Hemolytic activity and antibiotic resistance profiles of Staphylococcus aureus isolates from clinical patients
Yen-Hsi Lai, Chih-Chen Kao, Min Yi Wong, Tsung-Yu Huang, Yu-Hui Lin, Chien-Wei Chen, Yao-Kuang Huang

TL;DR
This study examines the hemolytic activity and antibiotic resistance of Staphylococcus aureus in dialysis patients and others, revealing patterns that could improve treatment strategies.
Contribution
The study identifies distinct hemolysin gene and antibiotic resistance profiles in S. aureus isolates from vascular access and non-vascular access infections.
Findings
MRSA and MSSA isolates showed β- and γ-hemolytic phenotypes with hla and hld being the most common hemolysin genes.
Over 80% of isolates were ampicillin-resistant, with higher multidrug resistance in MRSA.
No significant differences were found in resistance patterns between vascular and non-vascular access isolates.
Abstract
Staphylococcus aureus is a common human pathogen that can cause vascular and skin infections, and patients undergoing hemodialysis are particularly susceptible to vascular access infections caused by S. aureus. Hemolysins are important virulence factors, and antibiotic resistance poses challenges for treatment. In this study, S. aureus isolates were collected from hemodialysis patients with vascular access, such as arteriovenous grafts, tunneled-cuffed catheters, and arteriovenous fistulas, as well as from non-vascular access infection (VAI) patients. The hemolytic phenotype and eight antibiotic susceptibility of these isolates were tested, and PCR was used to detect hemolysin (hla, hlb, hld, and hlgC/B) and antibiotic resistance genes (accA-aphD, tetM, and tetK). The results showed that methicillin-resistant S. aureus (MRSA) and methicillin-susceptible S. aureus (MSSA) isolates…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
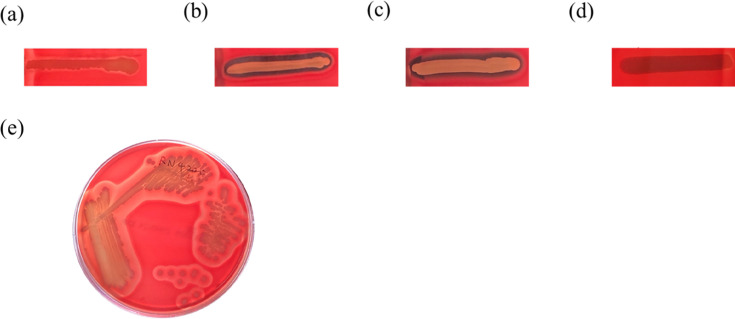 Fig 1
Fig 1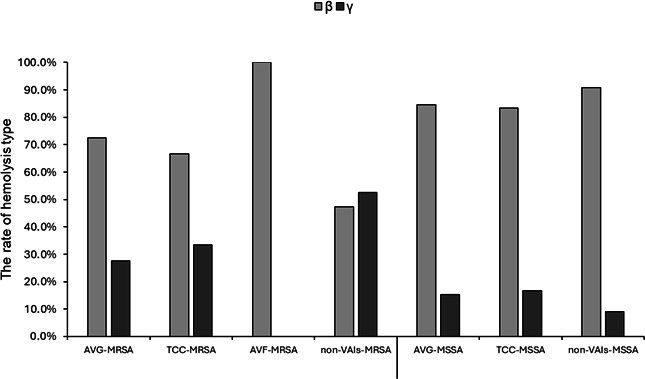 Fig 2
Fig 2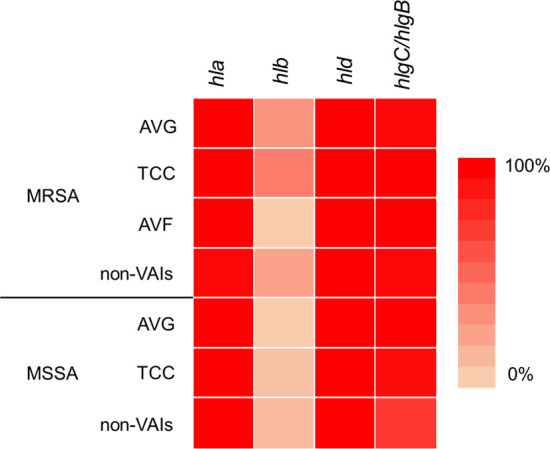 Fig 3
Fig 3| Type | Category | Hemolysis | Hemolysin gene(s) | No. |
|---|---|---|---|---|
| MRSA (102) | AVG (29) | β (21) | 8 | |
| 12 | ||||
| 1 | ||||
| γ (8) | 8 | |||
| TCC (33) | β (22) | 13 | ||
