Prevotella copri facilitates wound healing in mice through the sphingosine-CerS1-ceramide metabolic pathway
Meili Zhao, Yue Liu, Shuyao Lv, Taotao Mi, Nan Wang, Shuaishuai Zhang, Hailiang Liu

TL;DR
A specific gut bacterium, Prevotella copri, helps heal wounds in mice by influencing a key metabolic pathway involving ceramide production.
Contribution
Prevotella copri is identified as a key wound-healing bacterium through its sphingosine-CerS1-ceramide pathway, offering a new precision therapy strategy.
Findings
Prevotella copri accelerates wound healing by upregulating pro-regenerative factors like vascular endothelial growth factor.
P. copri produces sphingosine, which is converted to ceramide via CerS1, promoting keratinocyte proliferation and blood vessel formation.
Pharmacological inhibition of CerS1 delays healing, confirming the pathway's role in wound regeneration.
Abstract
Skin wound repair constitutes a sophisticated biological process involving spatiotemporally coordinated molecular cascades, with emerging evidence highlighting the dynamic regulatory role of skin microbiota. Utilizing a broad-spectrum antibiotic (ABX)-treated murine model, we identified Prevotella copri as a core functional commensal in the wound microecosystem that orchestrates tissue regeneration through metabolite-host crosstalk. ABX-induced microbial remodeling significantly enriched P. copri relative abundance, accelerated wound closure, and upregulated pro-regenerative factors vascular endothelial growth factor and epidermal growth factor. Metabolomic profiling revealed that P. copri-secreted sphingosine undergoes bioconversion to C18-ceramide via the non-canonical CerS1 pathway, driving keratinocyte hyperproliferation and neoangiogenesis. Pharmacological inhibition of CerS1 with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
