Global epidemiological and genetic characteristics of carbapenem-resistant Escherichia coli carrying blaIMP
Yueyue Hu, Chengjin Wu, Jiayao Yao, Jingyi Guo, Jie Sheng, Xinru Wang, Longjie Zhou, Xinyan Tang, Haotian Xu, Yunsong Yu, Xi Li, Xinhong Han

TL;DR
This study examines the global spread and genetic traits of Escherichia coli resistant to carbapenems due to the blaIMP gene, highlighting the role of plasmids in their transmission.
Contribution
The study provides the first comprehensive genomic and epidemiological analysis of blaIMP-positive CRECs, emphasizing their plasmid-mediated spread and global diversity.
Findings
blaIMP-positive CRECs carry highly transferable and stable IncN plasmids, enabling their regional and global dissemination.
Global analysis identified 82 sequence types, with ST131 being the most prevalent and showing clonal spread.
A strong correlation exists between antibiotic resistance genes and plasmid replicons, underscoring plasmids' role in resistance transmission.
Abstract
Carbapenem-resistant Escherichia coli (CREC) is a serious global health threat. While blaNDM-positive and blaKPC-positive CRECs are well-characterized, the genomic and epidemiological features of blaIMP-positive CRECs remain poorly understood. This study provides a comprehensive analysis of blaIMP-positive CRECs by integrating local and global surveillance data. We characterized nine blaIMP-4-positive CRECs from 94 CRECs collected from four tertiary hospitals in Zhejiang, China (2017–2024), using whole-genome sequencing and assays for antimicrobial susceptibility, plasmid transfer, stability, and fitness. The nine isolates were multidrug-resistant but susceptible to colistin, tigecycline, and amikacin. Seven diverse sequence types (STs) were identified, with high-risk ST1193 clones being first detected. The isolates had similar blaIMP-carrying IncN plasmids with high conjugation…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
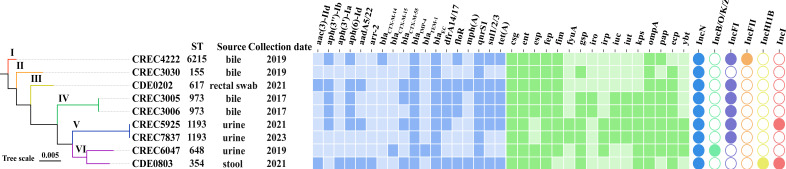 Fig 1
Fig 1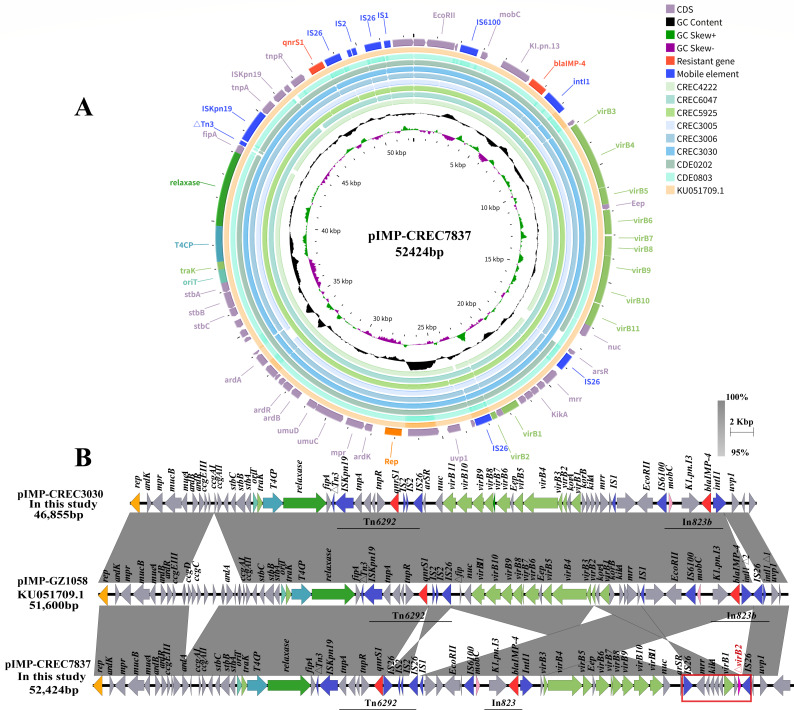 Fig 2
Fig 2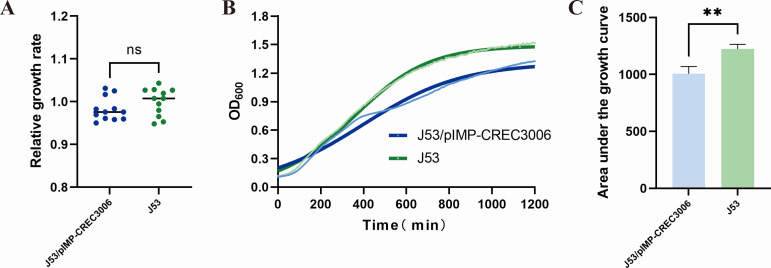 Fig 3
Fig 3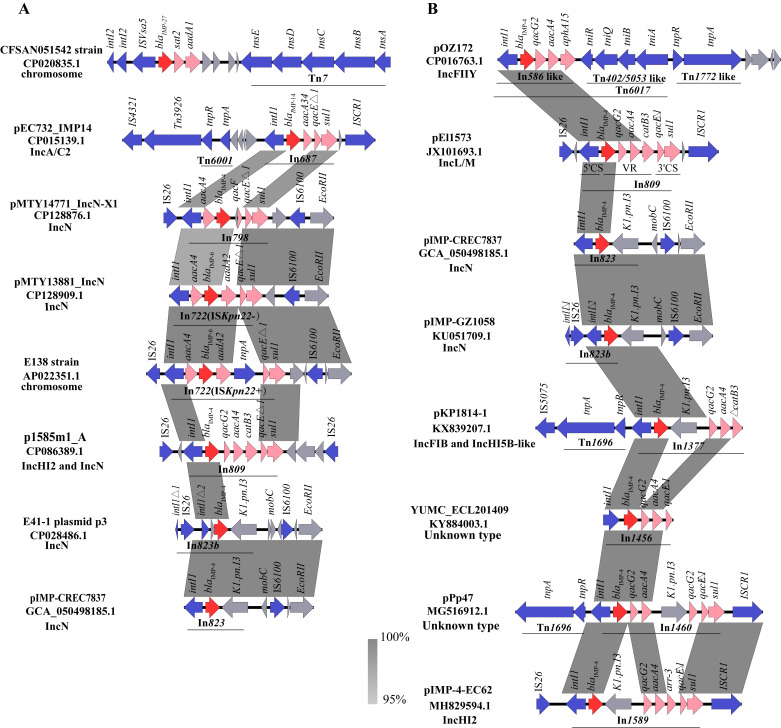 Fig 4
Fig 4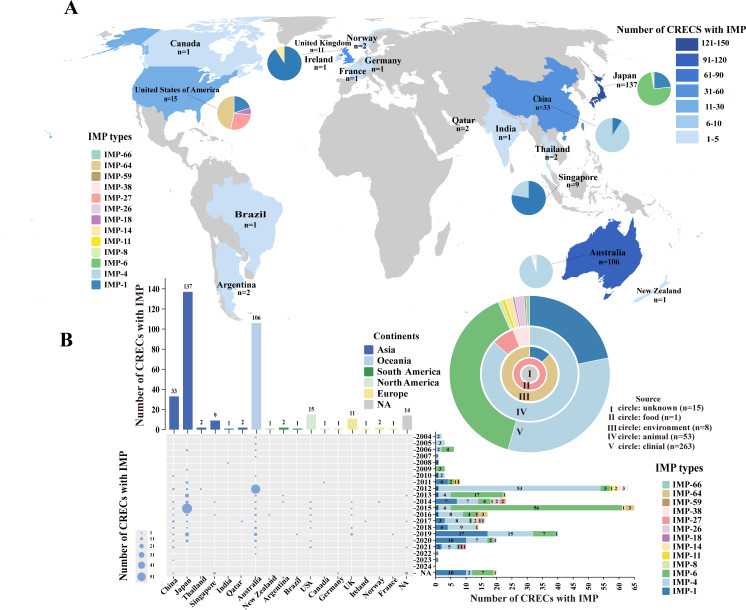 Fig 5
Fig 5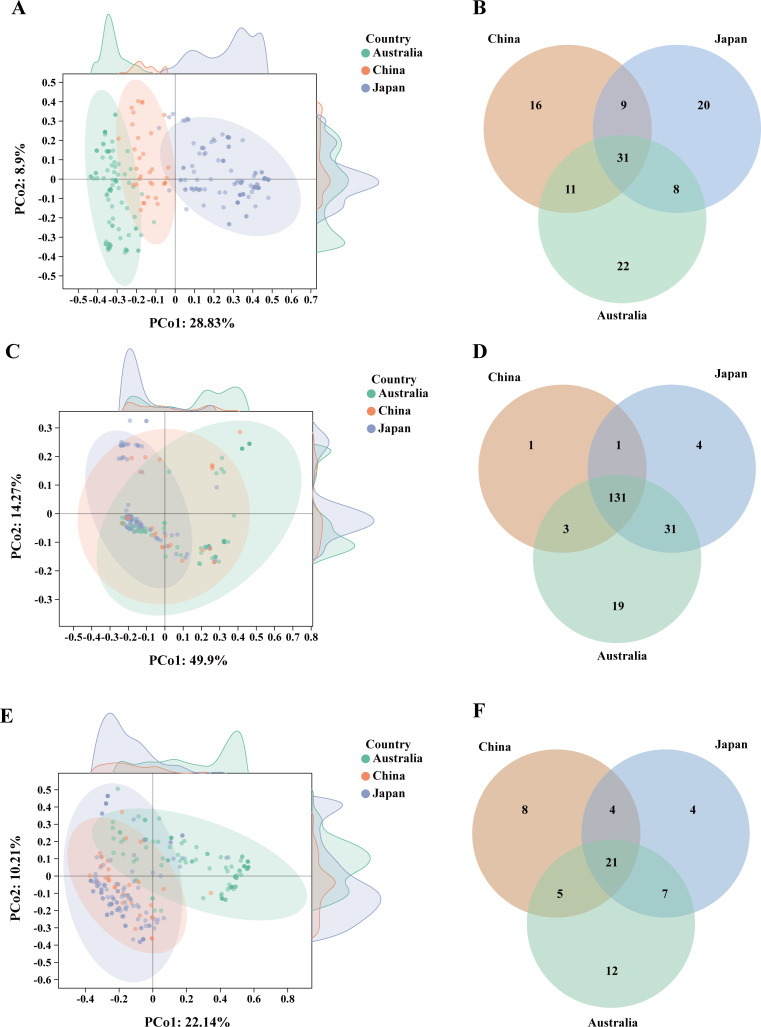 Fig 6
Fig 6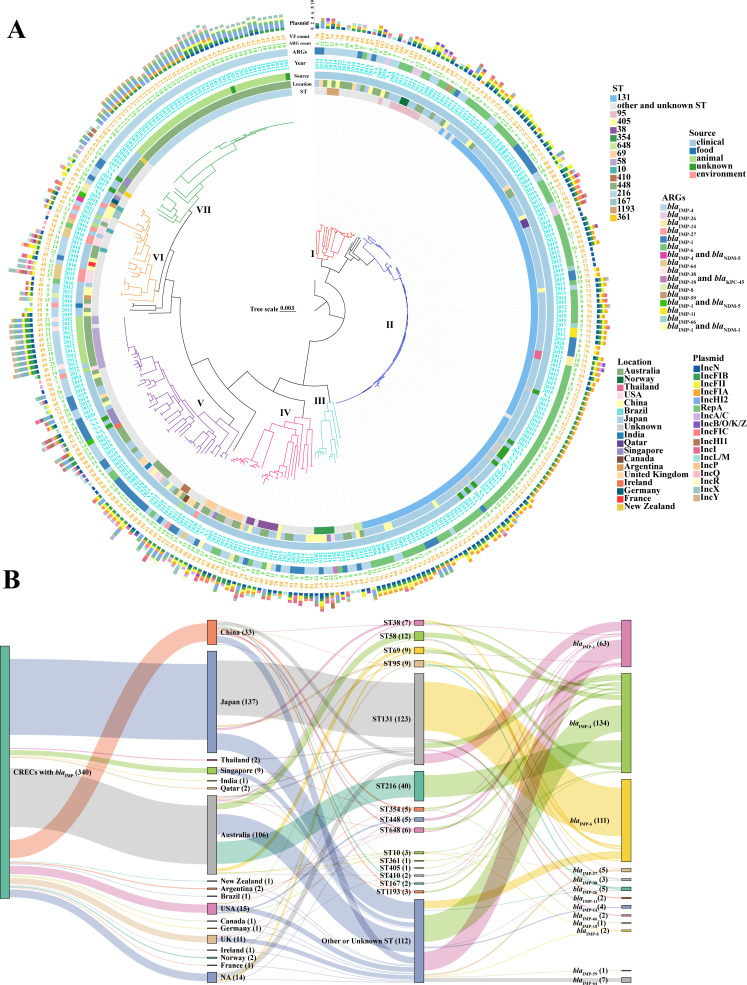 Fig 7
Fig 7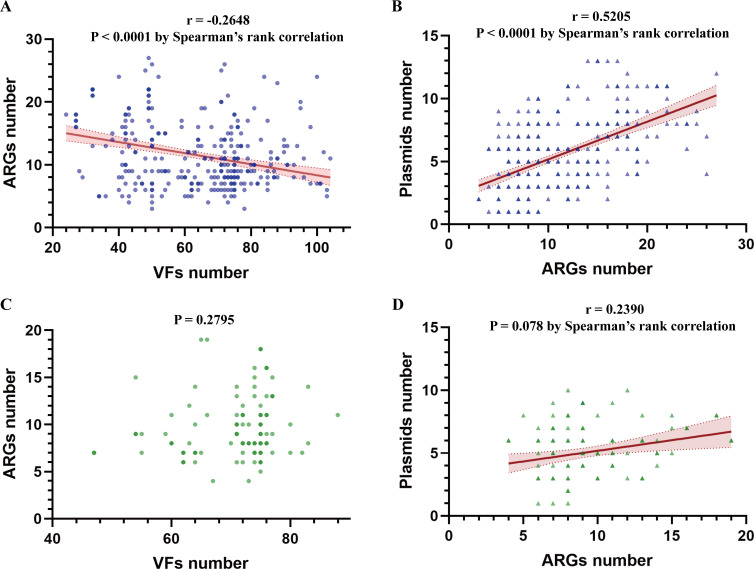 Fig 8
Fig 8| Isolates | MICs (μg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ETP | FEP | MEM | IPM | CAZ | CZA | AK | CST | TGC | CIP | |
| CREC4222 | 64 | >128 | 16 | 8 | >128 | >128 | 2 | <0.125 | 0.0625 | 32 |
| CREC6047 | 64 | >128 | 16 | 4 | >128 | 128 | 1 | <0.125 | 0.5 | 32 |
| CREC5925 | 8 | >128 | 2 | 2 | >128 | >128 | 1 | <0.125 | 0.0625 | 32 |
| CREC3030 | 32 | 32 | 16 | 2 | >128 | >128 | 4 | <0.125 | 0.5 | 0.25 |
| CREC3005 | 32 | 64 | 16 | 4 | >128 | >128 | 2 | <0.125 | 0.25 | 32 |
| CREC3006 | 32 | 16 | 8 | 4 | >128 | >128 | 1 | <0.125 | 0.5 | 32 |
| CDE0202 | 128 | >128 | 32 | 8 | >128 | >128 | 1 | <0.125 | 0.5 | 32 |