| 9 | ||||
| γ (11) | 11 | |||
| AVF (2) | β (2) | 2 | ||
| non-VAIs (38) | β (18) | 8 | ||
| 9 | ||||
|
| 1 | |||
| γ (20) | 20 | |||
| MSSA (77) | AVG (26) | β (22) | 22 | |
| γ (4) | 4 | |||
| TCC (18) | β (15) | 1 | ||
| 13 | ||||
| 1 | ||||
| γ (3) | 3 | |||
| Others (33) | β (30) | 3 | ||
| 18 | ||||
| 9 | ||||
| γ (3) | 3 |
| Drug | MRSA (102) | MSSA (77) | ||||||
|---|---|---|---|---|---|---|---|---|
| AVF ( | AVG ( | TCC ( | Non-VAI ( | AVG ( | TCC ( | Non-VAI ( | ||
| AMP | S | 0 (0) | 0 (0) | 0 (0) | 1 (2.63) | 3 (11.54) | 0 (0) | 7 (21.21) |
| I | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| R | 2 (100) | 29 (100) | 33 (100) | 37 (97.37) | 23 (88.46) | 18 (100) | 26 (78.79) | |
| DOX | S | 2 (100) | 22 (75.86) | 14 (42.42) | 21 (55.26) | 14 (53.85) | 14 (77.78) | 20 (60.61) |
| I | 0 (0) | 1 (3.45) | 5 (15.15) | 1 (2.63) | 4 (15.38) | 3 (16.67) | 3 (9.09) | |
| R | 0 (0) | 6 (20.69) | 14 (42.42) | 16 (42.11) | 8 (30.77) | 1 (5.56) | 10 (30.30) | |
| GEN | S | 2 (100) | 15 (51.72) | 15 (45.45) | 14 (36.84) | 20 (76.92) | 12 (66.67) | 23 (69.70) |
| I | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (3.85) | 0 (0) | 1 (3.03) | |
| R | 0 (0) | 14 (48.28) | 18 (54.55) | 24 (63.16) | 5 (19.23) | 6 (33.33) | 9 (27.27) | |
| RIF | S | 2 (100) | 26 (89.66) | 32 (96.97) | 30 (78.95) | 24 (92.31) | 16 (88.89) | 32 (96.97) |
| I | 0 (0) | 2 (6.90) | 0 (0) | 4 (10.53) | 2 (7.69) | 0 (0) | 1 (3.03) | |
| R | 0 (0) | 1 (3.45) | 1 (3.03) | 4 (10.53) | 0 (0) | 2 (11.11) | 0 (0) | |
| TET | S | 2 (100) | 22 (75.86) | 10 (30.30) | 18 (47.37) | 15 (57.69) | 11 (61.11) | 12 (36.36) |
| I | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| R | 0 (0) | 7 (24.14) | 23 (69.70) | 20 (52.63) | 11 (42.31) | 7 (38.89) | 21 (63.64) | |
| LZD | S | 2 (100) | 29 (100) | 32 (96.97) | 38 (100) | 26 (100) | 18 (100) | 33 (100) |
| I | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| R | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| TEC | S | 2 (100) | 29 (100) | 33 (100) | 38 (100) | 26 (100) | 18 (100) | 32 (96.97) |
| I | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| R | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| VAN | S | 2 (100) | 29 (100) | 33 (100) | 38 (100) | 26 (100) | 18 (100) | 33 (100) |
