Functional characterization of TGFβ/BMP receptors in Echinococcus granulosus sensu stricto: implications for parasite survival and therapeutic targeting
Liang Li, Ning Yang, Guodong Lv, Hui Wang, Jing Li, Tuerganaili Aji, Chuanshan Zhang, Renyong Lin

TL;DR
This study explores how TGFβ/BMP receptors in a tapeworm help it survive and suggests these receptors could be targeted for new treatments.
Contribution
The study identifies TGFβ/BMP receptors in Echinococcus granulosus s.s. as potential therapeutic targets for cystic echinococcosis.
Findings
EgTR1 and EgTR2 receptors are expressed during larval stages, with EgTR2 showing high transcription in activated protoscoleces.
Pharmacological inhibition of TGFβ/BMP signaling reduced parasite viability in vitro and cyst weight in vivo.
EgTR2 interacts with human BMP2, indicating a unique parasitic adaptation mechanism.
Abstract
Cystic echinococcosis (CE), a zoonosis caused by the larval stage of the tapeworm Echinococcus granulosus sensu lato (E. granulosus s.l.), remains a global health burden. Despite the conserved role of the transforming growth factor beta (TGFβ)/Smad signaling in cellular regulation across species, the molecular mechanisms of TGFβ/bone morphogenetic proteins (BMP) receptors in E. granulosus sensu stricto (E. granulosus s.s.) are poorly understood. The TGFβ/BMP receptors of E. granulosus s.s. were cloned and molecularly characterized. The positive binding interaction between receptors and their ligands was identified using the yeast two-hybrid system. To investigate the role of TGFβ/BMP signaling in E. granulosus s.s. development and survival, LDN193189 was employed in both the in vitro co-culture system and subsequent in vivo bioassays to investigate its pharmacological effects. Both…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
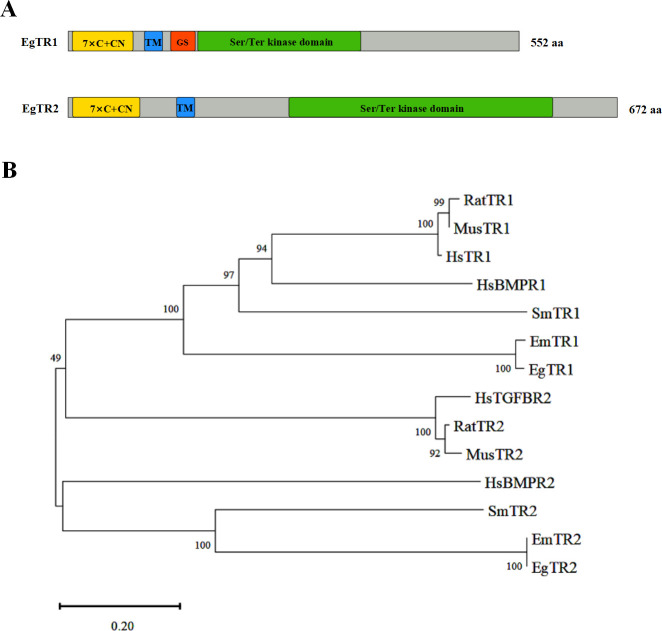 Fig 1
Fig 1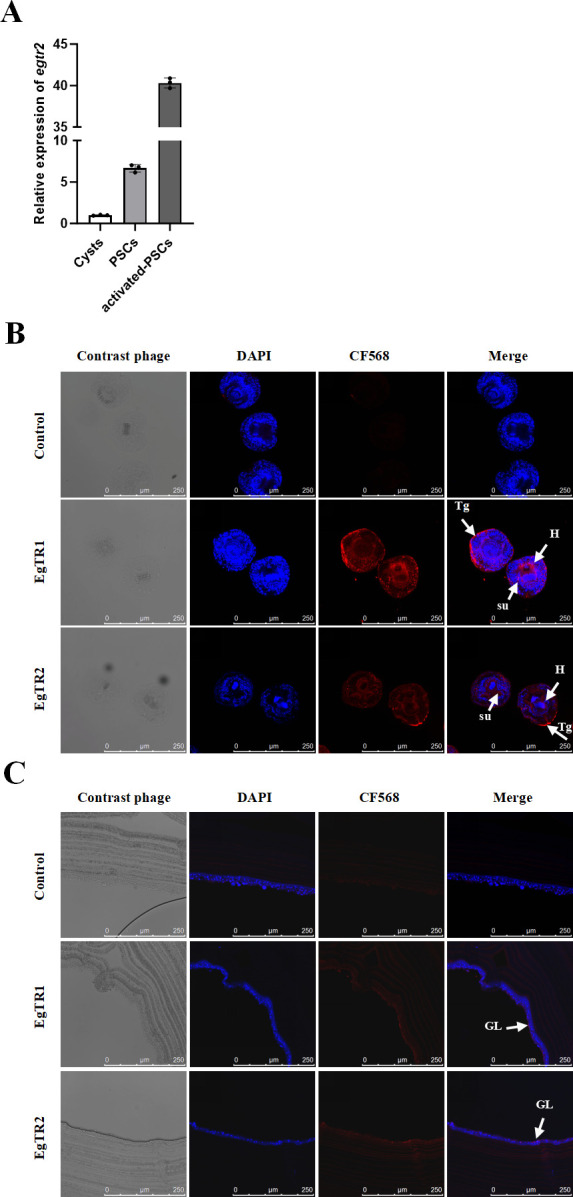 Fig 2
Fig 2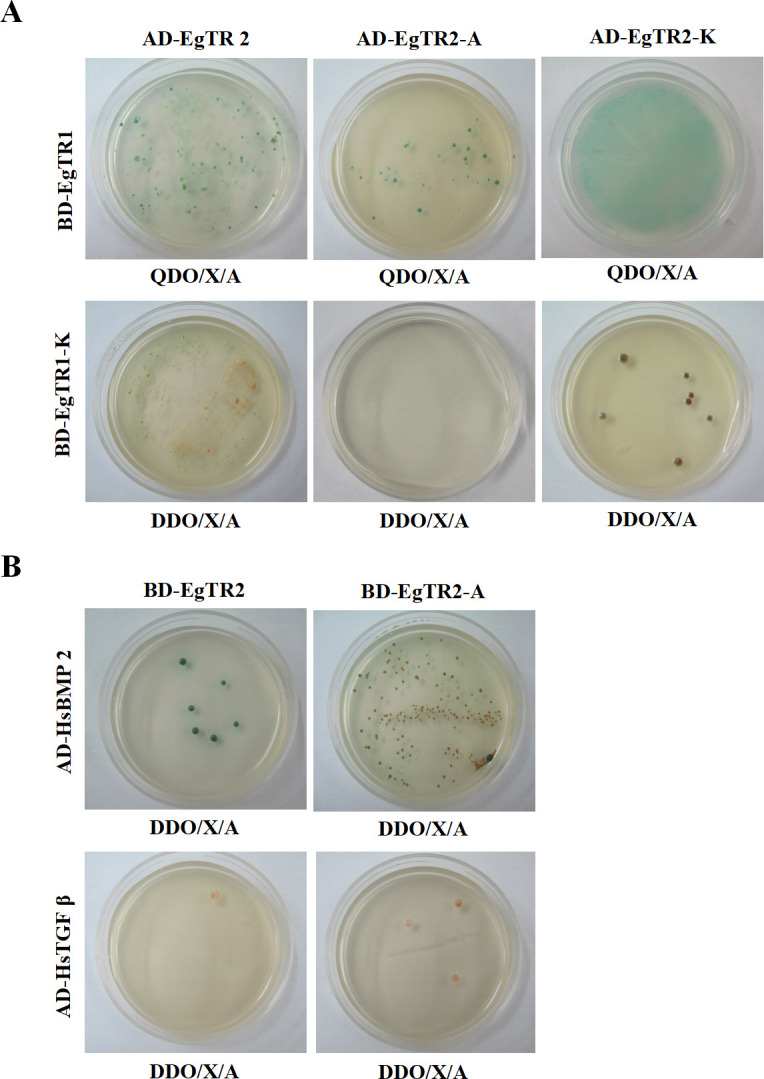 Fig 3
Fig 3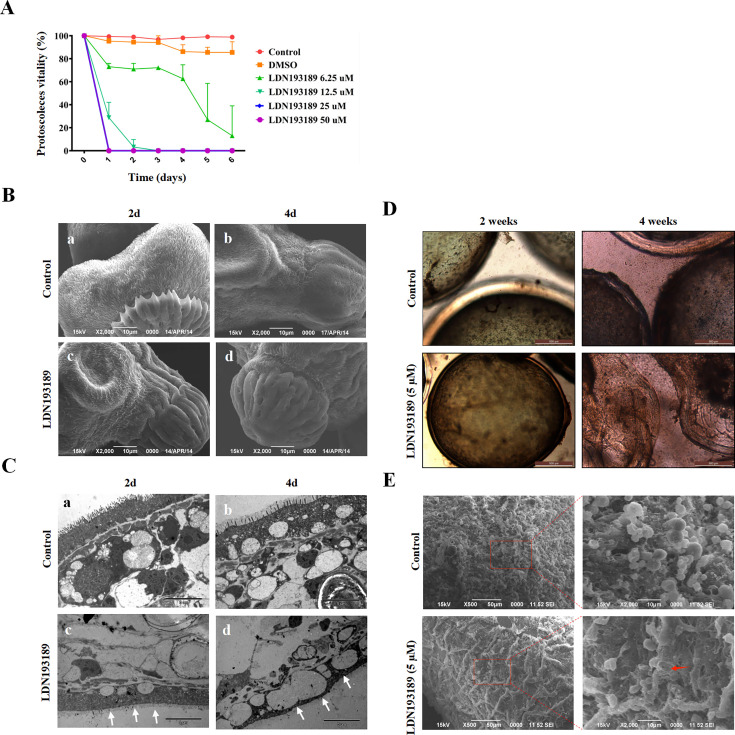 Fig 4
Fig 4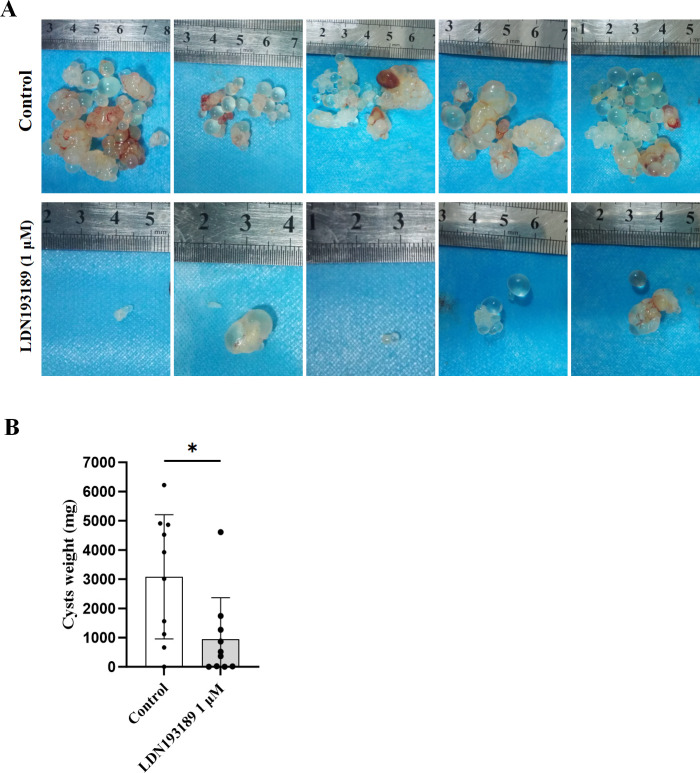 Fig 5