| CREC7837 | 32 | 128 | 8 | 4 | >128 | >128 | 2 | <0.125 | 0.0625 | 32 |
| J53/pIMP-CREC4222 | 16 | 64 | 16 | 4 | >128 | >128 | 1 | <0.125 | 0.125 | 0.25 |
| J53/pIMP-CREC6047 | 16 | 128 | 16 | 4 | >128 | >128 | 1 | <0.125 | 0.125 | 0.5 |
| J53/pIMP-CREC5925 | 8 | >128 | 2 | 2 | >128 | 64 | 4 | <0.125 | 0.125 | 0.25 |
| J53/pIMP-CREC3030 | 8 | 4 | 2 | 1 | >128 | >128 | 2 | <0.125 | 0.5 | 0.25 |
| J53/pIMP-CREC3005 | 32 | 128 | 16 | 4 | >128 | >128 | 2 | <0.125 | 0.5 | 0.25 |
| J53/pIMP-CREC3006 | 32 | 16 | 8 | 2 | >128 | >128 | 0.5 | <0.125 | 0.5 | 0.5 |
| J53/pIMP-CDEC0202 | 16 | 16 | 8 | 2 | >128 | >128 | 0.5 | <0.125 | 0.25 | 0.25 |
| J53/pIMP-CDEC0803 | 8 | 4 | 8 | 2 | >128 | >128 | 1 | <0.125 | 0.25 | <0.125 |
| J53 | <0.125 | <0.125 | 0.25 | 0.25 | 0.5 | 0.5 | 1 | <0.125 | 0.5 | <0.125 |
| ATCC25922 | <0.125 | <0.125 | <0.125 | <0.125 | 0.25 | 0.25 | 1 | <0.125 | <0.125 | <0.125 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotic Use and Resistance · Escherichia coli research studies
INTRODUCTION
Carbapenems, with their good permeability and strong antimicrobial activity, were often used to treat serious infections with multidrug-resistant Gram-negative organisms (1). However, in recent years, the worldwide overuse of β-lactam drugs (especially carbapenems) has led to increasing reports of carbapenem-resistant Enterobacteriaceae and even the rapid spread of nosocomial infections (2). The China Antimicrobial Surveillance Network 2024 (https://www.chinets.com/) report showed that Escherichia coli was the primary clinically isolated strain. Carbapenem-resistant Escherichia coli (CREC) slightly increased to 3.6%, up by 0.5% from last year. So the total number of CRECs should not be overlooked. CRECs can not only persist in hospital environments and spread among patients and healthcare workers, but also disseminate widely in sewage, surface water, vegetation soil, as well as in animals, food, and healthy populations, posing a significant threat to public health (3–6).
The main mechanism of carbapenem resistance in E. coli is the production of carbapenemases, among which IMP-type metallo-β-lactamases are one of the prevalent acquired carbapenemases. The IMP-1 enzyme was detected in Pseudomonas aeruginosa by Japanese scholars in 1991 (7). As time progressed, more and more IMP variants appeared, including IMP-2 (8), IMP-4 (9), IMP-6 (10), and so on. Currently, there are 111 IMP variants (http://bldb.eu/, last updated on 18 September 2025). IMP variants exhibit different hydrolysis efficiencies, and their diversity enables bacteria to continuously adapt to complex environments, posing severe challenges to antibiotic stewardship (11–13).