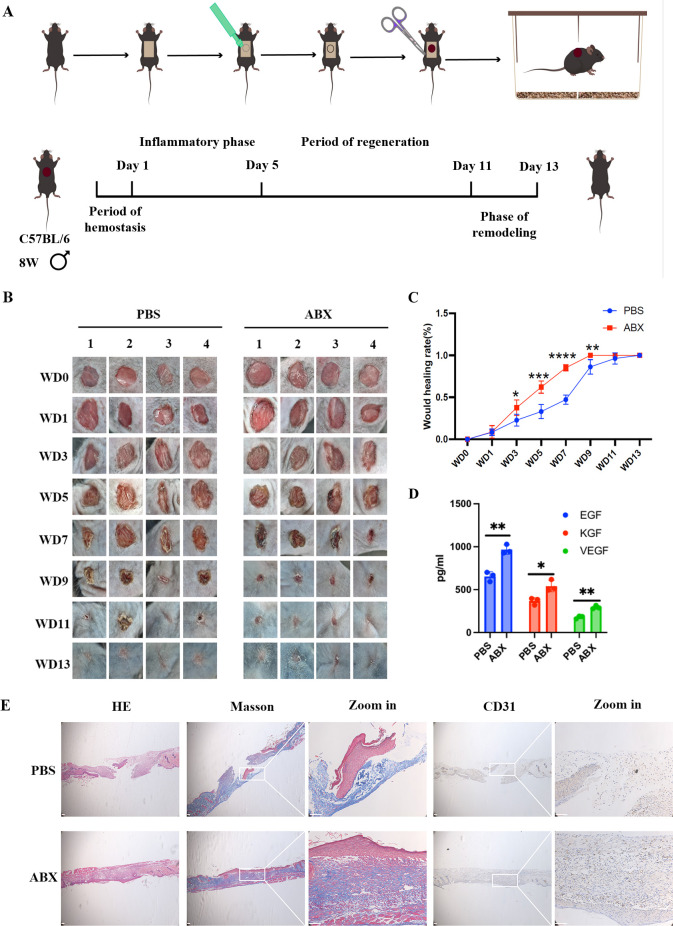 Fig 1
Fig 1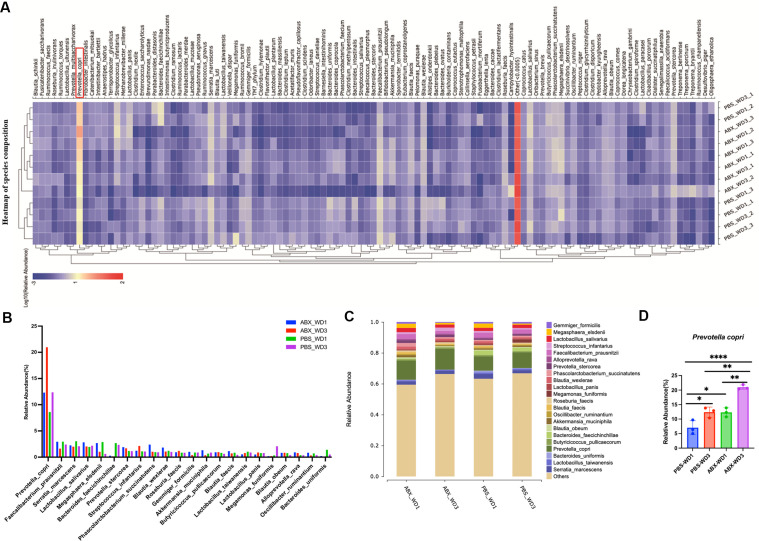 Fig 2
Fig 2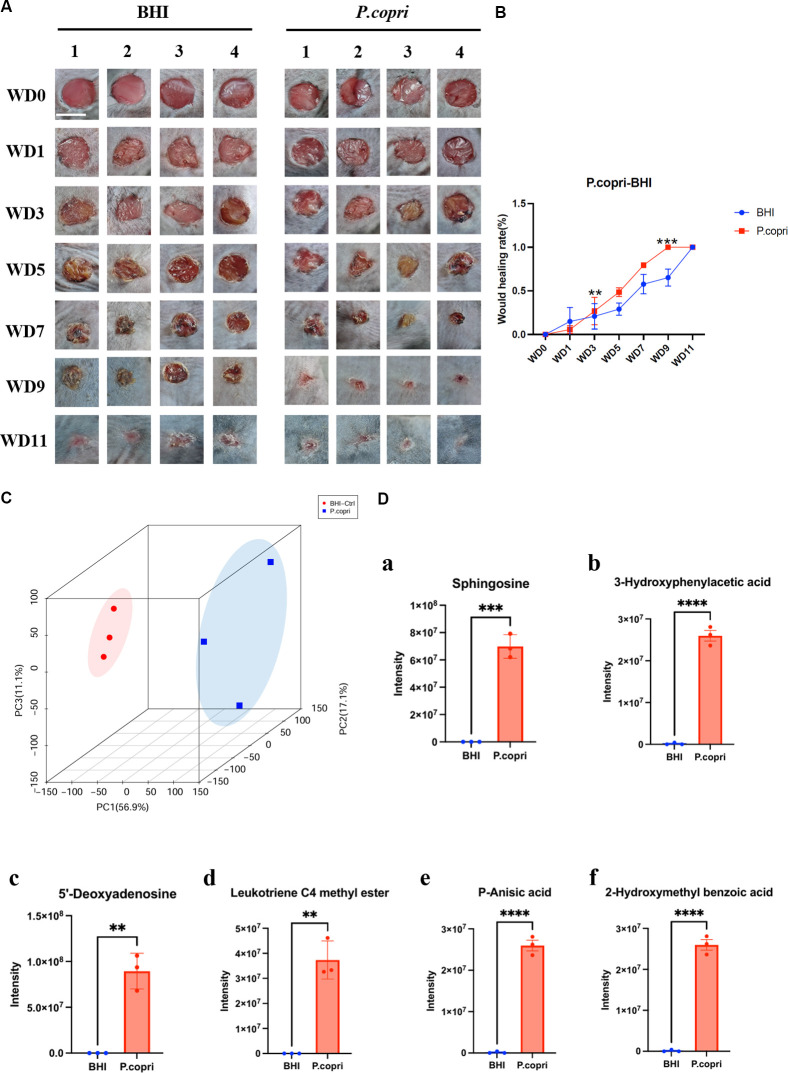 Fig 3
Fig 3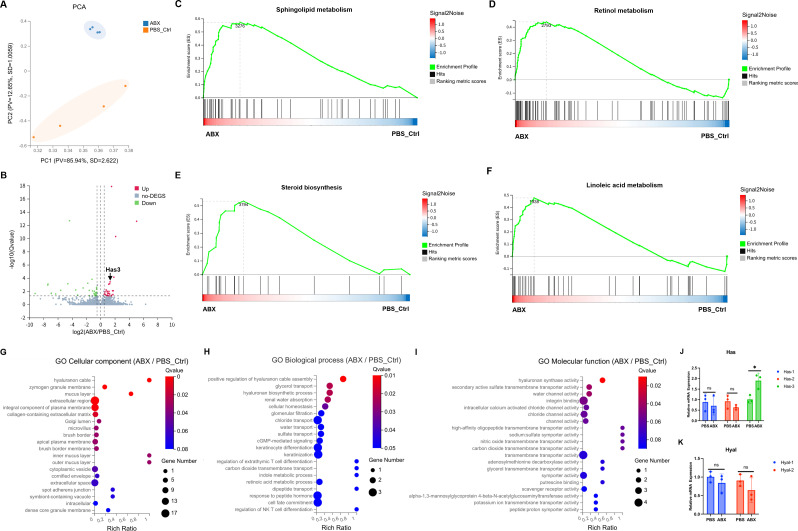 Fig 4
Fig 4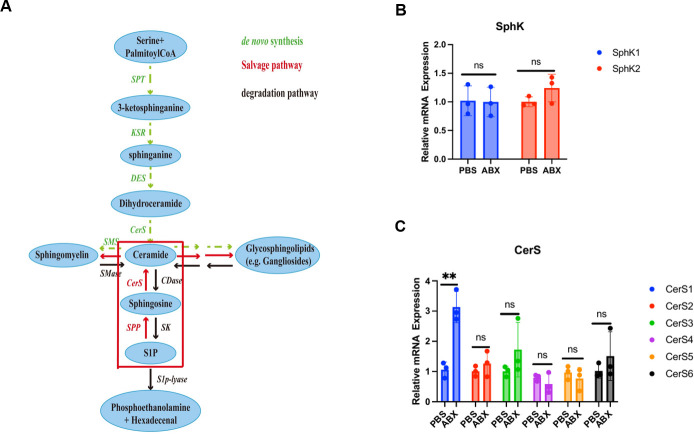 Fig 5
Fig 5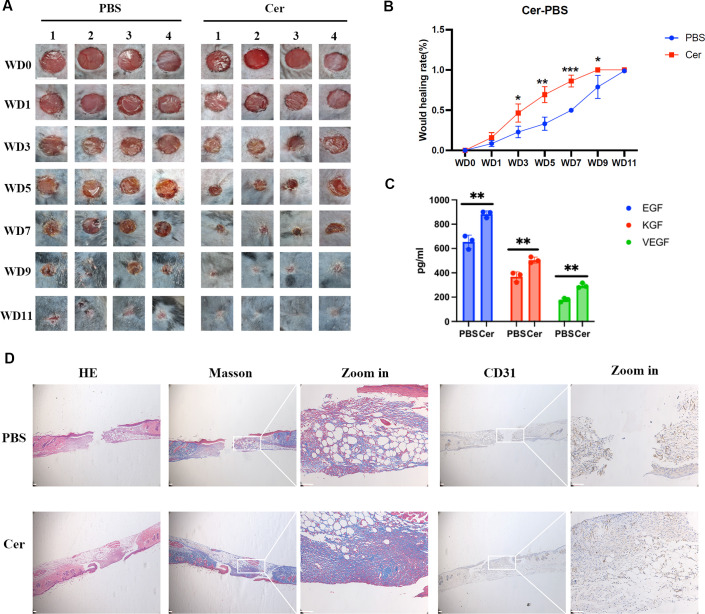 Fig 6
Fig 6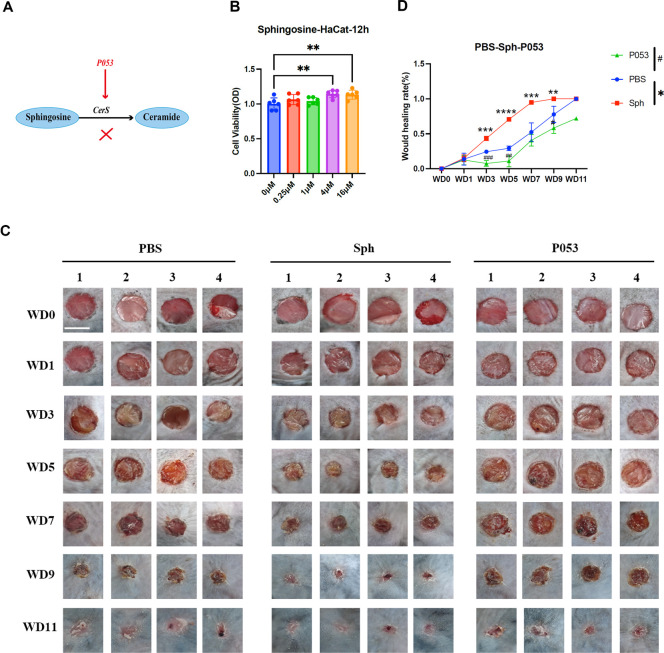 Fig 7
Fig 7- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —Fundamental Research Funds for the Central Universitieshttp://dx.doi.org/10.13039/501100012226
- —Natural Science Foundation of Shanghai under the Shanghai Action Plan for Science, Technology and Innovation
- —The Tongji University Medicine-X Interdisciplinary Research Initiative
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWound Healing and Treatments · Sphingolipid Metabolism and Signaling · Bacterial biofilms and quorum sensing
INTRODUCTION
The skin, the largest organ of the human body, serves as a critical barrier against the invasion of external pathogens. Diverse microorganisms—including bacteria, fungi, and viruses—colonize the skin surface, forming a complex and dynamic microbiota. Notably, skin microbes, particularly those inhabiting wounds, rarely exist in a free-living planktonic state but instead assemble into structured biofilm communities (1–3). These biofilms confer enhanced resistance to environmental stressors and antimicrobial agents (4, 5), fundamentally shaping the ecological dynamics and functional impact of the skin microbiota.