Fig 5| Vector | Growth with interaction partner | ||
|---|---|---|---|
| AD-EgTR2 | AD-EgTR2-A | AD-EgTR2-K | |
| BD-EgTR1 | + + | + | + + |
| BD-EgTR1-K | + | −− | + |
| BD-EgTR2 | BD-EgTR2-A | ND | |
| AD-HsBMP2 | + | + | ND |
| AD-HsTGF-β | −− | −− | ND |
- —Xinjiang Tianshan Project
- —Key program of Xinjiang Natural Science Foundation
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic infections in humans and animals · Echinoderm biology and ecology · Cancer Research and Treatments
INTRODUCTION
Cystic echinococcosis (CE) is caused by the larvae of the Echinococcus granulosus sensu lato (E. granulosus s.l.) (1). This zoonosis has a global distribution, with 5%–30% of the population in western China exposed to E. granulosus s.l. (2). CE poses a significant public health burden, with recent estimates of the global burden averaging 285,500 disability-adjusted life years (DALYs) for human CE. It is listed by the World Health Organization (WHO) as one of the 20 neglected tropical diseases, particularly impacting impoverished rural communities (1, 3). The life cycle of E. granulosus s.l. involves definitive hosts (dogs) and intermediate hosts (ungulates, predominantly sheep). Humans become infected through accidental ingestion of tapeworm eggs, leading to the development of fluid-filled metacestode larvae, primarily in the liver and lungs (4).
The transforming growth factor beta (TGFβ)/Smad signaling pathway is highly conserved across species, including nematodes, Drosophila, mice, and humans. This pathway regulates essential cellular processes such as proliferation, differentiation, adhesion, migration, and embryonic development (5). Components of the TGFβ/Smad pathway comprise TGFβ superfamily ligands, transmembrane serine/threonine receptors (type I/II), and intracellular Smad proteins. The TGFβ superfamily includes TGFβs, activins, inhibins, and bone morphogenetic proteins (BMPs) (6). The type I and type II transmembrane receptors share structural homology, each containing an extracellular ligand binding domain, an