Some blaIMP-positive CRECs have low levels of imipenem resistance, which leads to an underestimation of IMP prevalence; for instance, IMP-6 remains susceptible to imipenem (14). The blaIMP gene in E. coli usually co-exists with blaCTX-M resistance genes or other resistance mechanisms, leading to resistance to multiple antibiotics such as cephalosporins, aminoglycosides, quinolones, carbapenems, and novel β-lactamase inhibitor combinations (e.g., ceftazidime-avibactam). In addition, most blaIMP genes are located on plasmids and frequently associated with integrons, insertion sequences, and transposons, which mediate their efficient horizontal transfer within the Enterobacteriaceae (11). Previous studies showed that blaIMP genes not only had a higher prevalence in Asian countries and Australia, but also had the ability to spread worldwide (15, 16). The blaIMP-positive CRECs, with high transmissibility and drug resistance, pose severe challenges to clinical treatment and public health.
However, a detailed characterization of blaIMP-positive CRECs and a systematic global epidemiological analysis remain limited. This study aims to address this gap by comprehensively characterizing the nine blaIMP-4-positive CREC strains isolated from Zhejiang, China, with a particular focus on their plasmid transmission properties, and by elucidating the global epidemiological status, transmission features, and potential threats of blaIMP-positive CRECs through a large-scale analysis of publicly available genomic data. This study will enable us to understand their distribution and transmission and help us formulate effective regulatory measures.
RESULTS
Characteristics of the nine CRECs
Among the 94 CRECs, 77 isolates were NDM carbapenemase producers, nine isolates carried the blaIMP-4 gene, and four isolates carried the blaKPC-2 gene. In addition, among the 94 CRECs, 4 isolates also exhibited carbapenem resistance mediated by other uncharacterized mechanisms. In this study, the nine blaIMP-4-positive CRECs were further studied. The patients were all over 60 years old, and the strains were mainly isolated from bile, urine, and rectal swabs (Table S1). After whole-genome sequencing (WGS), the nine strains all carried IncN plasmids, blaIMP-4 and qnrS1 resistance genes. The majority of these isolates also contained extended-spectrum β-lactamase gene (blaCTX-M), aminoglycoside genes (aph(6)-Id, aph(3'')-Ib), sulfonamide gene (sul), trimethoprim gene (dfrA), and tetracycline gene (tet(A)). In addition, all isolates carried multiple virulence genes, including those for bacterial biofilm formation, adhesion, and iron uptake; genes encoding enterotoxin and an effector protein (which interferes with signaling pathways); and genes associated with bacterial immune evasion mechanisms (Fig. 1; Table S2). A phylogenetic tree was constructed based on genomic single nucleotide polymorphisms (SNPs), and the nine strains could be divided into six distinct phylogenetic clades (Fig. 1). According to previous studies, clonal dissemination can be defined based on a pairwise SNP distance of ≤200 between E. coli isolates (17, 18). Combined with multilocus sequence typing (MLST), it was found that CREC5925 and CREC7387 were the same clone (ST1193), and CREC3005 and CREC3006 shared the same clone (ST973). For the remaining strains, their sequence types (STs) were relatively scattered, with a large difference in SNPs (Fig. S1).