| I | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| R | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| MRSA (102) | No. (%) | MSSA (77) | No. (%) | |
|---|---|---|---|---|
| AVF (2) | 0 (0) | AVF (0) | 0 (0) | N/A |
| AVG (29) | 6 (20.7) | AVG (26) | 4 (15.4) | 0.612 |
| TCC (33) | 16 (48.5) | TCC (18) | 5 (27.8) | 0.151 |
| Non-VAI (38) | 16 (42.1) | Non-VAI (33) | 10 (30.3) | 0.303 |
| Type | Category | Genes | DOX | GEN | TET | DOX/TET | GEN/TET | DOX/GEN/TET |
|---|---|---|---|---|---|---|---|---|
| MRSA (102) | AVG (29) | 9 | ||||||
| 1 | 1 | |||||||
| 3 | ||||||||
| TCC (33) | 2 | 3 | ||||||
| 2 | 2 | 1 | ||||||
| 1 | 7 | |||||||
| 1 | ||||||||
| 2 | 2 | 1 | ||||||
| AVF (2) |
| – | – | – | – | – | – | |
| Non-VAI (38) | 7 | 1 | ||||||
| 2 | ||||||||
| 1 | 8 | |||||||
| 3 | ||||||||
| 2 | 1 | |||||||
| MSSA (77) | AVG (26) | 2 | ||||||
| 2 | ||||||||
| 3 | 3 | |||||||
| TCC (18) | 1 | |||||||
| 3 | 1 | |||||||
| 2 | ||||||||
| Non-VAI (33) | 1 | |||||||
| 1 | 7 | |||||||
| 5 |
- —Chiayi Chang Gung Memorial Hospitalhttp://dx.doi.org/10.13039/501100012283
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Bacterial biofilms and quorum sensing · Bacterial Identification and Susceptibility Testing
INTRODUCTION
Staphylococcus aureus is a common pathogenic bacterium in humans. It can cause blood vessels, skin infections, ulcers, and other diseases or deep and systemic infections such as osteomyelitis, endocarditis, pneumonia, and bacteremia. In addition, it can colonize the host without causing any disease (1). About 30% of the population carries this bacterium. Studies have found that dialysis patients are more likely to be infected with S. aureus than ordinary patients. The main reason is that S. aureus can adhere to host cells or matrix surfaces, such as medical devices, leading to cell aggregation and the production of toxins (2, 3). Dialysis equipment thus serves as a good site for bacterial attachment, and skin punctures at vascular access sites and the use of central venous hemodialysis and peritoneal dialysis polymer catheters are also factors that increase the risk of infection (4).