Prevotella copri, an obligate anaerobe, persists on oxygen-exposed skin by occupying hypoxic niches such as deep hair follicles (>200 µm depth), where restricted oxygen diffusion creates localized anaerobic environments (6). Its survival is further facilitated through integration into polymicrobial biofilms, wherein oxygen consumption by peripheral bacteria maintains an anoxic core (4). Although P. copri is typically low in abundance on skin compared to the gut, its presence has been reliably detected via metagenomic sequencing, circumventing the limitations of aerobic cultivation (7).
Wound healing proceeds through four highly coordinated and overlapping phases: hemostasis, inflammation, proliferation, and remodeling. The process initiates with hemostasis, characterized by fibrin clot formation mediated by platelet activation (8, 9). This is followed by an inflammatory phase involving neutrophil and monocyte infiltration, which clears debris and primes the tissue for repair via tightly regulated cytokine signaling (10, 11). The proliferative phase is driven by three central mechanisms: re-epithelialization, facilitated by keratinocyte migration under the guidance of keratinocyte growth factor (KGF) and epidermal growth factor (EGF) (12–14); angiogenesis, stimulated by vascular endothelial growth factor (VEGF) and basic fibroblast growth factor, leading to new vessel formation (15–17); and matrix reconstitution, in which fibroblasts degrade the provisional matrix via matrix metalloproteinases and synthesize new extracellular matrix components such as type III collagen and proteoglycans (13, 18).