Characteristics of the nine CRECs. Distribution of representative ARGs, VFs, plasmid Inc. types (not all are shown here), and phylogenetic relationships of the nine Escherichia coli from this study. The solid figures represent the presence of the corresponding ARGs, VFs, and plasmid replicons.
Antimicrobial susceptibility testing
According to minimal inhibitory concentrations (MICs), the results of antimicrobial susceptibility testing showed that all collected strains were resistant to ceftazidime, cefepime, ceftazidime-avibactam, and ertapenem. Except for CREC3030, the remaining strains (88.89%) were resistant to ciprofloxacin. For meropenem, only CREC5925 showed intermediate susceptibility, while the other strains had low-level resistance (MICs: 4 mg/L–16 mg/L). Additionally, three strains (CREC5925, CREC3030, and CDE0803) exhibited intermediate susceptibility to imipenem, and the rest had low-level resistance. All strains were sensitive to polymyxin, tigecycline, and amikacin, providing potential options for treatment (Table 1). Except for CREC7837, which failed to conjugate successfully, the blaIMP-4 genes of the other eight strains (88.89%, 8/9) were verified to have successfully conjugated to the recipient strain E. coli J53 (Table 1).
Genomic characteristics of blaIMP-4-carrying IncN plasmids
In plasmid map structure comparison, nine strains contained blaIMP-4-carrying IncN plasmids with high consistency and similar backbone structures (Fig. 2A). We further performed third-generation sequencing on the CREC7837 strain. In NCBI BLAST, the available pIMP-CREC7837 and pIMP-CREC3030 plasmid sequences exhibited 99.99% nucleotide identity and 100% coverage with pIMP-GZ1058 (KU051709.1) from E. coli (Fig. 2B). These three IncN plasmids featured rep, mobC, stbABC, ardABRK, mucAB, and kikA genes essential for replication and maintenance. In addition, a conjugation transfer system including origin of transfer site (oriT), relaxase, the type IV coupling proteins (T4CPs), and the type IV secretion system (T4SS) (virB1–11) was found. It is noteworthy that the virB2 gene in the pIMP-CREC7837 plasmid was partially deleted. The IncN plasmid carried two resistant regions, including the blaIMP-4 located on a class 1 integron and the qnrS1 located on the Tn6292 transposon. In pIMP-CREC7837 and pIMP-CREC3030 plasmids, the intI1 integrase of class 1 integron (In823) captured the blaIMP-4 resistance gene cassette, tandem with group II intron K1.pn.I3, a mobilization protein gene (mobC), and IS6100 insertion sequences to form the structure of intI1-blaIMP-4-K1.pn.I3-mobC-IS6100. The qnrS1 was detected on Tn6292, which was a complex transposon with a core structure consisting of the following components: ISKpn19-tnpA-tnpR-qnrS1-IS2-IS26 (Fig. 2B).
Genomic characteristics of the blaIMP-4 gene in our strains. (A) Circular sequence alignment of plasmids bearing blaIMP-4. The pIMP-CREC7837 plasmid served as a reference for alignment with the other eight blaIMP-4 CRECs in this study and pIMP-GZ1058 (KU051709.1), similar plasmids available from the NCBI database. (B) Linear sequence alignment. Genetic environment of blaIMP-4 on pIMP-CREC7837, pIMP-GZ1058, and pIMP-CREC3030 plasmids. The regions with highly similar sequences were marked in gray (nucleotide identity of ≥95%). The blaIMP-4 and qnrS1 genes are shown in red; mobile elements were drawn in blue.
These three plasmids also showed differences: (i) In the pIMP-CREC7837 plasmid, the insertion of IS26 on both sides of virB1 and ΔvirB2 led to the rearrangement of T4SS genes. And the IS26 inserted downstream of the 209 bp sequence site of the virB2 gene resulted in the deletion of the subsequent sequence, which may lead to the destruction of virB2 gene integrity, loss of conjugative pilus function, and finally the failure of conjugation. (ii) In pIMP-CREC7837 and pIMP-CREC3030, the intI1 integrase was intact, whereas in pIMP-GZ1058, intI1 was truncated by IS26 to form the In823b structure. (iii) In addition, due to IS26 inserted between qnrS1 and IS2, gene rearrangement occurred downstream of qnrS1 in the pIMP-CREC7837 plasmid (Fig. 2B).
Plasmid transfer capacity, plasmid stability, and fitness cost
Plasmid conjugation frequency assay showed the blaIMP-4-carrying IncN plasmids were transferred by conjugation to the E. coli J53 recipient at high frequencies of 5.3 × 10^−2^ to 8.7 × 10^−1^ per donor cell. Due to the nine strains harbored similar blaIMP-4-carrying IncN plasmids, we randomly selected one J53/pIMP-CREC3006 transconjugant to assess the fitness cost and plasmid stability. The results showed that there was no statistically significant difference in the relative growth rate between the J53/pIMP-CREC3006 transconjugant and E. coli J53 (P > 0.05). But a statistically significant difference was observed in the area under the growth curve (AUC) at 20 h (decreased by approximately 17.73%, P < 0.05). These results collectively indicated that the transconjugant carrying the IncN plasmid had a slight fitness cost (Fig. 3). In addition, the J53/pIMP-CREC3006 transconjugant was cultured for 10 consecutive days without antibiotics, and the IncN plasmid remained 100% after 100 generations. These results indicated that the plasmid exhibited high stability, strong transfer ability, and slight fitness cost, which facilitated the dissemination of the blaIMP-4 genes.
*Growth profile of the recipient strain E. coli J53 and transconjugant E. coli J53/pIMP-CREC3006 (A–C). (A) Relative growth rate. (B) Growth curve. (C) AUC. This experiment was performed in four replicates and repeated three times. (ns > 0.05; *P < 0.05; *P < 0.01; unpaired t tests with F test).
