analysis
We added 139 C. glabrata isolates from NCBI to our collection to analyze the Canadian isolates within a broader global context (Table S2) (25, 52–55). In total, there were 281 C. glabrata isolates from 10 countries (Australia, n = 62; Belgium, n = 6; Canada, n = 151; Denmark, n = 8; France, n = 9; Germany, n = 3; Italy, n = 2; Switzerland, n = 2; Taiwan, n = 2; and the United States, n = 36) included in the global phylogenomic analysis based on SNVs in the core genome. Since only isolates with reported MICs for fluconazole, micafungin, caspofungin, and/or amphotericin B were included, our global phylogeny is biased toward Canadian isolates, with more than half (53.7%) from Canada. Cluster labels from the Canadian phylogeny were applied to the global phylogeny using the same cluster definition of two or more isolates <1,000 SNVs. Of the 281 C. glabrata isolates, 271 (96.4%) grouped into one of 25 clusters that ranged in size from two to 63 isolates (Fig. 2). The isolates (PRJNA361477, Table S2) used to support the existence of seven major global clades of C. glabrata were included in our global analysis and were distributed across only eight of the clusters we observed using our <1,000 SNV cutoff (52). While it is challenging to compare lineages defined across studies due to the lack of standardized criteria, our global phylogenomic analysis highlights that the global diversity of C. glabrata exceeds previous reports.
Maximum-likelihood phylogeny of global C. glabrata isolates (n = 281) based on SNVs in the core genome. The global analysis includes isolates (highlighted in black) from 10 countries (Australia [AUS], n = 62; Belgium [BEL], n = 6; Canada [CAD], n = 151; Denmark [DNK], n = 8; France [FRA], n = 9; Germany [GER], n = 3; Italy [ITA], n = 2; Switzerland [SWI], n = 2; Taiwan [TWN], n = 2; and the United States [USA], n = 36) with susceptibility data for fluconazole, micafungin/caspofungin, and/or amphotericin B. Isolates resistant/non-wild type to fluconazole (red), micafungin/caspofungin (green), and/or amphotericin B (yellow) are represented in colored boxes.
Eight clusters of the global C. glabrata phylogeny grouped according to country (Australia: clusters XIX, XX, and XXI; Canada: clusters IV, VI, IX, and XIII; and Taiwan: cluster XXIV) (Fig. 2). Several of these clusters were significantly associated with geographic region of isolation (cluster IX, P value = 0.0322; cluster XXIV, P value = 0.0022; cluster XIX, P value = 0.0481; cluster XX, P value = 0.0481; and cluster XXI, P value = 0.0022).
Eighty-three isolates were resistant to azoles, 38 were resistant to echinocandins, and one was resistant to amphotericin B. Isolates resistant to at least one antifungal drug were found in 22 of 25 clusters (Fig. 2). No resistant isolates were present in clusters XI, XXII, and XXIV (Fig. 2). Similar to the Canadian phylogeny, clusters I and III were enriched with antifungal-resistant isolates (Fig. 2). In cluster I, 32/63 (50.8%) of isolates were resistant to at least one antifungal drug, while 13/34 (38.2%) were resistant in cluster III. The association of cluster I with antifungal resistance was not maintained at the global level (P value > 0.05).
Genetic mechanisms of antifungal resistance
We analyzed known target genes associated with antifungal resistance in C. glabrata to determine the genetic basis of resistance for isolates in our collection. For this analysis, we only considered variants that were unique to antifungal-resistant C. glabrata isolates. We investigated genes associated with azole resistance in C. glabrata, including PDR1, CDR1, PDH1 (CDR2), SNQ2, UPC2A, and ERG11 (Table S4). The HS regions of FKS1 and FKS2 were examined in micafungin-resistant C. glabrata. For the single amphotericin B non-wild-type isolate in our collection, we examined ERG2, ERG3, and ERG6. All variants identified in the analyzed genes are described in Table S4.
PDR1
Azole resistance in C. glabrata has been previously associated with three key regions in PDR1: the ID (312–382 aa), MHR (539–632 aa), and TAC (800–1107 aa) (16). Thirty-six PDR1 variants were identified in 38/52 (73.1%) fluconazole-resistant C. glabrata isolates (Table 3). Most PDR1 substitutions identified in fluconazole-resistant C. glabrata isolates were in gene regions that have previously been associated with azole drug resistance (ID, n = 6; MHR, n = 5; and TAC, n = 13) (Table 3). Twelve variants were outside of the regions previously described as being associated with azole resistance, spanning amino acid positions 124–291 (region preceding the ID) and 693–772 (between the MHR and the TAC regions) (Table 3).