Nearly all S. aureus strains secrete diverse enzymes and cytotoxins such as hemolysins, nucleases, proteases, lipases, hyaluronidases, and collagenases and can also produce superantigens (SAgs), a family of potent immunostimulatory exotoxins that suppress host defenses against pathogenic strains. The primary function of these proteins may be to convert local tissues into nutrients needed for bacterial growth (5). Hemolysin plays an important role in the pathogenesis of all diseases caused by S. aureus by helping to lyse host cell membranes, disrupt or evade the immune system, and release nutrients for pathogen survival and disease development (6). Hemolysins can be divided into four types including α-, β-, δ-, and γ-hemolysins. α-Hemolysin, also known as alpha-toxin, is a cytotoxic, hemolytic protein that causes extensive damage to host cells including epithelial cells, endothelial cells, erythrocytes, monocytes, and keratinocytes, destroys cell membranes, and induces apoptosis. Encoded by the hla gene on the chromosome, it plays a role in many bacterial infections (5, 7). β-Hemolysin significantly affects the function of human immune cells. It is a sphingomyelinase encoded by hlb. It breaks down various types of cells according to the content of sphingomyelin, such as lymphocytes, keratinocytes, and neutrophils (6, 8). δ-Hemolysin is a low molecular weight exotoxin, a 26-amino acid peptide toxin produced by certain strains of S. aureus. It is encoded by the hld gene, which is part of RNAIII, an accessory gene regulator related to the phenolic soluble regulatory proteins (PSM), a family of peptide toxins. This toxin has multiple functions, including lysis of red blood cells, post-phagocytic neutrophils, and other mammalian cells, as well as toxic effects on other bacterial cells (5, 9). Moreover, β-toxin and δ-toxin exhibit strong hemolysis through synergistic effect (10). γ-Hemolysin is a pore-forming toxin (PFT) produced by S. aureus that lyses red blood cells in a variety of mammalian species (11). In addition, γ-toxin exerts toxic effects on phagocytes at the site of infection to evade the immune system. Furthermore, this hemolysin is encoded by the hlg gene (5, 7).
The invention of antibiotics has led to the emergence of drug-resistant strains, with S. aureus being particularly notorious for its resistance to penicillin. Epidemic clones often originate from one or a few successful lineages and have spread globally, and methicillin-resistant S. aureus (MRSA) is the most prominent example of these outbreaks (12). Its methicillin resistance is conferred by the mecA gene (13). In addition, S. aureus also possesses other antibiotic resistance genes, such as aminoglycoside (such as gentamicin) resistance, which is coded by aacA-aphD gene, and tetracycline (such as doxycycline and tetracycline) resistance, which is coded by tetK and tetM genes (14, 15). Drug-resistant strains have emerged, including multidrug-resistant strains that are resistant to three or more classes of antibacterial agents. These strains pose significant challenges for clinical treatment (16, 17).
This study mainly investigates the hemolytic types and hemolytic gene combination distribution of MRSA and methicillin-susceptible S. aureus (MSSA) in the vascular access infection (VAI) and non-VAI, as well as the status of antibiotic minimum inhibitory concentration (MIC) and antibiotic resistance genes, to provide reference for future treatment of patients in the renal dialysis pathway.
RESULTS
Detection of hemolytic phenotype among the collected S. aureus isolates on the sheep blood agar plate (BAP)
Hemolysis on blood agar is classified as β (clear zone, complete lysis), α (greenish or brownish zone, partial lysis), or γ (no lysis). Among the collected S. aureus isolates, only β- and γ-hemolytic phenotypes were observed. Even isolates showing only faint hemolysis were classified as β-hemolytic (Fig. 1a), while strongly β-hemolytic isolates displayed a distinct, transparent zone (Fig. 1b). The control strain produced β-hemolysin, which synergized with δ-hemolysin to enhance hemolysis at the intersection zone, also classified as β-hemolysis (Fig. 1c). γ-Hemolytic isolates showed no visible changes on the BAPs (Fig. 1d). The RN4220 strain, used as a quality control, exhibited only β-hemolytic activity (Fig. 1e).
Hemolysis phenotypes of S. aureus isolates from vascular access and non-VAI infections. (a–c) Exhibits the β-hemolysis, (d) displays isolates with γ-hemolysis, and (e) represents the S. aureus control strain.
Distribution of hemolysis types among MRSA and MSSA isolates
Among the 179 S. aureus isolates, 102 were identified as MRSA and 77 as MSSA. The majority of isolates exhibited hemolytic activity on blood agar, with β-hemolysis being more prevalent than γ-hemolysis and accounting for approximately or over 70% in most groups. In contrast to isolates from other infection sources, γ-hemolysis was observed more frequently than β-hemolysis among non-VAI MRSA isolates (Fig. 2).
Hemolysis phenotype profiles of S. aureus among collected isolates. The percentage of hemolysis types in MRSA and MSSA isolates.