A subpopulation of fibroblasts differentiates into α-smooth muscle actin-positive myofibroblasts, which promote wound contraction (19, 20). During the final remodeling phase, type I collagen gradually replaces type III collagen, and the vasculature is refined through selective pruning (19, 21). Emerging evidence underscores the skin microbiota’s dual role in modulating healing. For instance, while Staphylococcus aureus can impair repair through the secretion of proteases and hemolysins (22), it also contributes to epithelial and neural regeneration via IL-1β-IL-1R-MyD88 axis and IL-17A/IL-17RA signaling, respectively (23, 24). Beneficial commensals support healing through multiple mechanisms, including TLR-mediated antimicrobial peptide induction (25–27), competitive suppression of pathogenic biofilms (28), short-chain fatty acid-dependent barrier reinforcement (29, 30), and immunomodulation via Treg/Th17 balance regulation (31).
These insights redefine the skin microbiota from a passive “pathogen reservoir” to a dynamic therapeutic interface. Current clinical practices—often reliant on indiscriminate microbial eradication—overlook functional heterogeneity among commensal species. To bridge this gap, developing spatiotemporal maps of microbial-host crosstalk and deciphering strain-specific mechanistic pathways—particularly regarding metabolite signaling—will catalyze precision microecological therapies. Such advances promise to revolutionize wound care, transitioning from empirical antimicrobial regimens to microbiota-reprogramming strategies that harness ecological resilience and metabolic virtuosity, ultimately reshaping the frontier of regenerative medicine.