Only three fluconazole-resistant isolates (7.9%) in our collection had PDR1 variants that were previously experimentally tested and shown to confer azole resistance in C. glabrata, namely G346D (45, 48), D876Y (16), and G1099D (45) (Table 3). The remaining 35 (92.1%) fluconazole-resistant isolates had either a suspected PDR1 resistance variant (i.e., not experimentally confirmed) reported in at least one previous study or a potential novel PDR1-resistant variant reported for the first time in this study (Table 3).
Thirteen PDR1 variants suspected to confer azole resistance in C. glabrata were found in 17 fluconazole-resistant isolates (Table 3). Suspected PDR1 variants are those that were identified in only azole-resistant C. glabrata isolates in at least one other previous study. Most isolates harbored a single, unique suspected PDR1 resistance variant, with the exceptions of G583S (n = 4) and D876N (n = 3), which were found in multiple isolates. Suspected PDR1 variants K274E and D876N occupy the same positions as experimentally confirmed variants K274N (56) and D876Y (16), respectively.
Twenty potentially novel PDR1 resistance variants were identified in 20 fluconazole-resistant C. glabrata isolates and were absent in susceptible-dose dependent isolates (Table 3). To our knowledge, this is the first instance these PDR1 variants have been reported. Most isolates with novel PDR1 variants had a single, unique variant except for N764I, which was found in two isolates. Some potential novel PDR1 resistance variants, including W256R (57), L291F (16, 51), G611V (46), R761G (46, 54), N764I (16, 43, 49), and E1083G (16, 50), were located in the same positions as previously suspected PDR1 resistance variants but with an alternate amino acid change.
Several PDR1 variants were identified in our collection that did not contribute to azole resistance in C. glabrata. PDR1 variants V27A, S391L, T488R, and S651N were present exclusively in susceptible-dose dependent isolates, while S76P, V91I, L98S, V134A, L139I, T143P, D243N, R250K, G493D, and T745A were identified in both susceptible-dose dependent and resistant isolates (Table S4).
Other genes associated with azole resistance
Apart from PDR1, there are several other genes that have been implicated in azole resistance in C. glabrata (Table S4). We investigated variants in efflux pumps belonging to the ABC transporter superfamily (e.g., CDR1, PDH1, and SNQ2) as potential mechanisms of azole resistance. There were four fluconazole-resistant C. glabrata isolates that had CDR1 variants (H1098Y, n = 3; and A1120T, n = 1). PDH1 (CDR2) variants were found in three fluconazole-resistant C. glabrata isolates (G872S, n = 1; H1129R, n = 1; and K1130I, n = 1). Five fluconazole-resistant isolates had variants in SNQ2 (T613S, n = 2; G624R, n = 1; E823D, n = 1; and K830N, n = 1). Substitutions in ERG11 and UPC2A are known mechanisms of azole resistance in other Candida species, such as C. albicans; however, little evidence has been reported in C. glabrata. Seven fluconazole-resistant C. glabrata isolates (L20P, n = 1; L97F, n = 1; K152R, n = 1; and V155A, n = 4) had ERG11 variants, while two fluconazole-resistant isolates had UPC2A variants (T260QinsQQ, n = 1; and L528I, n = 1). Isolates that carried either CDR1 A1120T, SNQ2 E823D/K830N, or ERG11 L20P did not contain PDR1 substitutions that could potentially represent genetic determinants of azole resistance (Table S4). The CDR1 variant was located downstream of the second ABC transporter (857-1099 aa), while the SNQ2 variants were located upstream. There were no similar variants identified in C. glabrata or other Candida species that were associated with azole resistance in either CDR1 or SNQ2 (58). The ERG11 L20P variant was located near the N-terminal, distant from the catalytic region that is associated with resistance. In C. albicans, an adjacent substitution in ERG11 (Q21L) was identified but had a limited effect on azole susceptibility when experimentally tested (59). Further testing is required to experimentally confirm the role of these potential novel variants in azole resistance in C. glabrata.