Detection of hemolysis activity and hemolysin genes in S. aureus isolates
Four hemolysin genes were detected in S. aureus isolates, and the prevalence of these hemolysin genes was similar between MRSA and MSSA isolates. The majority of the collected S. aureus isolates carried the hla, hld, and hlgC/hlgB genes. Notably, the hld gene was present in 100% of both MRSA and MSSA isolates, followed by the hla gene. Only a small proportion of MRSA isolates and a few TCC-MSSA and MSSA isolates from non-VAI infections harbored the hlb gene (Fig. 3).
Heatmap of hemolysis genes in MRSA and MSSA isolates from vascular access infection (VAI) and non-VAI infections. AVG, arteriovenous graft; AVF, arteriovenous fistula; MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-sensitive Staphylococcus aureus; TCC, tunneled-cuffed catheter.
Detection of the hemolysin gene patterns in different hemolysis types among S. aureus isolates
MRSA and MSSA isolates were further categorized by infection source (VAI and non-VAI), and their hemolytic gene patterns were analyzed according to β-hemolysis and γ-hemolysis phenotypes. The predominant gene pattern was hla-hld-hlgC/hlgB, which was detected in 50.8% of MRSA isolates with β-hemolysis and 79.1% of MSSA isolates with β-hemolysis. Notably, this gene pattern was present in 100% of both MRSA and MSSA isolates with γ-hemolysis. The second most common gene pattern was hla-hlb-hld-hlgC/hlgB, accounting for 46% of MRSA isolates with β-hemolysis (Table 1). In all hemolysin gene patterns, the hld gene was revealed in the gene heatmap (Fig. 3) and indicated that this gene was the important factor in hemolysis among the collected S. aureus isolates.
Detection of MICs of antimicrobials and multiple drug resistance among S. aureus isolates
Eight antimicrobials were tested, arteriovenous fistula (AVF)-MRSA isolates were only resistant to ampicillin and susceptible to other antibiotics. Ampicillin resistance was observed in 99% (101/102) of MRSA isolates and 87% (67/77) of MSSA isolates. All MRSA and MSSA isolates were susceptible to linezolid, teicoplanin, and vancomycin, except for a few isolates that were not tested. Arteriovenous graft (AVG)-MRSA and tunneled-cuffed catheter (TCC)-MRSA isolates were equally resistant and susceptible to gentamicin. Most MRSA and MSSA isolates were susceptible to doxycycline and rifampicin, though a small number of isolates were resistant to these antimicrobials (Table 2). The proportion of antimicrobial resistance was higher in MRSA isolates than in MSSA isolates; however, for doxycycline and tetracycline resistance, the higher ratio of AVG-MSSA isolates was resistant to these agents compared to AVG-MRSA isolates. This trend was also observed between non-VAI MRSA and MSSA isolates resistant to tetracycline. The prevalence of multidrug resistance (MDR) was higher among MRSA isolates compared to MSSA isolates (Table 3). Furthermore, the MDR ratio of S. aureus with AVG infection was less than that of TCC and non-VAI infection. However, statistical analysis revealed no significant difference in MDR rates between MRSA and MSSA isolates (P value > 0.05).
Analysis of the correlation between antimicrobial gene patterns and antimicrobial resistance
There were five antibiotic resistance gene patterns among the collected S. aureus isolates. Gene patterns of accA-aphD and accA-aphD/tetM were prevalent among MRSA isolates (both 85.71%, 18/21), tetK (16.46%, 13/79), and accA-aphD/tetK (17.72%, 14/79) gene pattern were prevalent in the MSSA isolates. We found that 31.03% (9/29) AVG-MRSA and 18.42% (7/38) non-VAI-MRSA isolates were resistant to gentamicin and frequently carried the accA-aphD genes. Additionally, some isolates harbored both accA-aphD and tetM genes, showing resistance to doxycycline, gentamicin, and tetracycline, particularly among 21.21% (7/33) TCC-MRSA and 21.05% (8/38) non-VAI-MRSA isolates. In contrast, the tetK gene was commonly found in MSSA isolates resistant to tetracycline, with the accA-aphD/tetK gene pattern conferring resistance to gentamicin and tetracycline (Table 4).