RESULTS
Microbial regulation of cutaneous regenerative dynamics
To systematically investigate how microbiota regulates wound healing, we established a localized intervention model comparing broad-spectrum antibiotic (ABX)-treated wounds and PBS-treated controls (Fig. 1A). Wound healing assays revealed that ABX-induced microbial depletion significantly accelerated wound closure compared to controls (Fig. 1B and C). Transcriptomic analysis showed marked upregulation of pro-regenerative factors, including VEGF, EGF, and KGF, in the ABX group (Fig. 1D). Histological assessment further demonstrated enhanced tissue regeneration: hematoxylin and eosin (H&E) staining revealed restored epidermal-dermal integrity, and Masson’s trichrome staining showed increased and well-organized collagen deposition. In addition, immunohistochemical analysis of CD31 indicated markedly promoted neovascularization in ABX-treated wounds (Fig. 1E). Together, these results suggest that antibiotic-induced microbiota remodeling enhances wound repair through structural restoration, angiogenic activation, and upregulation of healing-promoting factors.
*Microbial modulation of cutaneous wound healing dynamics. (A) Schematic workflow of the murine excisional wound model. (B) Representative macroscopic images of 6 mm excisional wounds in ABX and PBS control groups across post-wounding days 0–13 (n = 4, scale bar: 5 mm). (C) The ABX-treated group exhibited accelerated wound healing, achieving complete closure by day 8, whereas the PBS control group healed at a significantly slower rate. (D) ELISA quantification of pro-regenerative growth factors in wound tissues at day 8 post-injury. (E) Histomorphometric analysis of day 8 wound tissues: hematoxylin and eosin (HE) staining illustrating epidermal-dermal reorganization, Masson’s trichrome highlighting collagen architecture, and CD31 immunohistochemistry mapping neovascularization (scale bar: 50 µm). Data represent mean ± SEM; statistical significance between groups was determined by two-tailed Student’s t-test (*P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001).
Elucidating microbial drivers of accelerated wound healing
Building on the observed acceleration of wound healing following antibiotic treatment, we hypothesized that remodeling of the microbial community—rather than passive pathogen removal—constitutes a key mechanism enhancing repair in ABX-treated wounds compared to PBS controls. To test this, we performed spatiotemporal 16S rRNA sequencing of the wound microbiota at critical inflammatory time points (24 and 72 h post-injury; Fig. 2A). Taxonomic profiling revealed Prevotella copri as the most abundant taxon in both groups during early healing (Fig. 2A). Divergent colonization patterns were observed between treatments: PBS controls showed gradual enrichment of P. copri, consistent with natural repair progression, while ABX treatment triggered rapid expansion of P. copri, leading to its dominance within 72 h (Fig. 2B through D). This pattern suggests a density-dependent regulatory relationship, where P. copri biomass correlates with the activation level of host regenerative pathways.
*Ecological succession of wound microbiota driven by Prevotella copri. (A) Temporal heatmap depicting microbial community dynamics during the inflammatory phase at 24 and 72 h post-injury. (B) Comparative analysis of the most abundant taxa, highlighting P. copri as the most abundant species across experimental groups. (C) Taxonomic composition profiles illustrating microbial structural shifts between ABX and PBS control wounds. (D) Quantitative validation of P. copri abundance at critical time points, demonstrating its accelerated expansion in ABX-treated wounds versus gradual enrichment in controls. Data represent mean ± SEM; statistical significance was determined by two-tailed Student’s t-test (*P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.0001).
These findings reposition P. copri as both an ecological organizer and a biochemical modulator in wound repair. Its colonization occurs in phases: initial niche establishment primes the wound microenvironment, followed by biomass accumulation that activates threshold-dependent healing mechanisms. This study moves beyond pathogen-centric perspectives and highlights microbial community engineering as a promising strategy for advancing regenerative therapies based on ecological principles.
Prevotella copri enhances wound healing via sphingosine-mediated metabolic and structural reprogramming
To elucidate the molecular mechanisms underlying the pro-regenerative effects of P. copri, we topically applied its secreted metabolites (10⁹ CFU/mL) to murine wounds and observed significantly accelerated wound closure (Fig. 3A and B), indicating a metabolite-mediated effect. Metabolomic profiling revealed distinct biochemical signatures in P. copri-treated wounds, with sphingosine identified as a central bioactive molecule, accompanied by anti-inflammatory metabolites (p-anisic acid, 4-hydroxyphenylacetic acid) and angiogenesis-modulating compounds (e.g., leukotriene C4 methyl ester; Fig. 3C and D; Table S1). Transcriptomic analysis further indicated systemic metabolic reprogramming in antibiotic-treated wounds (Fig. 4A and B), characterized by coordinated upregulation of pathways involved in sphingolipid, retinol, and linoleic acid metabolism (Fig. 4C through F). Notably, P. copri-derived sphingosine was correlated with activation of host sphingolipid pathways, suggesting a cross-kingdom signaling mechanism through which bacterial metabolites directly stimulate endogenous repair processes.