FKS genes
Substitutions in FKS1 (amino acid positions 625 and 629 in HS1) and FKS2 (positions 659, 660, 663, and 664 in HS1; positions 1375 and 1377 in HS2) are most strongly associated with echinocandin resistance in C. glabrata (21). In our collection, there were 13 micafungin-resistant C. glabrata isolates, and 12 (92.3%) had FKS HS variants that were previously correlated with echinocandin resistance, while one isolate was wild type (Table 4). Most variants (10 out of 12) were found at positions 659 and 663 in FKS2 HS1 (F659del, n = 3; S663P, n = 6; and S663F, n = 1). The remaining two variants were in FKS1 HS1 (S629P, n = 1; and D632V, n = 1). Of the four micafungin non-susceptible C. glabrata isolates (MYC-19-0008, MYC-19-0272, and MYC-19-0315) that had intermediate MICs, only one isolate (MYC-19-0315) had an FKS variant that was previously considered to be weakly associated with echinocandin resistance (FKS2 HS2, R1378S).
ERG genes
The mechanisms of polyene resistance in C. glabrata are not well characterized; however, specific ERG genes, including ERG2, ERG3, and ERG6, involved in the ergosterol biosynthetic pathway, have been implicated (Table S4). There were no variants of interest identified in our single amphotericin B non-wild-type isolate.
Copy number variant analysis
We identified CNVs in genes unique to fluconazole-resistant C. glabrata isolates. In total, there were 146 genes with copy numbers that deviated from the expected ploidy (Table S5). One isolate (MYC-19-0345) had two copies of PDR1 in combination with suspected (G1079R) and potential novel (G189V) PDR1 substitutions. There were amplifications of three other genes (CAGL0D06512g, CAGL0K05841g, and CAGL0F08041g) that may be associated with azole resistance in C. glabrata (Table S5). Further testing is required to determine the significance of increased gene copies on azole resistance in C. glabrata.
Associations between clusters and resistance variants in the Canadian collection
There were several examples of cluster-specific variants. The four isolates (MYC-19-0087, MYC-19-0181, MYC-19-0317, and MYC-19-0334; 17-70 SNVs) that had the suspected PDR1 variant G583S all belonged to cluster I. There were also two isolates in cluster III (MYC-19-0015 and MYC-19-0083, 59 SNVs) that harbored the potential novel PDR1 N764I variant. The three isolates (MYC-19-0015, MYC-19-0083, and MYC-19-0333; 59-300 SNVs) that harbored the CDR1 variant H1098Y were all located in cluster III. MYC-19-0015 and MYC-19-0083 (cluster III, 59 SNVs) also contained the T613S variant in SNQ2. All four isolates (MYC-19-0087, MYC-19-0181, MYC-19-0317, and MYC-19-0334; 17-70 SNVs) that had the ERG11 V155A variant were present in cluster I.
There were also examples of variants that were present in multiple clusters. The three isolates (MYC-19-0273, MYC-19-0297, and MYC-19-0320; 23338-24371 SNVs) that had the suspected PDR1 variant D876N were present in three separate clusters. PDR1 variants occupying the same position but with an alternate amino acid were found within the same and different clusters. The confirmed PDR1 variant D876Y found in a single isolate (MYC-19-0319) was also in the same cluster as an isolate that carried the D876N PDR1 variant. There were two isolates (MYC-19-0025 and MYC-19-0346) that had a PDR1 variant in position R761 with two different amino acid changes that were found in separate clusters. Most FKS variants identified in more than one micafungin-resistant isolate did not group together within clusters. The six isolates (MYC-19-0093, MYC-19-0094, MYC-19-0145, MYC-19-0159, MYC-19-0180, and MYC-19-0386) that carried the FKS2 HS1 variant S663P were found in four separate clusters (clusters I, VIII, XIV, and XV; 231-26990 SNVs). FKS variants that had different amino acid alterations at the same position were also not associated with the same cluster. For example, the FKS2 HS1 variant S633F was found in a different cluster from the six isolates that contained the S663P variant.
DISCUSSION
The global rise of invasive C. glabrata is worrisome since infections are becoming increasingly associated with antifungal resistance (7). While the rates of azole and echinocandin resistance in Canada were low during 2012–2016, higher rates have been reported elsewhere, highlighting the importance of understanding the mechanisms and dissemination of resistance (8, 60). In this study, we sequenced a comprehensive collection of 142 clinical C. glabrata isolates from across Canada, representing the largest national genomic study of C. glabrata from a single country to date. This collection was uniquely enriched with antifungal-resistant isolates from invasive infections to better understand the genomic phylogeny of resistant infection types. A genomics approach was also used to identify variants in genes associated with antifungal resistance to determine the genetic basis of resistance observed by phenotypic susceptibility testing.