DISCUSSION
S. aureus is a bacterium that possesses a variety of virulence factors that, individually or in combination, enable it to cause a wide range of diseases, including sepsis, infective endocarditis, pneumonia, eye infections, central nervous system infections, skin and soft tissue infections, and bone and joint infections (18). Hemolysin, leukotoxin (Panton-Valentine leukotoxin), and toxic shock syndrome toxin-1 contribute to the destruction of red blood cell membranes, impairment of phagocytic function of leukocytes, and induction of toxic shock syndrome, respectively (19). Among these toxins, hemolysins were classified as α-, β-, γ-, and δ-hemolysins, which are involved in the development of infectious diseases. α-, γ-, and δ-hemolysins are PFTs that induce lysis of target cells by forming pores in the cell membrane, whereas β-hemolysin (Hlb) is a non-pore-forming toxin (20). α-, β-, δ-, and γ-hemolysins were encoded by hla, hlb, hld, and hlg genes, respectively. Furthermore, S. aureus is known to exhibit a range of hemolytic phenotypes on blood agar, including α (partial), β (complete), and γ (none) hemolysis, consistent with previous studies (5, 7, 21). In our study, most of the collected S. aureus isolates exhibited hemolytic activity, with β-hemolytic isolates accounting for 61.8% of MRSA and 87% of MSSA. In many isolates, an expansion of the hemolytic zone was observed due to the interaction between β-hemolysin and δ-hemolysin, a phenomenon also reported in previous studies (22, 23). Regardless of variations in the presence of other hemolysin genes, hla and hld were the most frequently carried among S. aureus isolates (24–26). In our study, hla and hld were frequently detected in both MRSA and MSSA isolates, with hld being particularly prevalent. Similar findings regarding the widespread distribution of hld among staphylococcal strains have been observed in previous studies (27).
The results demonstrated that MRSA exhibited a higher prevalence of MDR compared to MSSA. Both MRSA and MSSA isolates were resistant to ampicillin, doxycycline, gentamicin, and tetracycline, a finding consistent with previous studies (28, 29). A correlation between antibiotic agents and resistance-associated genes was observed. In MRSA isolates, resistance was most frequently observed for gentamicin and doxycycline/gentamicin/tetracycline (both 17.64%, 18/102), corresponding predominantly to the accA-aphD and aacA-aphD/tetM gene patterns, respectively. In MSSA isolates, resistance to gentamicin/tetracycline (15.19%, 12/79) and tetracycline alone (12.66%, 10/79) was mainly associated with the aacA-aphD/tetK and tetK gene patterns, respectively. These findings are consistent with previous studies reporting that accA-aphD confers gentamicin resistance, while tetK and tetM are associated with tetracycline and doxycycline resistance (30, 31). Due to the impaired ability of the tetK protein to expel certain tetracycline derivatives, the tetK gene confers lower resistance to doxycycline (32), which is consistent with previous studies showing increased doxycycline susceptibility in S. aureus isolates harboring tetK or tetM (30, 31, 33). In our study, isolates harboring the tetK or tetM gene showed resistance to the doxycycline/tetracycline combination, but not to doxycycline alone.
Importantly, isolates from patients with VAI and those without showed largely similar experimental observations, with no notable differences in either hemolytic phenotypes or overall resistance across the eight tested antibiotics. Differences between MRSA and MSSA were mostly reflected in resistance gene patterns rather than in overall resistance.
Limitations of this study include its single-center design and relatively small sample size, which may affect generalizability. Nevertheless, these results provide insights into the hemolytic types and antibiotic resistance profiles of MRSA and MSSA isolates obtained from VAI and non-VAI patient specimens, supporting more targeted clinical management strategies.