intracellular kinase domain and an intermediate transmembrane region. Upon activation, the TGFβ ligands bind to the type II receptor (TβRII) on the cell surface, leading to TβRI phosphorylation and subsequent intracellular signal transduction through Smad-dependent or -independent pathways, ultimately modulating transcription factor expression in the nucleus (7, 8).
The TGFβ/Smad signaling pathway has been implicated in parasite proliferation, differentiation, development, and survival. In Schistosoma mansoni (S. mansoni), host-derived TGFβ ligands activate the membrane-localized TβRII and TβRI receptors, which initiate downstream Smad signaling to regulate the expression of S. mansoni gynaecophoral canal protein (9). RNA interference-mediated knockdown of the activin receptor SmInAct disrupts embryogenesis of eggs, demonstrating its essential role in egg development (10). In echinococcus, five Smad homologs, including one Co-Smad (EmSmadD), two AR-Smads (EmSmadA, EmSmadC), and two BR-Smads (EmSmadB, EmSmadE), have been characterized and are phosphorylated by human TGFβ/BMP receptors (11–13). The TGFβ/BMP receptor family members of E. multilocularis, EmTR1-4, are activated by human TGFβ/BMP2 cytokines, leading to the phosphorylation of EmSmadB and EmSmadE (14). Furthermore, the *E. multilocularis-*derived activin-like cytokine (EmACT) drives metacestode brood capsule and protoscolex formation through the TGFβ/BMP signaling pathway (15).
Recent studies have identified components of the TGFβ/Smad pathway in E. granulosus sensu stricto (E. granulosus s.s.). Four Smad homologs, EgSmadA, EgSmadC, EgSmadD, and EgSmadE, have been identified and characterized (16–19). Among these, EgSmadE translocates to Mv1Lu cell nuclei upon treatment with hsTGFβ1 or HsBMP2 and has been shown to interact directly with EgSmadD by yeast two-hybrid and pull-down analyses (18). Despite the characterization of Smad effectors in E. granulosus s.s., the upstream receptors and their ligand specificity remain uncharacterized, hindering therapeutic development (20, 21).