*Therapeutic efficacy and metabolic signature of Prevotella copri secretions in wound healing. (A) Macroscopic progression of 6 mm excisional wounds treated with P. copri supernatant versus BHI medium controls, captured longitudinally from day 0 to 11 (n = 4, scale bar: 5 mm). (B) Kinetic profile of wound closure rates demonstrating accelerated re-epithelialization in the P. copri group. (C) Three-dimensional principal component analysis plot revealing distinct metabolic clustering between treatment groups. (D) Identified bioactive metabolites mediating therapeutic effects (relative quantification). (a) sphingosine (ceramide precursor), (b) 3-hydroxyphenylacetic acid (anti-inflammatory), (c) 5´-deoxyadenosine (energy metabolism modulator), (d) leukotriene C4 methyl ester (angiogenic mediator), (e) p-anisic acid (microbial antagonism), and (f) 2-hydroxymethyl benzoic acid (matrix remodeling cofactor). Data represent mean ± SEM; statistical significance was assessed by two-tailed Student’s t-test (**P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001).
*Metabolic reprogramming and hyaluronic acid remodeling in wound microenvironments. (A) Principal component analysis chart derived from RNA sequencing data. (B) Volcano plot illustrating differential gene expression. (C–F) Gene set enrichment analysis revealed systemic activation of pro-regenerative pathways in antibiotic-treated wounds, including sphingolipid metabolism, retinol metabolism, steroid biosynthesis, and linoleic acid metabolism. (G–I) GO term analysis demonstrated coordinated upregulation of hyaluronic acid (HA)-related functions: enriched HA-rich extracellular matrix organization (cellular component), enhanced HA fiber assembly (biological process), and elevated HA synthase activity (molecular function). (J and K) Quantitative PCR validation confirmed microbiome-driven HA homeostasis regulation, with ABX treatment significantly upregulating HA synthase Has3 expression (P < 0.01) while maintaining stable levels of HA degradation enzymes Hyal1 and Hyal2 (ns, non-significant). Data represent mean ± SEM; statistical significance was assessed by two-tailed Student’s t-test (*P ≤ 0.05, *P ≤ 0.01; ns, not significant).
Concurrently, hyaluronic acid (HA) matrix remodeling was identified as an additional regenerative mechanism. ABX treatment significantly enriched pathways related to HA metabolism (Fig. 4G through I) and specifically upregulated Has3 (hyaluronic acid synthase), while expression levels of HA-degrading enzymes remained stable (Fig. 4J and K). This shift promoted net HA accumulation—a clinically established strategy for enhancing wound healing (32).
Together, these results support a dual mechanism whereby P. copri promotes wound repair through sphingosine-driven metabolic activation and HA-mediated matrix reinforcement, synergistically optimizing the wound microenvironment via microbial modulation of host biosynthetic networks. These findings position P. copri as a key microbial engineer capable of activating conserved mammalian repair pathways, highlighting the potential of targeted metabolite-based strategies to bridge microbial ecology and regenerative medicine.
Prevotella copri drives wound healing via a non-canonical sphingosine-CerS1-ceramide axis
Mechanistic investigation into the regenerative function of Prevotella copri revealed its specific ability to synthesize sphingosine, which activates host ceramide biosynthesis via CerS1-mediated metabolic reprogramming (Fig. 5A). Transcriptional and quantitative PCR (qPCR) analyses of sphingolipid pathway genes showed selective upregulation of CerS1 in ABX-treated wounds, while sphingosine kinases (SphK1/2) remained unchanged—a pattern indicative of non-canonical sphingosine signaling (Fig. 5B and C). This “downstream activation, upstream stabilization” profile suggests that microbially derived sphingosine bypasses typical phosphorylation routes and directly promotes CerS1-driven ceramide production.