Whole-genome sequencing is a powerful tool for resolving the genomic phylogeny of C. glabrata, offering high resolution and superior discriminatory power by analyzing genetic variation across the entire genome (26). The maximum-likelihood phylogeny of C. glabrata isolates from Canada shown in this study represented more than 75% of the core genome, capturing a far stronger and more finely resolved phylogenetic signal compared to other traditional typing methods, such as MLST, which typically rely on six genetic loci (FKS1, LEU2, NMT1, TRP1, UGP1, and URA3) to differentiate strains of C. glabrata (61).
A previous study based on 32 globally distributed isolates proposed the existence of seven major clades of C. glabrata (52). Our global analysis, which included these isolates, provided further evidence that the diversity of C. glabrata is much greater than originally observed, consistent with other studies (57, 62). In the absence of previously published criteria to define clusters, we used a cutoff of 1,000 SNVs to define a cluster. Clusters were separated by at least 20,000 SNVs in most cases. Using these criteria, we determined there were at least 15 genetically related lineages of C. glabrata in Canada. As more sequences of C. glabrata become publicly available for phylogenomic analysis, developing a standardized set of criteria for denoting clusters based on SNV thresholds will allow for more meaningful comparisons across studies.
C. glabrata has sometimes displayed geographic specificity of clusters (61). There were no significant associations between clusters and regions within Canada; however, some clusters were significantly associated with isolates from Canada, Australia, and Taiwan in our global analysis. In terms of infection source, previous studies have not found differences in clusters based on invasive and non-invasive isolates (63). Our study supports these findings as clusters were not associated with sterile or non-sterile sites of infection. We did, however, identify a large cluster that was significantly associated with antifungal resistance in our Canadian collection. Cluster I (n = 29) contained sterile and non-sterile isolates from all regions of Canada, with approximately two-thirds of the isolates resistant to at least one antifungal drug. This is the first reported instance of a C. glabrata lineage being associated with antifungal resistance to our knowledge, which may provide evidence of a strain of C*. glabrata* with a higher propensity to develop resistance.
There were several pairs of isolates in our collection that were serial isolates from the same patient. These within-patient isolates were closely related, differing by <20 SNVs, suggesting the same strain was isolated from the patient each time. One pair of isolates in our collection had different susceptibilities, the first isolate being fluconazole-resistant, while the second was fluconazole-susceptible, dose-dependent, but micafungin-resistant. This may reflect the evolution of resistance that can occur within a patient due to fitness costs and selective pressure during antifungal therapy. Other studies have included isolates from the same patient (57, 62). In one study, four pairs of isolates from separate patients were sequenced to determine if the patient harbored more than one strain of C. glabrata. Of the four pairs of isolates, only one patient had two different strains of C. glabrata (25). In one study, blood cultures from patients were observed to contain genetically diverse strains (64).
Whole-genome sequencing has also been used to detect molecular mechanisms of resistance in C. glabrata. We examined genes associated with azole and echinocandin resistance to determine the genetic basis of antifungal resistance. There were 58 fluconazole-resistant C. glabrata isolates, and 38 had variants in PDR1 that were primarily found in regions that were previously correlated with azole resistance. Few studies have experimentally confirmed the effect of PDR1 variants on azole resistance (14–16, 45, 56, 65). Only three isolates in our collection harbored experimentally verified variants of PDR1. Over 150 PDR1 variants identified in previous studies of C. glabrata are suspected to decrease azole susceptibility; however, these variants have not been experimentally validated (16, 42–44, 46, 47, 49–51, 57, 66–68). Most isolates in our collection had PDR1 variants that could potentially describe azole resistance. Some PDR1 variants have been previously reported in azole-resistant C. glabrata isolates in at least one other study, and detection of these suspected PDR1 variants in our collection lends further support that these variants may indeed be associated with azole resistance. Some suspected PDR1 variants fall outside of PDR1 regions associated with azole resistance. Twenty PDR1 variants were reported for the first time here. Some variants occupied the same position as previously identified PDR1 variants, but with alternate amino acid substitutions. Experimental testing of a greater number of PDR1 variants identified within and outside of the key regions will better delineate variants associated with azole resistance.
Almost all micafungin-resistant isolates in our collection had FKS1 or FKS2 variants (at position 629/632 or 659/663, respectively) that are strongly associated with echinocandin resistance. A single isolate harboring no FKS mutations yet was phenotypically resistant, suggesting that it may have a currently unknown mechanism of resistance. Our study also supports the previous finding that echinocandin resistance in C. glabrata is more strongly associated with FKS2 HS variants compared to FKS1 in 10 out of 13 variants we have described (46, 69).