Core genetic characterization of the blaIMP in E. coli
The blaIMP gene was usually located on the integron within a plasmid, leading to the horizontal transfer of resistance genes. Therefore, we further explored the blaIMP gene context in E. coli. All available complete genome sequences of blaIMP-positive E. coli at NCBI were downloaded. The schematic plot contained five blaIMP genotypes: blaIMP-1, blaIMP-4, blaIMP-6, blaIMP-14, and blaIMP-27 (Fig. 4A). It was worth mentioning that the blaIMP-27 gene was located in the chromosomal class 2 integron with intI2-ISVsa5-blaIMP-27-sat2-aadA1-Tn7-like structure. Other blaIMP genes were mainly located in class 1 integrons of plasmids (including IncN, IncA/C2, IncHI2/IncN, etc). The classical backbone of class 1 integrons consisted of two conserved regions and the middle variable region. The 5′ conserved segment included the intI1 gene. The 3′ conserved segment contained the qacEΔ1 gene and sul1 gene (19). The middle variable region was the most flexible part of the integron, harboring one or more gene cassettes. The specific classification of integrons was related to the composition of these gene cassettes. In687, In798, In722, In809, In823, and In823b were described in E. coli, with In722 (ISKpn22−) located on the IncN plasmid, and In722 (ISKpn22+) on the chromosome (Fig. 4A). We have further sorted out the following eight integron types containing the blaIMP-4 gene, including In586, In809, In823, In823b, In1377, In1456, In1460, and In1589 (Fig. 4B). In809 was identified with the structure of blaIMP-4-qacG2-aacA4-catB3-qacE△1-sul1 in various plasmids. In823 and In823b were usually discovered in the IncN plasmid. In these integrons, the blaIMP genes usually co-exist with one or more resistance genes, especially the aminoglycoside resistance gene aacA4. The majority of qacEΔ1 gene and sul1 gene are retained.
Genetic characterization of the blaIMP in Enterobacteriaceae. (A) Schematic representation of different class 1 integrons carrying the blaIMP gene in Escherichia coli. (B) Schematic representation of different class 1 integrons carrying the blaIMP-4 gene in Enterobacteriaceae. Arrows indicate transcription directions of genes. The blaIMP genes are shown in red, and other resistant genes on class 1 integrons were labeled in pink; mobile elements were drawn in blue. The regions with highly similar sequences were marked in gray (nucleotide identity of ≥ 95%).
Global surveillance of blaIMP-positive CRECs
To further monitor the global blaIMP-positive CRECs, we assessed all available 340 genomes of blaIMP-positive CRECs in GenBank (Table S3). These isolates were mainly from 17 countries, with Japan accounting for the highest proportion of 40.29% (137/340), followed by Australia 31.18% (106/340), and China 9.71% (33/340). The USA, UK, and Singapore also had some distribution, and other countries were scattered (Fig. 5A). Among the CRECs, 13 blaIMP variants were identified, with blaIMP-4 accounting for 39.41% (134/340), followed by blaIMP-6 (32.65%, 111/340) and blaIMP-1 (18.53%, 63/340). Specifically, blaIMP-6 was the dominant variant in Japan, accounting for approximately 73.72% (101/137), followed by blaIMP-1 (23.36%, 32/137). In Australia, the blaIMP-4 variant was predominant, representing 95.28% (101/106). In China, blaIMP-4 was also the main variant, accounting for 90.91% (30/33). We also found blaIMP-27 and blaIMP-64 CRECs with regional endemicity, which were isolated in the USA. From 2004 to 2010, few blaIMP-positive E. coli strains were reported. Between 2011 and 2021, the overall detection rate remained relatively high with fluctuations, dominated by blaIMP-1, blaIMP-4, and blaIMP-6 variants, while other variants emerged sequentially. Notably, the number of blaIMP-4 in Australia surged in 2012, and the number of blaIMP-6 in Japan surged in both 2013 and 2015 (Fig. 5B). Among the blaIMP-positive CRECs, clinical sources accounted for 77.35% (263/340), predominantly blaIMP-6 (39.16%, 103/263), blaIMP-4 (32.32%, 85/263), and blaIMP-1 (21.67%, 57/263). Animal sources accounted for 15.59% (53/340), with blaIMP-4 accounting for 86.79% (46/53), followed by blaIMP-27 and blaIMP-38, which had a strong correlation with animal origin. Regarding environmental sources, one E. coli strain carrying both blaIMP-1 and blaNDM-5 genes was detected in a river in China in 2018, and another seven E. coli strains with blaIMP-64 all originated from housing environments of animals in the USA (Fig. 5B). This showed that E. coli in animals and environments can also serve as a reservoir for blaIMP resistance genes.
Global surveillance of blaIMP-positive CRECs. (A) Global phylogeographic distribution of 340 blaIMP-positive CRECs, including the nine isolates in our study. The different colored blocks represent various quantity levels. Proportional distribution of enzymatic phenotypes across six countries (number of blaIMP-positive CRECs ≥ 5). Thirteen IMP enzymes are indicated by distinct colors. (B) Timeline of blaIMP-positive CREC outbreaks in 17 countries. The bubble size was proportional to the number of strains. The bar chart illustrates the quantities and proportions of the 13 enzymes. A circular plot displayed the distribution of 340 blaIMP-positive CRECs across various sources and proportions of the 13 enzymes. NA, unavailable relevant information.
Comparison of antimicrobial resistance genes, virulence factor genes, and plasmid replicons from different countries
Principal coordinate analysis (PCoA) was conducted to elucidate the distribution patterns of antimicrobial resistance genes (ARGs), virulence factor genes (VFs), and plasmids from the top 3 contributing countries of blaIMP-positive CRECs (Fig. 6A, C, and E). The results showed that the distribution of ARGs among isolates reported from China, Japan, and Australia clustered with relative independence and exhibited some diversity. In contrast, the distribution of VFs among isolates from the three countries showed similar patterns. Additionally, a comparable distribution of plasmids was observed between China and Japan. A Venn plot was used for co-existence analysis of ARGs, VFs, and plasmids from the three countries (Fig. 6B, D, and F). The blaIMP-positive CRECs from Australia carried the highest and most unique numbers of ARGs, VFs, and plasmids, and the three countries showed relatively high overlap in ARGs, VFs, and plasmids, especially VFs.
Distribution characteristics of ARGs, VFs, and plasmid replicons of blaIMP-positive CRECs in China, Japan, and Australia (the data are listed in Tables S4 to S6). (A) PCoA of ARGs across the three countries. (B) Venn diagram of ARG types among the three countries. (C) PCoA of VFs across the three countries. (D) Venn diagram of VF types among the three countries. (E) PCoA of plasmids across the three countries. (F) Venn diagram of plasmid types among the three countries. Each dot in the figure represents a strain. Different colors represent different countries.
ST and phylogenetic analysis of blaIMP-positive CRECs
We conducted further analysis to evaluate the nine strains within a broader global context, as well as to understand the global epidemic STs, evolutionary characteristics, and other features of 340 blaIMP-positive CRECs. Among these 340 blaIMP-positive CREC strains, 82 STs have been identified, while remained 10 strains have untyped. We selected STs with an isolation count ≥5, as well as those widely reported in the literature with high resistance or virulence, for analysis and mapping. Over the past 20 years, prevalent ST strains have shown dynamic changes. From 2004 to 2010, the number of strains was small, with ST131 strains isolated during this period. ST131 was relatively prevalent during 2011–2015, with a significant increase in 2015. ST216 and ST58 strains increased in 2012. From 2016 to the present, the overall prevalence rate of ST131 has been lower than before; meanwhile, ST38, ST69, ST95, ST167, ST354, ST448, ST648, ST1193, and others have also been isolated occasionally, with no dominant ST emerging (Fig. S2).