*Non-canonical sphingosine metabolic rewiring in Prevotella copri-enhanced healing. (A) Schematic representation of sphingolipid metabolism pathways, highlighting the P. copri-mediated bypass of classical sphingosine phosphorylation. (B and C) qPCR validation of sphingosine pathway components in day 3 wounds: SphK1/2 expression remained stable (ns, non-significant), while ceramide synthase CerS1 exhibited 3.8-fold upregulation in ABX-treated wounds versus PBS controls (*P ≤ 0.01). Data represent mean ± SEM; statistical significance was assessed by two-tailed Student’s t-test.
Functional validation confirmed the therapeutic role of ceramide: topical application significantly accelerated wound closure (Fig. 6A and B) and upregulated expression of EGF, KGF, and VEGF (Fig. 6C). Histological examination revealed enhanced restoration of the epidermal-dermal junction, increased collagen density, and promoted neovascularization, supporting ceramide’s role as the central effector molecule in P. copri-mediated repair (Fig. 6D).
*Ceramide accelerates wound healing through structural and molecular remodeling. (A) Macroscopic progression of 6 mm excisional wounds treated with 0.781 µM ceramide (Cer) versus PBS controls, captured longitudinally from day 0 to 11 (n = 4, scale bar: 5 mm). (B) Kinetic profile of wound closure rates demonstrating Cer-mediated healing acceleration (ceramide dose: 0.781 µM). (C) ELISA quantification of pro-regenerative factors at day 8 post-injury, showing Cer-induced upregulation of EGF, KGF, and VEGF. (D) Histomorphometric analysis of day 8 tissues: hematoxylin and eosin (HE) staining revealing enhanced epidermal-dermal junction reconstruction, Masson’s trichrome demonstrating increased collagen density, and CD31 immunohistochemistry confirming elevated neovascularization (scale bar: 50 µm). Data represent mean ± SEM; significance was determined by two-tailed Student’s t-test (*P ≤ 0.05, **P ≤ 0.01, **P ≤ 0.001).
Critically, the necessity of CerS1 was established through pharmacological inhibition: co-administration of sphingosine with the CerS1 antagonist P053 completely abolished the healing acceleration, whereas sphingosine alone sustained pro-regenerative effects (Fig. 7A through D). This combination of gain-of-function (ceramide supplementation) and loss-of-function (CerS1 inhibition) definitively identifies the sphingosine-CerS1-ceramide axis as the core mechanism underlying the therapeutic action of P. copri.
*Pharmacological validation of the sphingosine-ceramide axis in wound repair. (A) Mechanistic schema illustrating P053-mediated inhibition of CerS1-driven ceramide biosynthesis, disrupting the Prevotella copri-initiated regenerative cascade. (B) CCK-8 assay revealed sphingosine-induced keratinocyte hyperproliferation (1.9-fold, P < 0.001) within 12 h, confirming its direct mitogenic activity. (C) Macroscopic progression of 6 mm wounds treated with sphingosine (4 µM) or co-administered with the CerS1 inhibitor P053 (1 µM), demonstrating complete abolition of healing acceleration by pathway blockade (n = 4, scale bar: 5 mm). (D) Time-course quantification of closure rates: sphingosine monotherapy enhanced healing, while P053 co-treatment reverted kinetics to baseline levels. Data represent mean ± SEM; significance was determined by two-tailed Student’s t-test (**P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001).
The discovery of this microbial-metabolic bypass reveals an evolutionary convergence between bacterial metabolite production and host lipid signaling pathways. By precisely delivering bioactive metabolites that modulate conserved host repair pathways, P. copri exemplifies how microbiota can functionally augment endogenous regenerative networks, offering new avenues for microbiome-based therapeutic strategies.
DISCUSSION
Ecological and functional role of P. copri in wound healing
As a keystone symbiont within the skin wound microecosystem, Prevotella copri exhibits unique ecological and functional specialization in orchestrating tissue regeneration. This anaerobic bacterium thrives in both ABX and PBS wounds, leveraging β-lactamase-mediated antibiotic resistance to establish ecological dominance—a trait that ensures its survival under antimicrobial pressure. Longitudinal analysis revealed its intrinsic repair role: while ABX intervention triggered rapid P. copri proliferation peaking at 72 h post-injury, control wounds displayed progressive enrichment, underscoring its fundamental contribution to physiological healing beyond mere antibiotic selection. This “competitive release” dynamic—where microbiota restructuring alleviates ecological constraints—propels P. copri into a leadership role, accelerating repair through prioritized niche colonization.