MATERIALS AND METHODS
Bacterial isolates collection and identification
This research was conducted at Chiayi Chang Gung Memorial Hospital in Chiayi, Taiwan. A total of 179 S. aureus isolates were collected from patients with vascular access infections, including AVFs, prosthetic AVGs, TCCs, and other infections such as arthritis and pneumonia, between November 2013 and November 2023. The bacterial isolates were obtained from various sources, including blood, Hickman catheter tips, pus, tissue, wounds, and pleural effusion, and were cultured on BAPs. The isolates were identified using standard biochemical tests, including catalase and coagulase tests before 2019. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF) was used for identification after 2019. For routine culturing, the isolates were grown on tryptic soy agar and in tryptic soy broth under laboratory conditions. All isolates were stored in 15% glycerol stocks at −80°C.
Hemolysis phenotype analysis on the sheep BAP
Streak the center of the sheep BAP (Becton Dickinson, USA, trypticase soy agar with 5% sheep blood) with the RN4220 S. aureus strain that produces only beta-hemolysis. Test strains were streaked perpendicular to, but not touching, the central stripe, and the plates were incubated at 37°C for 24 h. After incubation, the hemolysis of the S. aureus isolates was observed, and the phenotypes on the SBA plates were recorded (7, 23, 34). α-Hemolysis: a greenish halo forms around the colonies, reflecting incomplete hemolysis. β-Hemolysis: cultures exhibit a translucent halo around and beneath the colonies, representing complete hemolysis. γ-Hemolysis: no noticeable zones are present around the colonies. Furthermore, the β-hemolysin produced by the control strain synergized with δ-hemolysin, resulting in enhanced hemolysis at the vertical intersection zone. This enhanced hemolysis was also classified as β-hemolysis (23, 35).
Hemolysin genes detection
In the hemolysis detection of bacterial isolates, α-, β-, δ-, and γ-hemolysins are encoded by hla, hlb, hld, and hlg genes, respectively. These four hemolysin genes, including hla, hlb, hld (36), and hlgC/hlgB (37) genes, were amplified utilizing the polymerase chain reaction (PCR) in previous studies.
MIC testing
This detection was conducted in accordance with a previous study (38). Bacterial isolates to be tested were streaked onto agar plates without inhibitors to obtain single colonies. The plates were incubated at 37°C for 18–24 h. For each isolate, three to five morphologically similar colonies were selected and transferred into a tube containing Mueller–Hinton broth for agitation (shocking) for 1–2 h. A 96-well sterile microtiter plate was labeled with the respective antibiotic concentrations, designating one row for each test, accommodating up to 10 different dilutions. Into columns 2–10, 50 µL of broth was added, while 100 µL of broth was added to the sterility control well (column 12) and 50 µL to the growth control well (column 11). Subsequently, add 100 µL of the antibiotic solution to column 1, then perform a twofold serial dilution of each antibiotic across the respective wells. For each bacterial isolate, the bacterial suspension was adjusted to a concentration of 1 × 10^8^ CFU/mL by vortexing (based on McFarland standard 0.5 or a optical density [OD] at 625 nm of 0.08–0.13) and diluted 1:100. Each well containing the antibiotic solution, along with the growth control well, was inoculated with 50 µL of the prepared bacterial suspension, achieving a final inoculum of 5 × 10^5^ CFU/mL. The microtiter plate was incubated at 37°C for 16–20 h. Overnight monitoring of the OD at 600 nm was performed using a 96-well plate reader. The MIC was defined as the lowest antibiotic concentration at which there was no visible bacterial growth (no turbidity observed with the naked eye) after 16–20 h of incubation. MIC standards and antimicrobial quality controls adhered to the guidelines of the Clinical and Laboratory Standards Institute (39).