The phylogenetic tree of 340 blaIMP-positive CRECs can be roughly divided into seven evolutionary clades. The nine strains we collected were distributed across different evolutionary clades among the global blaIMP-positive E. coli. The first E. coli carrying the blaIMP-4 gene, isolated from a urine sample of an Australian patient in 2004, belonged to ST131 (Fig. 7A). ST131 strains clustered in the phylogenetic tree, belonging to clade II, accounting for 36.18% (123/340), and were the most common type. Most of these strains were mainly derived from Japan, China, and Australia, with scattered distribution in many other countries. ST131 strains were mainly derived from clinical patients, with the predominant blaIMP-6, blaIMP-1, and blaIMP-4 (Fig. 7B). ST216 was dominant in Australia and only distributed in the country. ST216 clustered into clade VII in the phylogenetic tree, which only carried blaIMP-4 and was isolated from animals in 2012. ST58 clustered into clade V in the phylogenetic tree, except one from the USA, others mainly isolated from wild animals of Australia in 2012. The variants in the ST58 strains were mainly blaIMP-4 and blaIMP-38, which differed by one amino acid substitution (Ser214Gly). It showed that local clone outbreak and blaIMP gene mutation occurred in ST58 strains. ST216 and ST58 were clones prevalent in specific regions, relying on localized transmission routes and possessing the ability to adapt to the local environment (Fig. 7A). It can be observed that ST10, ST34, and ST58 have been isolated from both clinical patients and animals, while ST167 and ST218 have been isolated from both patients and the environment (Fig. 7A). This shows that the transmission of resistant bacteria is multi-source and complex, which increases the risk of their spread and poses a major challenge to public health.
(A) Phylogenetic tree of 340 blaIMP-positive CRECs collected from this study and the NCBI database. Phylogroups (I–VII) represent the seven clones of blaIMP-positive CRECs. (B) The Sankey diagram illustrated the relationships between STs and national distribution, as well as between STs and different blaIMP variants, among 340 blaIMP-positive CRECs.
Correlation analysis of ARGs with VFs and plasmid replicons in blaIMP-positive CRECs
Overall, we identified 149 different resistance genes, 206 different virulence genes, and 67 plasmid types among 340 blaIMP-positive CRECs. There was a weak but statistically significant negative correlation between the number of resistance and virulence genes carried by each isolate (r = –0.2648; P < 0.0001 by Spearman’s rank correlation) (Fig. 8A). Intriguingly, there was no correlation (P = 0.2795) between the number of resistance and virulence genes in ST131 (Fig. 8C). In addition, we found a positive correlation between the number of ARGs and plasmid replicons in each isolate (r = 0.5205, P < 0.001) and in ST131 (r = 0.2390, P = 0.0078), respectively (Fig. 8B and D). These results indicated that the plasmids played an important role in the transmission of ARGs. We also conducted a correlation analysis between STs and the number of resistance genes, virulence genes, and plasmids. The results showed that compared with the high-risk epidemic ST131, ST58 and ST216 carried a greater number of resistance genes and plasmids (P < 0.05), with no statistical differences observed in other STs; ST69 and ST95 carried more virulence genes (P < 0.05), while no statistical differences were found in ST38, ST354, and ST648 (Fig. S3). These STs in blaIMP-positive E. coli require focused monitoring and management.
Correlation analysis of ARGs with VFs and plasmid replicons. (A) Spearman correlation analysis between ARGs and VFs. (B) Spearman correlation analysis between ARGs and plasmid replicons. (C) Spearman correlation analysis between ARGs and VFs in ST131. (D) Spearman correlation analysis between ARGs and plasmids in ST131.
DISCUSSION
In this study, we characterized phenotypic and genomic features of nine blaIMP-4-positive CRECs from Zhejiang, China, and first reported the high-risk ST1193 clone carrying blaIMP-4 gene. The nine multidrug-resistant strains showed resistant phenotypes, with polymyxin, tigecycline, and amikacin provided as last options for treatment. CREC5925 and CREC7387 were the same clone (ST1193), and CREC3005 and CREC3006 belonged to the same clone (ST973), while other strains showed high genetic variability with relatively scattered STs. However, the blaIMP-4-carrying IncN plasmids among different clones were highly consistent, indicating that the IncN plasmid mediated the horizontal transfer of the blaIMP-4 gene.
The IncN-type plasmid, known for its efficient conjugative properties, typically consists of four conjugative modules: oriT, a relaxase gene, the gene encoding the T4CP, and the gene cluster encoding the T4SS (20). In our collected strains, the IncN plasmid carried complete conjugation transfer system for self-transmission, with the pIMP-CREC7837 plasmid excluded due to IS26-mediated disruption of the virB2 gene (21, 22). IS26 not only helps spread resistant genes but can also stop their spread by inserting randomly.
The broad-host-range IncN plasmids carry diverse resistance genes (e.g., to ESBLs, carbapenems, quinolones) and are key vectors in spreading carbapenemase genes (like blaNDM, blaKPC, and blaIMP) among Enterobacteriaceae (e.g., Escherichia spp., Klebsiella spp., Citrobacter spp., and Proteus spp.) (23–27). Additionally, the efficient conjugation, stability, and low fitness cost of the IncN plasmids in our study underscore their role as effective blaIMP vectors. This is consistent with reports attributing blaIMP-1/6 outbreaks in Japan to highly conjugative IncN-pST5 plasmids (28), and with findings that IncN plasmids (e.g., pIMP-1495) exhibit greater stability than F or HI2 types and impose minimal fitness cost (25, 27), collectively highlighting their critical function in blaIMP dissemination among Enterobacteriaceae.
The evolutionary trajectory of IncN plasmids may involve fusion with other types (e.g., IncX3, IncR), likely mediated by IS26, which enhances their stability, transferability, and resistance gene cargo (29–31). This potential is underscored by the diverse plasmid backgrounds (67 types identified) among global blaIMP-CRECs, where conjugative plasmids like IncN, IncF, and IncHI2 are prevalent, facilitating blaIMP exchange and host range expansion.
Beyond plasmids, mobile genetic elements like class 1 integrons significantly facilitate blaIMP transmission. These integrons dynamically organize resistance gene cassettes, with expression levels influenced by cassette position relative to the promoter (19). In our sorted integrons, blaIMP-4 was located in the first resistance gene cassette and was more readily expressed than other resistance genes. It was presumably associated with the widespread use of carbapenem antibiotics in recent years. Conversely, in In798 and In722, blaIMP followed aacA4 possibly reflecting aminoglycoside-driven selection. This cassette flexibility allows integrons to adapt rapidly to antibiotic pressures (32), making them valuable markers for blaIMP epidemiology.
Additionally, the integrons are frequently flanked by transposons or insertion sequences. This genetic context promoted blaIMP mobility, enabling its entry into vast plasmid clusters. For instance, In809 in Australian gulls promoted blaIMP dissemination via conjugative plasmids (5), while In823 in our strains formed a mobilizable unit with IS6100, serving as a resistance gene dissemination platform.