Our study had several limitations. The scope was limited to the analysis of genes known to be associated with resistance to the three major classes of antifungals and provides no insight into novel resistance genes. Future genome-wide association studies may identify significant variants in novel resistance genes or the combined effect of mutations in multiple target genes. Since we did not have access to patient health information, we were unable to analyze correlations between patient treatment history and antifungal resistance. Similarly, without access to travel history, we were also unable to determine if the Canadian isolates clustered within the global analysis were potentially acquired during travel or medical tourism. Finally, experimentally validating the role of suspected and potential novel resistance variants through gene editing and/or gene expression analysis is essential to confirm their role in antifungal resistance.
In summary, whole-genome sequencing of the largest collection of clinical C. glabrata isolates from Canada to date was useful for determining the genomic phylogeny and potential genetic mechanisms of antifungal resistance of this pathogen across Canada. We observed high levels of genetic diversity, with at least one cluster associated with phenotypic antifungal resistance. Most fluconazole-resistant isolates harbored suspected or potential novel resistance variants in PDR1, while micafungin resistance was primarily described by substitutions in the HS regions of FKS genes. Antifungal-resistant isolates were present in almost all clusters, suggesting that resistance most likely arises independently from selective pressure during antifungal therapy rather than dissemination of resistant clones. Our study provides a baseline for future genomic studies of C. glabrata and outbreak investigations in Canada. Future work should focus on experimentally testing resistance variants to improve genotypic prediction of antifungal resistance in C. glabrata.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nucci M, Anaissie E. 2001. Revisiting the source of candidemia: skin or gut? Clin Infect Dis 33:1959–1967. doi:10.1086/32375911702290 · doi ↗ · pubmed ↗
- 2Rodrigues CF, Silva S, Henriques M. 2014. Candida glabrata: a review of its features and resistance. Eur J Clin Microbiol Infect Dis 33:673–688. doi:10.1007/s 10096-013-2009-324249283 · doi ↗ · pubmed ↗
- 3Vazquez JA, Dembry LM, Sanchez V, Vazquez MA, Sobel JD, Dmuchowski C, Zervos MJ. 1998. Nosocomial Candida glabrata colonization: an epidemiologic study. J Clin Microbiol 36:421–426. doi:10.1128/JCM.36.2.421-426.19989466752 PMC 104553 · doi ↗ · pubmed ↗
- 4Goemaere B, Lagrou K, Spriet I, Hendrickx M, Becker P. 2018. Clonal spread of Candida glabrata bloodstream isolates and fluconazole resistance affected by prolonged exposure: a 12-year single-center study in Belgium. Antimicrob Agents Chemother 62:e 00591-18. doi:10.1128/AAC.00591-1829784839 PMC 6105788 · doi ↗ · pubmed ↗
- 5Abbes S, Sellami H, Sellami A, Makni F, Mahfoudh N, Makni H, Khaled S, Ayadi A. 2011. Microsatellite analysis and susceptibility to FCZ of Candida glabrata invasive isolates in Sfax Hospital, Tunisia. Med Mycol 49:10–15. doi:10.3109/13693786.2010.49356120586679 · doi ↗ · pubmed ↗
- 6Arastehfar A, Lass-Flörl C, Garcia-Rubio R, Daneshnia F, Ilkit M, Boekhout T, Gabaldon T, Perlin DS. 2020. The quiet and underappreciated rise of drug-resistant invasive fungal pathogens. J Fungi 6:138. doi:10.3390/jof 6030138 PMC 755795832824785 · doi ↗ · pubmed ↗
- 7Pfaller MA, Diekema DJ, Turnidge JD, Castanheira M, Jones RN. 2019. Twenty years of the SENTRY antifungal surveillance program: results for Candida species From 1997-2016. Open Forum Infect Dis 6:S 79–S 94. doi:10.1093/ofid/ofy 35830895218 PMC 6419901 · doi ↗ · pubmed ↗
- 8Fuller J, Dingle TC, Bull A, Shokoples S, Laverdière M, Baxter MR, Adam HJ, Karlowsky JA, Zhanel GG, Canadian Antimicrobial Resistance Alliance (CARA) and CANWARD. 2019. Species distribution and antifungal susceptibility of invasive Candida isolates from Canadian hospitals: results of the CANWARD 2011-16 study. J Antimicrob Chemother 74:iv 48–iv 54. doi:10.1093/jac/dkz 28731505645 · doi ↗ · pubmed ↗