This study characterizes EgTR1 and EgTR2 in E. granulosus s.s., determining their larval stages localization, protein-protein interactions, and kinase functions. We further evaluate the inhibitory and cytolytic effects of TGFβ/Smad pathway inhibitors using both in vitro and in vivo assays in E. granulosus s.s..
RESULTS
Molecular cloning and characterization reveal evolutionary conservation of TGFβ/BMP receptor homologs in E. granulosus s.s.
To identify TGFβ/BMP receptor homologs, specific primers were designed for amplifying sequences from E. granulosus s.s. Subsequently, two open reading frames (ORFs) cDNA fragments were amplified, and sequence analysis revealed 97.29%, 97.97% similarity to EmTR1 and EmTR2, respectively. The full-length ORF of EgTR1 (GeneBank accession number: CAJ43247) comprised 1,656 bp, encoding a 552 amino acid protein with a predicted molecular mass of 61.2 kDa (Fig. 1A). Computational algorithms predict that EgTR1 protein structure consists of a cysteine-rich receptor binding domain, a transmembrane domain, and a serine/threonine kinase domain with typical GS-box (motif SGSGSG) in the intracellular domain (Fig. S1). Similarly, the full-length ORF of EgTR2 (GeneBank accession number: PQ724305) comprised 2,016 bp, encoding a 672 amino acid protein with a predicted molecular mass of 74.2 kDa (Fig. 1A). The EgTR2 protein structure includes a receptor binding domain at the N-terminus, a transmembrane domain, and a serine/threonine kinase domain at the C-terminus (Fig. S2). The phylogenetic tree was constructed using the neighbor-joining method, incorporating TGFβ receptors and BMP receptors. Amino acid sequence alignments performed with DNAMAN software (Version 9.0) revealed high evolutionary conservation of EgTR1 and EgTR2. EgTR1 shared 96.93% identity with EmTR1, while EgTR2 exhibited 97.62% identity with EmTR2. (Fig. 1B).
Structural and phylogenetic analysis of E. granulosus s.s TGFβ/BMP receptors. (A) Domain architectures of EgTR1 (type I) and EgTR2 (type II) receptors. Color-coded functional domains include cysteine-rich extracellular regions (yellow; 7 conserved cysteines with CC-CN motif), transmembrane domain (blue), GS box (red orange), and serine/threonine kinase domain (green). (B) Unrooted neighbor-joining phylogenetic tree constructed using MEGA11 (v11.0.13) with 1,000 bootstrap replicates. Full-length protein sequences were aligned via ClustalW, demonstrating evolutionary relationships between EgTR homologs and vertebrate TGFβ/BMP receptors. GenBank accession numbers: E. granulosus s.s., EgTR1, CAJ43247; E. granulosus s.s., EgTR2, PQ724305; E. multilocularis, EmTR1, CAH56492; E. multilocularis, EmTR2, CAQ76822; S. mansoni, SmTR1, AAC16404; S. mansoni, SmTR2, AAV67030; M. musculus, MusTR1, AAA40495; M. musculus, MusTR2, AAK98605; R. norvegicus, RatTR1, AAA83216; R. norvegicus, RatTR2, NP_112394; H. sapiens, HsTR1, ACZ58375; H. sapiens, HsTR2, NP_001020018; H. sapiens, HsBMPR1, BAA19765; H. sapiens, HsBMPR2, KAI4037675.
EgTR1 and EgTR2 were involved in dynamic transcriptional regulation during E. granulosus s.s. development
The transcriptional expression levels of egtr2 transcripts were assessed through quantitative reverse transcription polymerase chain reaction (qRT-PCR) across larval development stages. The levels of egtr2 transcription varied among the three stages (activated PSCs, PSCs, and cysts), with the highest levels observed in activated PSCs and the lowest in cysts. The egtr2 transcripts were 40-fold more abundant in activated PSCs than in cysts and 6.6-fold higher in activated PSCs compared to the PSCs (Fig. 2A). Consistent with previous reports on egtr1 expression (18), these data highlight dynamic transcriptional regulation of TGFβ receptors during development. To further characterize the expression distribution of EgTR1 and EgTR2, an immunofluorescence analysis was conducted using mice anti-sera. The results showed that EgTR1 and EgTR2 were predominantly localized in the tegument tissue, sucker, and hooks in invaginated PSCs. Similarly, in in vitro cultured cysts, EgTR1 and EgTR2 were observed in the cytoplasm within the germinal layer. No fluorescence signals were observed in negative controls (Fig. 2B and C).
Subcellular localization of EgTR1 and EgTR2 in E. granulosus larval stages. (A) Quantitative PCR analysis was performed on total RNA isolated from cysts, protoscoleces (PSCs), and activated-PSCs. A bar graph comparing the relative expression levels (means ± SD) of egtr2 normalized to the levels of egelp. The relative expression value was averaged from triple samples, and the experiment was repeated three times. (B) Immunofluorescence localization in PSCs, and (C) in vitro-cultivated cysts. Sections were incubated with affinity-purified EgTR1-immunized mice sera and EgTR2-immunized mice sera, respectively, and F(ab’)2 goat anti-mouse antibody conjugated to CF568. Controls were incubated with preimmunized mice sera. Nuclei were stained with DAPI (4′,6-diamidino-2-phenylindole dihydrochloride, blue). Cells positive (CF568, red) for EgTR1 or EgTR2 are indicated by white closed arrows. Tg, tegument; H, hooks; su, suckers; GL, germinal layer; LL, laminated layer.