Antimicrobial resistance gene detection
PCR was utilized to identify the isolates as MRSA, which exhibited oxacillin resistance and tested positive for the mecA gene using primers described previously (40). Additionally, the accA-aphD, tetK, and tetM genes, encoding aminoglycoside and tetracycline resistance, respectively, were amplified as described by a previous study (14).
Statistical analysis
The difference in the rate of MDR between MRSA and MSSA isolates was analyzed using the chi-square test in IBM SPSS Statistics for Windows, Version 22.0 (Armonk, NY: IBM Corp.). A P value of less than 0.05 was considered statistically significant.
Conclusion
This study investigated S. aureus isolates from both dialysis and non-dialysis patients. The isolates were classified into MRSA and MSSA, and their characteristics were analyzed according to VAI and non-VAI status. Isolates from VAI and non-VAI patients showed generally similar hemolysis patterns and antibiotic resistance profiles, suggesting comparable phenotypic traits regardless of dialysis status. MRSA isolates exhibited a higher proportion of MDR and their resistance gene profiles differed slightly in composition from those of MSSA. These findings highlight that while dialysis status may not strongly influence S. aureus phenotypes, the distinction between MRSA and MSSA remains critical. The phenotypic and genetic investigation of S. aureus isolates is crucial for infection management and the development of targeted treatment strategies.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Qiu Y, Xu D, Xia X, Zhang K, Aadil RM, Batool Z, Wang J. 2021. Five major two components systems of Staphylococcus aureus for adaptation in diverse hostile environment. Microb Pathog 159:105119. doi:10.1016/j.micpath.2021.10511934339796 · doi ↗ · pubmed ↗
- 2Seyyed Mousavi MN, Mehramuz B, Sadeghi J, Alizadeh N, Oskouee MA, Kafil HS. 2017. The pathogenesis of Staphylococcus aureus in autoimmune diseases. Microb Pathog 111:503–507. doi:10.1016/j.micpath.2017.09.02828919485 · doi ↗ · pubmed ↗
- 3Balaban N, Gov Y, Bitler A, Boelaert JR. 2003. Prevention of Staphylococcus aureus biofilm on dialysis catheters and adherence to human cells. Kidney Int 63:340–345. doi:10.1046/j.1523-1755.2003.00733.x 12472801 · doi ↗ · pubmed ↗
- 4Becker P, Hufnagle W, Peters G, Herrmann M. 2001. Detection of differential gene expression in biofilm-forming versus planktonic populations of Staphylococcus aureus using micro-representational-difference analysis. Appl Environ Microbiol 67:2958–2965. doi:10.1128/AEM.67.7.2958-2965.200111425708 PMC 92967 · doi ↗ · pubmed ↗
- 5Dinges MM, Orwin PM, Schlievert PM. 2000. Exotoxins of Staphylococcus aureus. Clin Microbiol Rev 13:16–34, doi:10.1128/CMR.13.1.1610627489 PMC 88931 · doi ↗ · pubmed ↗
- 6Divyakolu S, Chikkala R, Ratnakar KS, Sritharan V. 2019. Hemolysins of Staphylococcus aureus−an update on their biology, role in pathogenesis and as targets for anti-virulence therapy. Adv Infect Dis 9:80–104. doi:10.4236/aid.2019.92007 · doi ↗
- 7Al-Wahid ZHA, Al-Abbas MJA. 2023. Distribution of hla, hlb, hlg C, hld & cyl A hemolysin genes and their alleles in different bacterial species isolated from the variant clinical sources in Basrah. J Popul Ther Clin Pharmacol 30:180–198. doi:10.47750/jptcp.2023.30.03.021 · doi ↗
- 8Jia Y, Guan Z, Liu C, Huang M, Li J, Feng J, Shen B, Yang G. 2024. Staphylococcus aureus β-hemolysin causes skin inflammation by acting as an agonist of epidermal growth factor receptor. Microbiol Spectr 12:e 0222723. doi:10.1128/spectrum.02227-2338059627 PMC 10783061 · doi ↗ · pubmed ↗