The blaIMP-27 was chromosomally located within a Tn7-associated class 2 integron. In722 (ISKpn22−) was located on the IncN plasmid, while In722 (ISKpn22+) was located on the chromosome. Besides mediating the movement of integrons between plasmids and chromosomes (21), IS26 further drives integron evolution, as seen in pIMP-GZ1058 where it inactivated intI1, yet still facilitated resistance gene transfer. Similarly, p128379-IMP plasmid (MF344559.1) lacked the integrase intI1 but contained the IS26, which can also play a role in drug-resistant gene recombination, transfer, and expression (21). In summary, the integrons are recognized as the primary platform for the capture and expression of blaIMP, while transposons and IS elements (particularly IS26) facilitate the structural evolution and mobilization of the entire resistance unit.
In global blaIMP-positive CRECs, blaIMP-6, blaIMP-4, and blaIMP-1 were predominant variants. Japan, China, and Australia were the top 3 contributors. Australia carried the highest number of ARGs, VFs, and plasmids. There were differences in the composition of ARGs among the three countries, indicating the regional evolution and dissemination processes of ARGs were relatively independent. The Spearman analysis showed plasmids play important roles in mediating the spread of resistance genes. In addition, we found that there is a certain degree of complementarity between the number of ARGs and VFs in the blaIMP-positive CRECs, but ST131 is excluded. This suggests that bacteria need to maintain the balance of virulence and resistance to decrease a fitness cost (33, 34). But the high-risk epidemic ST131 has a flexible ability for adaptive evolution, with coevolution processes or independent acquisition pathways for its resistance genes and virulence genes (35–37). ST131 can possess both high drug resistance and virulence without excessively compromising its own survival. This trait may be attributed to the evolutionary advantages of this clone, which is also one of the reasons for it becoming a globally dominant clone.
MLST and phylogenetic analysis have revealed the genetic and evolutionary diversity of blaIMP-positive CRECs. At present, 82 STs have been identified, and the phylogenetic tree can be roughly divided into seven evolutionary clades. Obviously, ST131 was the most common type. Due to increased use of carbapenems, the number of ST131 blaIMP-positive CRECs began to increase around 2010, a trend that was consistent with the global spread of ST131 (38). In 2015, ST131 clone outbreaks may have occurred in Japan’s clinical setting, contributing to the prevalence of blaIMP-6. And the frequency of the IMP-6 phenotype was statistically significantly associated with usage of third-generation cephalosporins in Japan (39). ST131 blaIMP-positive CRECs were not only highly prevalent in certain regions but also widely distributed globally. Additionally, we found that the ST216 clone broke out in Australian silver gulls in 2012. A high incidence of IncHI2 plasmids carrying blaIMP-4 was identified in this outbreak. The IncHI2 plasmids were similar to those from Australian human-derived clinical isolates, posing severe environmental pollution and public health risk (5).
Notably, we first reported two high-risk ST1193 clones carrying the blaIMP-4 resistance gene. The sequence type is notable for its rapid rise to prominence across the globe over a short period of time and shares similar characteristics with ST131, mimicking the evolutionary trajectory of ST131 (38). An epidemiology study indicated that it was a significant cause of community-acquired urinary tract and bloodstream infections. ST1193 was the second most common AMR clone (behind ST131) in a geographically well-defined human population (38). ST1193 is universally resistant to fluoroquinolones due to quinolone resistance-determining region mutations and frequently associated with the ESBL gene (blaCTX-M-15), which encodes resistance to most penicillins and cephalosporins (38). The emergence of ST1193 strains carrying carbapenem-resistant genes on mobilizable plasmids further poses challenges for clinical treatment. Global multidrug-resistant high-risk clones, including ST38, ST131, ST167, ST405, ST410, ST648, and ST1193, are all distributed in blaIMP-positive CRECs (38). Additionally, the ST69, ST95, and ST131 have been confirmed closely associated with bloodstream infections (40). These pose potential challenges to antimicrobial stewardship and infection control. Appropriate monitoring measures should be implemented to prevent these high-risk clones, posing an increasingly serious threat to public health.
The limitation of this study is that, in NCBI, there are few records of strains isolated from 2022 to the present, making it impossible to reflect the prevalence of blaIMP-positive CRECs in recent years. This may be related to the following factors: some countries or regions may not have performed genomic sequencing of strains due to economic reasons; in addition, the lag in uploading sequencing information may have led us to underestimate the prevalence of blaIMP-positive CRECs.
Conclusion
This study mainly describes the phenotypic and genomic characteristics of E. coli harboring blaIMP-4-carrying IncN plasmids, which are responsible for the horizontal transfer of blaIMP−4 gene among clones in the local region. It further clarifies the important role of other mobile genetic elements, such as class 1 integrons, IS26, and transposons, in the transmission of blaIMP resistance genes. Furthermore, our global genomic epidemiological study has elucidated the spatiotemporal distribution of blaIMP-positive CRECs, providing a comprehensive understanding of their evolutionary trajectory and revealing high genetic diversity (82 STs), with ST131 predominant (36.18%). The regional evolution and dissemination processes of ARGs were relatively independent among China, Japan, and Australia. Correlation analysis indicated that plasmids play an important role in the transmission of ARGs. It is necessary to strengthen the monitoring of high-risk clones, such as ST131 and ST1193, reduce the resistance burden to carbapenem antibiotics, and prevent them from posing an increasingly serious threat to public health.
MATERIALS AND METHODS
Clinical isolation and resistance gene screening
From January 2017 to December 2024, a retrospective survey for 94 carbapenem-resistant E. coli isolates was identified from four tertiary hospitals in Zhejiang, China. Common carbapenemase genes (blaKPC, blaIMP, blaVIM, blaOXA-48, and blaNDM) were amplified by polymerase chain reaction (PCR), and the blaIMP-positive products were further whole-genome sequenced. Strain identification was performed using MALDI-TOF MS (bioMérieux, Marcy l’Etoile, France) (41).
Antimicrobial susceptibility testing
The MICs of the nine blaIMP-4-positive CRECs in this study were determined by the broth microdilution method, and the results using the breakpoints recommended by the 2025 Clinical and Laboratory Standards Institute guidelines for interpretation (42), except for tigecycline, which was interpreted according to the European Committee on Antimicrobial Susceptibility Testing guidelines (https://www.eucast.org/clinical_breakpoints). A total of 10 antibiotics were tested, including cefepime, ceftazidime, imipenem, meropenem, ertapenem, amikacin, ciprofloxacin, colistin, tigecycline, and ceftazidime-avibactam. E. coli ATCC 25922 was used as a quality control strain to ensure the accuracy of the experiment.
Conjugative transfer experiments and conjugation frequencies
To explore the transferability of plasmids harboring blaIMP-4, a conjugative transfer assay was performed, as previously described (43). First, monoclonal colonies of donor and recipient E. coli J53 bacteria were collected into 2 mL of Luria-Bertani (LB) broth (Sangon Biotech, Shanghai, China), reached the logarithmic growth phase, mixed at a ratio of 1:1, and cultured on MH plates with a filter membrane overnight at 37℃. Transconjugants were screened on MH agar containing sodium azide (200 µg/mL) and ampicillin (100 µg/mL). Finally, the transconjugants were confirmed by PCR, MALDI-TOF MS, and MIC results.