Yeast two-hybrid analysis revealed functional domain localization of EgTR2-EgTR1 interactions and host-parasite ligand binding
The yeast two-hybrid system was utilized to investigate the interactions between EgTR1 and EgTR2, as well as human TGFβ/BMP ligands. Bait and prey vectors were co-transformed into the yeast strain Y2H Gold, and interactions were assessed on selective media. pGADT7-EgTR2 (AD-EgTR2) and pGBKT7-EgTR1 (BD-EgTR1) transformants exhibited strong interactions under high-stringency conditions (synthetically defined [SD]/-Trp/-Leu/-Ade/-His/X-α-gal/AbA), while AD-EgTR2 and BD-EgTR1-K (the intracellular kinase domain) transformants displayed weak interactions under low-stringency conditions (SD/-Trp/-Leu/X-α-gal/AbA). Additionally, to investigate the host-parasite interactions, human TGFβ and BMP2 were cloned as fusions with the Gal4 activation domain, while EgTR2 and the extracellular ligand-binding activation domain (EgTR2-A) were fused to the Gal4 binding domain and co-transformed into the yeast strain Y2H Gold. The results indicated that AD-HsBMP2 interacted with both EgTR2 and EgTR2-A, as evidenced by the transformants grown under low-stringency conditions (SD/-Trp/-Leu/X-α-gal/AbA) (Fig. 3 and Table 1; Fig. S3).
Yeast two-hybrid analysis of interactions between EgTR1, EgTR2, and signaling partners. (A) Interaction analysis between EgTR1 and EgTR2 proteins. (B) Interaction analysis between EgTR2 and human homologs (HsBMP2, HsTGFβ). Translational fusion constructs were generated by cloning EgTR1, EgTR2, HsTGF-β, and HsBMP2 into the Gal4 activation domain (pGADT7 vector) or Gal4 DNA binding domain (pGBKT7 vector) vectors. Yeast double transformants (strain Y2HGold) were cultured on selective media for 5 days at 30°C. High stringency conditions: SD/-Trp/-Leu/-Ade/-His/X-gal AbA (QDO/X-α-Gal/AbA); Low stringency conditions: SD/-Trp/-Leu/X-gal AbA (DDO/X-α-Gal/AbA).
TGFβ/BMP pathway inhibitor LDN193189 inhibited the viability of E. granulosus s.s. in vitro
Previous evidence highlights the pivotal role of TGFβ/BMP signaling in echinococcus anterior pole morphogenesis and protoscolex development (22). To further validate the role of TGFβ/BMP signaling in the growth and development of E. granulosus s.s., PSCs and cysts were cultured in vitro with or without LDN193189. As illustrated in Fig. 4, PSCs exhibited striking dose-dependent susceptibility to LDN193189. Specifically, after 1 day of incubation, PSC viability was 95.28%, 73.13%, 28.76%, 0%, and 0% at LDN193189 concentrations of 0, 6.25, 12.5, 25, and 50 µM, respectively. By day 3, viability declined to 97.17%, 72.25%, 0%, 0%, and 0% at the same concentrations. On day 5, only 6.25 µM LDN193189 group retained partial viability to 26.97%, while all other treated groups showed complete lethality. In contrast, the untreated control group maintained >96% throughout the 5-day period (Fig. 4A). Morphological alterations in PSCs including tegumental alterations such as hooks loss and soma contraction observed via scanning electron microscope (SEM) (Fig. 4B). Transmission electron microscope (TEM) revealed ultrastructural changes such as microtriches shedding, cytoplasmic vacuolization, nuclear chromatin condensation, and the presence of residual lamellar bodies (Fig. 4C). For in vitro cultured cysts, LDN193189 treatment (5 µM for 4 weeks) induced structural collapse, including reduced turgor, germinal layer detachment from the laminated layer, and residual cellular debris (Fig. 4D). SEM confirmed micromorphological disruption, with the germinal layer fully detached from the acellular laminated layer. In contrast, DMSO-treated cysts retained intact architecture (Fig. 4E).
Effects of TGFβ/BMP receptor inhibitors on E. granulosus s.s. protoscoleces (PSCs) viability and hydatid cyst morphology. (A) Dose-dependent inhibition of PSCs viability. PSCs were treated with increasing concentrations of LDN193189 (6.25–50 µM) for 6 days. Cell viability was quantified using eosin Y exclusion assay (1%, wt/vol). Data represent mean ± SD of three independent experiments (n = 3). Controls: Untreated PSCs (Control) and 1% DMSO-treated PSCs (DMSO). (B and C) Ultrastructural alterations in PSCs. Morphological changes were analyzed using Scanning electron microscopy (SEM) (B) and transmission electron microscopy (TEM) (C) after incubation with either DMSO (control) or inhibitors 2, 4 days. (D) Cyst development inhibition. Cysts were incubated with 5 µM LDN193189 for 2 and 4 weeks, and the morphological changes of cysts were observed using light microscopy. (E) Ultrastructural changes of cysts detected by SEM after incubation with either DMSO (control) or inhibitors (LDN193189, 5 µM) 4 weeks. Red arrow indicates the ultrastructural changes.