To calculate conjugation frequency, we immersed and mixed the above overnight-cultured filter membranes in 0.9% NaCl solution. Then, we diluted the mixture to the appropriate concentration gradient and spread it on two types of MH agar plates: one containing both ampicillin (100 µg/mL) and sodium azide (200 µg/mL), and another containing only ampicillin (100 µg/mL). The conjugation frequency was calculated by dividing the number of transconjugants (CFU/mL) by the number of donor strains (CFU/mL), as previously described (44).
Stability of the plasmid
To determine the stability of plasmids in transconjugants, a passaging experiment was performed in a non-antibiotic environment (45). Three monoclonal transconjugants were inoculated in 2 mL of LB broth without antibiotics at 37°C with constant shaking. Then, 2 µL of overnight culture was serially passed into 2 mL of fresh LB broth every day for 10 days. Overnight cultures were diluted to appropriate concentration gradients every 5 days and then spread on non-selective MH plates. Fifty monoclones were selected from each plate for PCR to verify whether the plasmid was lost.
Growth rate determination
We determined the growth rate of transconjugants to describe their fitness cost. Four monoclonal transconjugants were grown overnight. The overnight cultures, with a 1:100 dilution ratio in LB broth, were added to a flat-bottom 100-well plate; each overnight culture was repeated three times. The plates were incubated at 37°C with shaking at 200 rpm. The optical density of each culture at 600 nm was measured every 5 min for 20 h by Bioscreen C analyzer (Oy Growth Curves Ab. Ltd., Finland). The relative growth rate was estimated based on OD_600_ curves using an R script, as previously described (46). Both the growth curve and the area under the curve were derived from these OD_600_ measurements. Then, they were analyzed and visualized by GraphPad Prism version 9.
Whole-genome sequencing and genomic analysis
WGS was conducted using the Illumina HiSeq and Nanopore MinION platforms at Zhejiang Tianke (Hangzhou, China). Complete genome sequences were assembled using the hybrid assembly tool Unicycler 0.4.8 (47) and annotated using RAST (https://rast.nmpdr.org/). ResFinder v.4.6.0 and PlasmidFinder v.2.0.1 from the Center of Genomic Epidemiology (https://www.genomicepidemiology.org/services/) were used to identify antibiotic resistance genes and plasmid types, respectively. The Orit Finder website (https://bioinfo-mml.sjtu.edu.cn/oriTfinder/) was used to forecast the self-transfer or mobilizing capability of bacterial mobile genetic elements (20). BLAST from NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi/) was used to find similar sequences of plasmids and genes. Sequence comparisons were performed using BLASTn v.2.4.0 and visualized using Easyfig v.2.2.3 and Proksee (https://proksee.ca/). Heatmaps were utilized for the visualization of SNPs using ChiPlot (https://www.chiplot.online/).
Global phylogeographic, MLST, and phylogenetic analysis of blaIMP in E. coli
In a total of 486,101 E. coli strains from NCBI, we screened 340 non-redundant, fully assembled whole-genome sequences of blaIMP-positive CRECs, including nine strains in our study (https://www.ncbi.nlm.nih.gov/pathogens/isolates, accessed on 9 July 2025). Species were delineated based on an average nucleotide identity (ANI) >95% using FastANI (https://github.com/ParBLiSS/FastANI). A total of 340 CRECs harboring antibiotic resistance genes, virulence genes, and the Inc-type plasmid of the strain were screened and identified using ABRicate (https://github.com/tseemann/abricate). Multilocus sequence typing was performed using the MLST tool (https://github.com/tseemann/mlst). The phylogenetic tree was constructed using Roary and FastTree and further visualized using ChiPlot (https://www.chiplot.online/). Adobe Illustrator v.27.9.1 was used to map the global geographic distribution.
Statistical analysis
We performed principal coordinate analysis based on the Jaccard algorithm and constructed confidence circles at a 95% confidence level to evaluate the composition of ARGs, VFs, and plasmids in blaIMP-positive CRECs from China, Japan, and Australia. (https://www.chiplot.online/) (16, 17). The Venn plot was used on the online website (https://www.bioinformatics.com.cn/static/others/jvenn/example.html) for co-existence analysis of three countries. GraphPad Prism version 9 was utilized for Spearman analysis and visualization of ARGs, VFs, and plasmid replicons. An adjusted P-value <0.05 was considered significant. Absolute r-values indicated correlation strength: 0.5–1.0 (high), 0.3–0.5 (moderate), 0.1–0.3 (low), and <0.1 (no correlation) (48).
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tompkins K, van Duin D. 2021. Treatment for carbapenem-resistant Enterobacterales infections: recent advances and future directions. Eur J Clin Microbiol Infect Dis 40:2053–2068. doi:10.1007/s 10096-021-04296-134169446 PMC 8527571 · doi ↗ · pubmed ↗
- 2Meletis G. 2016. Carbapenem resistance: overview of the problem and future perspectives. Ther Adv Infect Dis 3:15–21. doi:10.1177/204993611562170926862399 PMC 4735501 · doi ↗ · pubmed ↗
- 3He W-Y, Lv L-C, Pu W-X, Gao G-L, Zhuang Z-L, Lu Y-Y, Zhuo C, Liu J-H. 2023. Characterization of an international high-risk Escherichia coli ST 410 clone coproducing NDM-5 and OXA-181 in a food market in China. Microbiol Spectr 11:e 0472722. doi:10.1128/spectrum.04727-2237166308 PMC 10269901 · doi ↗ · pubmed ↗
- 4Mahmoud NE, Altayb HN, Gurashi RM. 2020. Detection of carbapenem-resistant genes in Escherichia coli isolated from drinking Water in Khartoum, Sudan. J Environ Public Health 2020:2571293. doi:10.1155/2020/257129332612664 PMC 7306079 · doi ↗ · pubmed ↗
- 5Dolejska M, Masarikova M, Dobiasova H, Jamborova I, Karpiskova R, Havlicek M, Carlile N, Priddel D, Cizek A, Literak I. 2016. High prevalence of Salmonella and IMP-4-producing Enterobacteriaceae in the silver gull on Five Islands, Australia. J Antimicrob Chemother 71:63–70. doi:10.1093/jac/dkv 30626472769 PMC 4681372 · doi ↗ · pubmed ↗
- 6Li Y, Zhang Y, Sun X, Wu Y, Yan Z, Ju X, Huang Y, Zhou H, Wang Z, Wang S, Zhang R, Li R. 2024. National genomic epidemiology investigation revealed the spread of carbapenem-resistant Escherichia coli in healthy populations and the impact on public health. Genome Med 16:57. doi:10.1186/s 13073-024-01310-x 38627827 PMC 11020349 · doi ↗ · pubmed ↗
- 7Watanabe M, Iyobe S, Inoue M, Mitsuhashi S. 1991. Transferable imipenem resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother 35:147–151. doi:10.1128/AAC.35.1.1471901695 PMC 244956 · doi ↗ · pubmed ↗
- 8Riccio ML, Franceschini N, Boschi L, Caravelli B, Cornaglia G, Fontana R, Amicosante G, Rossolini GM. 2000. Characterization of the metallo-beta-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of bla(IMP) allelic variants carried by gene cassettes of different phylogeny. Antimicrob Agents Chemother 44:1229–1235. doi:10.1128/AAC.44.5.1229-1235.200010770756 PMC 89849 · doi ↗ · pubmed ↗