LDN193189 pretreatment impairs E. granulosus s.s. cyst development in vivo
To assess the impact of TGFβ/BMP pathway inhibitors on PSCs, E. granulosus s.s. PSCs were pretreated to 1 µM LDN193189 in vitro for 30 days, followed by a mouse bioassay to assess the efficacy. Following 6 months after intraperitoneal inoculation with the pretreated PSCs, the mean cyst significantly decreased from 3,078.85 ± 2,017.05 mg (DMSO control) to 940.39 ± 1,349.65 mg (LDN193189-pretreated group, P = 0.0165). These findings demonstrate that pharmacological inhibition of the TGFβ/BMP pathway significantly impairs parasite survival and cyst formation in vivo (Fig. 5).
*In vivo efficacy of LDN193189 against E. granulosus s.s. cyst development in BALB/c mice. (A) Representative hydatid cysts harvested 6 months post-intraperitoneal inoculation with PSCs. Pretreatment conditions: 30-day in vitro exposure to 1 µM LDN193189 (test) or DMSO (control). (B) Quantitative analysis of cyst biomass (wet weight). Bar graph indicating the distribution of cyst weights (milligrams) in the different treatment groups (P = 0.0165). Data are presented as mean ± SD. Statistical comparisons between control and LDN193189 were performed using Welch’s unequal variances t-test (P ≤ 0.05).
DISCUSSION
The TGFβ/BMP signaling pathway, an evolutionarily conserved pathway, plays a critical role in modulating cellular proliferation, tissue morphogenesis, and organ development across species (23). Previous studies have elucidated the functional involvement of the TGFβ/Smad signaling cascade in diverse biological processes in schistosomes, including mating behavior, vitelline cell maturation, and egg embryogenesis (9, 24–26). Notably, in Echinococcus multilocularis, TGFβ/BMP signaling components are essential in anterior pole formation and PSC development (14, 15). While these pathways have been implicated as therapeutic targets for cancer and infectious diseases, the TGFβ receptors in E. granulosus s.s. remained uncharacterized.
In this study, we identified and functionally characterized two TGFβ receptors in E. granulosus s.s., designated as EgTR1 and EgTR2. Sequence alignment via BLASTp revealed that EgTR1 shares high homology with TGFβ receptor subgroup members in multiple species, particularly Schistosoma mansoni TGFβ receptors and E. multilocularis EmTR1. EgTR2, a novel type II receptor, exhibited close similarity to E. multilocularis EmTR2. Structural analysis demonstrated that both receptors possess a conserved extracellular domain featuring a Cys-box with seven flanking cysteine residues, coupled with an intracellular serine/threonine kinase domain. Notably, EgTR2 lacks the canonical GS-box characteristic of type I receptors, thus confirming its classification within the TGFβ/BMP type II receptor family.
Yeast two-hybrid assays demonstrated robust interactions between EgTR2 and both EgTR1 and HsBMP2. These findings align with prior reports of cross-species receptor-ligand interactions in E. multilocularis, where heterodimerization of EmTR1 with human bone morphogenetic protein type II receptor (BMPRII) activates BMP-regulated Smads (EmSmadB/EmSmadE) upon BMP2 stimulation (13, 14). Functional studies suggest that a single type II receptor can activate multiple type I receptors, enabling host-derived TGFβ/BMP ligands to initiate downstream Smad signaling (15). For instance, human TGF-β1 induces SmTR2-mediated activation of SmTR1 in schistosomes, promoting SmSmad2/SmSmad4 heterodimerization and subsequent regulation of gynecophoral canal protein (GCP) expression (9). In E. granulosus s.s., previous work identified the type I receptor EgTR1 and four Smad proteins (EgSmadA/C/D/E) (16–18, 27). Notably, EgSmadE undergoes nuclear translocation in vitro following exposure to human TGF-β1 or BMP2 (18). In this research, we observed elevated EgTR2 expression in activated PSCs compared to cysts or quiescent PSCs, which is consistent with the expression patterns of other TGFβ/Smad pathway components (18). Immunolocalization studies localized EgTR1 and EgTR2 to the tegument, sucker, and hooks of PSCs, and hydatid cyst germinal layer, suggesting their involvement in host-parasite interface dynamics, a phenomenon further supported by host-derived cytokine interactions (28–30).
Current CE management relies on surgical resection and benzimidazole (BZ) chemotherapy though BZ toxicity underscores the need for novel therapies. Kinase inhibitors have emerged as promising candidates (1, 31–35). We evaluated LDN193189, a BMP receptor kinase inhibitor, against E. granulosus s.s. PSCs. LDN193189 significantly reduced PSC viability and induced ultrastructural damage, including germinal layer detachment from the laminated layer. Additional inhibitors targeting specific pathways—WZB117 (GLUT1 inhibitor) and LDN212854 (BMPRI inhibitor)—demonstrated efficacy against E. granulosus s.s. and E. multilocularis, respectively (36, 37). These findings underscore inhibitors as potent anti-parasitic agents, offering novel strategies for CE treatment.
In conclusion, this study characterized two TGFβ/BMP signaling molecules of E. granulosus s.s. and examined the interaction of EgTR2 with EgTR1, and human BMP2, respectively. TGFβ/BMP receptor kinase would be a potential target for anti-parasitic drugs, such as LDN193189. Thus, further adjustments of treatment are to explore, such as optimal dosage and off-target effects require further investigation. In addition, new TGFβ/BMP signaling inhibitors offer starting points for the development of novel chemotherapeutic agents.
MATERIALS AND METHODS
Parasite material and in vitro cultivation
Protoscoleces (PSCs) were aseptically extracted from cysts of freshly slaughtered sheep infected with E. granulosus s.s. and cultured in vitro, as previously described (38). Activation of the PSCs using pepsin was conducted following the established method, which involved incubating the larvae in a low-pH solution (0.5 mg/mL pepsin in 0.85% [wt/vol] sodium chloride, pH 2.0, 37°C) for 30 min. The in vitro cultivation of cysts under axenic conditions was carried out using a previously established protocol (38).
Nucleic acid isolation, cloning, and sequencing
Total RNA was isolated from in vitro cultured PSCs and cysts, as described in a previous study (16). The cDNA was synthesized using the RevertAid first-strand cDNA Synthesis Kit (Thermofisher Scientific, USA) according to the manufacturer’s instructions. Based on the conserved regions of EmTR1 (GeneBank accession number: AJ841786.1, TGFβ receptor I of E. multilocularis) and EmTR2 (GeneBank accession number: FM178547.1, TGFβ receptor II of E. multilocularis), in conjunction with two homologous sequences in the supercontig database of E. granulosus (http://www.sanger.ac.uk/cgi-bin/blast/submitblast/Echinococcus), we identified two high-scoring segment pairs from E. granulosus s.l.. Specific primers used for EgTR1 and EgTR2 amplification were shown in Table S1. PCR products were cloned using the PCR cloning kit (TaKaRa, China) and then ligated to a pMD19-T Vector (TaKaRa, China). Positive clones were subsequently sequenced (Sangon Biotech Inc., China).
Gene structure and phylogenetic analysis
To identify the gene structures of egtr1 and egtr2, we analyzed the partial sequences available in the supercontig database of E. granulosus s.l. (http://www.sanger.ac.uk/cgi-bin/blast/submitblast/Echinococcus), using BLASTN with the egtr1 and egtr2 cDNA sequences. The phylogenetic analysis was constructed based on the predicted amino acid sequences of EgTR1, EgTR2, and other TGFβ/BMP receptors retrieved from GeneBank datebase (accessible via NCBI; https://www.ncbi.nlm.nih.gov) using the MEGA program (Version 11.0.13) as previously described (39). The transmembrane region of the conserved domain of EgTR2 was predicted using TMHMM (Version 2.0), and the conserved domain was identified using GenBank tools (40).
Expression analysis in larval developmental stages
Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was conducted to measure the gene transcriptional expression levels of egtr1 and egtr2 at various developmental stages. Total RNA extracted from E. granulosus s.s. PSCs and cysts were reverse transcribed, and the resulting cDNA was used as the templates for qRT-PCR amplification with SYBR Green PCR master mix (QIAGEN, Germany). Specific primers used for egtr1, egtr2, and egelp (used as an internal control) amplification are shown in Table S1. qRT-PCRs were performed in triplicate (final volume of 20 µL), on a CFX96 PCR system (Bio-Rad Inc.). The experimental protocol followed the manufacturer’s instructions, including an initial denaturation at 95°C for 2 min, 40 cycles of denaturation at 95°C for 15 s, annealing at 60°C for 30 s, melt curve analysis. Relative gene expression was calculated using the 2^−∆∆CT^ method with egelp as the normalization control.
Preparation of mouse anti-sera and immunolocalization analysis
The open reading frame (ORF) of EgTR1 (1,656 bp, 552 amino acids) and EgTR2 (2,016 bp, 672 amino acids) were cloned into pET-28α (+), respectively. Recombinant His-tagged proteins expressed in Escherichia coli BL21 were affinity-purified and used for mice immunization, following a previously established immunization strategy (21). Pre-immune mice sera served as negative controls. Eight weeks after the final immunization, the anti-sera were collected, and the serum titer was determined by enzyme-linked immunosorbent assay (ELISA), demonstrating high titers (>1:256,000 for EgTR1 and >1:512,000 for EgTR2) (Fig. S4).
Immunolocalization of EgTR1 and EgTR2 was performed on frozen sections of fresh PSCs and cysts following established protocols (18). The sections were fixed for 15 min, immersed in blocking buffer (10% BSA) for 30 min, washed with phosphate-buffered saline (PBS), and incubated overnight at 4°C with EgTR1 and EgTR2 anti-sera, respectively (1:200 dilution). Subsequently, the sections were washed with PBS and incubated with the fluorochrome-conjugated secondary antibody (Alexa Fluor 568, Thermo Fisher Scientific Inc., USA) for 2 h at room temperature in darkness. Nuclei were stained with 4,6-diamidino-diamidino-2-phenylindole (DAPI, Cell Signaling Technology, USA) for 5 min. Imaging was performed using a confocal laser scanning fluorescence microscope (Leica TCS SP8, Germany). The primary anti-sera were replaced by pre-immunized mice sera in the negative control.
Yeast two-hybrid analysis of EgTR1-EgTR2 interaction and ligand specificity
Yeast two-hybrid assays were used to assess the binding interaction between EgTR1, EgTR2, and hsBMP2. The full-length EgTR2, the extracellular ligand-binding activation domain (EgTR2-A, residues 1–128), and the intracellular kinase domain (EgTR2-K, residues 268–600) were cloned into the vectors pGADT7 (AD-EgTR2, AD-EgTR2-A, and AD-EgTR2-K), and pGBKT7 (BD-EgTR2, BD-EgTR2-A and BD-EgTR2-K), respectively. Similarly, HsTGFβ and HsBMP2 were cloned into the vector pGADT7 (AD-HsTGFβ and AD-HsBMP2). In parallel, full-length EgTR1 and its intracellular domain (EgTR1-K) were cloned into pGBKT7 (BD-EgTR1 and BD-EgTR1-K), respectively. The interaction assays were carried out by co-transformation of bait and prey plasmids into the yeast strain Y2HGold, following a previously described protocol (18).
In vitro effects of LDN193189 on E. granulosus s.s. viability
E. granulosus s.s. PSCs and cysts were obtained as previously described (18). Cysts with diameters of 2–4 mm were selected for in vitro experimental studies. Viable PSCs were seeded in 96-well cell culture plates, while cysts were seeded in 6-well plates and cultured in RPMI 1640 medium containing 10% fetal bovine serum, 100 U/mL penicillin, and 100 µg/mL streptomycin. The culture was maintained in a 5% CO_2_ atmosphere at 37°C.
After 3 days of normal culture, PSCs with viability exceeding 95% were divided into different groups: (i) DMSO group, treated with 0.2% (vol/vol) DMSO and (ii) inhibitor groups, treated with gradient concentrations of the BMP pathway inhibitor LDN193189 (50 µM, 25 µM, 12.5 µM, 6.25 µM, Selleck Chemicals, Shanghai, China). The PSCs were incubated at 37°C in 5% CO_2_, and the medium was replaced every 3–4 days. Viability of PSCs was assessed by 1% eosin staining combined with morphological analysis. For ultrastructural analysis, PSCs and cysts treated with LDN193189 were collected and examined using scanning electron microscopy (SEM) (LEO1430VP, ZEISS, Germany) and transmission electron microscopy (TEM) (JEM-1230, Hitachi, Japan) as previously described (22).
Assessment of E. granulosus s.s. PSCs' viability in vivo
To evaluate the effectiveness of LDN193189-treated PSCs, female BALB/c mice (6 weeks old, specific pathogen-free) were intraperitoneally injected 1,000 LDN193189-treated PSCs (pre-treated in vitro with 1 µM LDN193189). The control group received PSCs treated with PBS containing 0.2% DMSO. After 6 months of infection, the mice were euthanized, and the weight of parasite tissue was measured.
Statistical analysis
Results were expressed as mean ± standard deviation (SD) and analyzed using GraphPad Prism 10.0 (GraphPad Software, San Diego, CA). Continuous variables were analyzed by Student’s t-test (normally distributed data) or Kruskal-Wallis’s test (non-parametric data), with P < 0.05 considered statistically significant.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wen H, Vuitton L, Tuxun T, Li J, Vuitton DA, Zhang W, Mc Manus DP. 2019. Echinococcosis: advances in the 21st Century. Clin Microbiol Rev 32:e 00075-18. doi:10.1128/CMR.00075-1830760475 PMC 6431127 · doi ↗ · pubmed ↗
- 2Zhang W, Zhang Z, Wu W, Shi B, Li J, Zhou X, Wen H, Mc Manus DP. 2015. Epidemiology and control of echinococcosis in central Asia, with particular reference to the People’s Republic of China. Acta Trop 141:235–243. doi:10.1016/j.actatropica.2014.03.01424686096 · doi ↗ · pubmed ↗
- 3Casulli A. 2021. New global targets for NT Ds in the WHO roadmap 2021-2030. P Lo S Negl Trop Dis 15:e 0009373. doi:10.1371/journal.pntd.000937333983940 PMC 8118239 · doi ↗ · pubmed ↗
- 4Woolsey ID, Miller AL. 2021. Echinococcus granulosus sensu lato and Echinococcus multilocularis: a review. Res Vet Sci 135:517–522. doi:10.1016/j.rvsc.2020.11.01033246571 · doi ↗ · pubmed ↗
- 5Massagué J, Sheppard D. 2023. TGF-β signaling in health and disease. Cell 186:4007–4037. doi:10.1016/j.cell.2023.07.03637714133 PMC 10772989 · doi ↗ · pubmed ↗
- 6Wrana JL, Attisano L, Wieser R, Ventura F, Massagué J. 1994. Mechanism of activation of the TGF-β receptor. Nature 370:341–347. doi:10.1038/370341 a 08047140 · doi ↗ · pubmed ↗
- 7Derynck R, Zhang YE. 2003. Smad-dependent and Smad-independent pathways in TGF-β family signalling. Nature 425:577–584. doi:10.1038/nature 0200614534577 · doi ↗ · pubmed ↗
- 8Kang JS, Liu C, Derynck R. 2009. New regulatory mechanisms of TGF-beta receptor function. Trends Cell Biol 19:385–394. doi:10.1016/j.tcb.2009.05.00819648010 · doi ↗ · pubmed ↗