The antibiotic-induced dominance of P. copri represents an artificial monoculture resulting from broad-spectrum depletion of competitors (e.g., vancomycin/neomycin eliminating gram^+^/gram^−^ facultative anaerobes), which bypasses natural wound succession, where anaerobes like Prevotella gradually emerge in hypoxic niches within polymicrobial biofilms (6).
Comparative mechanisms among commensals
While Prevotella copri is a well-established gut commensal (33), its detection on skin, though low in abundance, highlights the metabolic flexibility of this anaerobe and the capacity of skin microenvironments to support transient colonization. Our findings that P. copri promotes repair add to a growing list of beneficial wound microbes, including Staphylococcus epidermidis (34) and Lactobacillus rhamnosus GG (35–37). Notably, the mechanism we propose—dependent on bacterially derived sphingosine—is distinct from the sphingomyelinase activity of S. epidermidis or the lysate-driven migration induced by L. rhamnosus, underscoring the diverse strategies microbes employ to influence host repair. Furthermore, it is well-established that different commensals employ distinct mechanisms to interact with the host. For instance, Bacteroides fragilis modulates host immunity through polysaccharide A via the TLR2 pathway (38), whereas our results support that P. copri facilitates wound healing through a unique sphingolipid-mediated metabolic pathway.
Sphingosine-mediated mechanism and divergent microbial strategies
Our findings that P. copri-derived sphingosine accelerates wound healing present a fascinating parallel and point of divergence from the work of Zheng et al. (34), who demonstrated that commensal Staphylococcus epidermidis secretes a sphingomyelinase (ScpA) that hydrolyzes host-derived sphingomyelin into ceramides to fortify the epidermal permeability barrier in homeostatic skin. This comparison reveals that distinct commensals have evolved divergent strategies to modulate host lipid biology: S. epidermidis functions as an “enzymatic modifier” by processing host lipids on the skin surface, whereas P. copri acts as a “de novo synthesizer” of bacterial sphingosine within deep hypoxic niches like hair follicles and wound beds. Consequently, their physiological impacts are context-specific; while S. epidermidis primarily enhances structural barrier integrity, P. copri predominantly promotes dynamic tissue repair through stimulating keratinocyte migration and re-epithelialization.
The ceramide-microbiota axis across tissues
The intriguing interplay between microbiota and ceramide signaling is an emerging theme in host-microbe biology, particularly in the gut (39). Our discovery of a P. copri-ceramide axis in skin wound healing invites a comparative discussion of shared and unique features of this pathway across organ systems. Several shared features are notable: commensal bacteria serve as a significant source of sphingolipids in both gut and skin environments, with Bacteroidetes (e.g., Bacteroides spp.) being major producers in the gut (40), analogous to our findings involving the Bacteroidetes member P. copri on skin. Moreover, ceramides and their metabolites—such as sphingosine-1-phosphate (S1P)—from both sources are likely to engage conserved host signaling pathways (e.g., S1P receptors) to modulate fundamental cellular processes including epithelial proliferation and immune cell migration.
However, important differential features exist. First, the functional outcome is starkly different: bacterial ceramides in the gut are often associated with metabolic dysfunction and inflammation (39, 41), whereas *P. copri-*derived sphingosine demonstrates pro-healing and anti-inflammatory roles in acute skin wounds. Second, the microenvironments differ substantially; the gut constitutes a nutrient-rich, chronically antigenic setting where ceramide signaling frequently contributes to barrier dysfunction during dysbiosis, whereas the skin wound represents a site of active repair and transient inflammation in which localized ceramide/sphingosine delivery may preferentially promote re-epithelialization and inflammation resolution. Finally, the specific molecular species of bacterial sphingolipids (e.g., differing in chain length and saturation) may vary across bacterial genera and strains, potentially leading to divergent host responses—the unique structure of P. copri-derived sphingolipids could thus be a key determinant of its beneficial effects on skin.
Integrated healing mechanism and therapeutic implications
The bacterium’s therapeutic efficacy arises from a dual survival-repair strategy. Ecologically, β-lactamase genes create microbial sanctuaries, enabling persistence amidst antimicrobial challenges. Metabolically, P. copri synthesizes sphingosine, which activates the CerS1-ceramide axis to directly stimulate keratinocyte proliferation and neovascularization. This synergy of ecological resilience and metabolic signaling redefines P. copri: no longer just an opportunistic colonizer, it acts as a conductor of active repair, adept at dominating both disturbed and natural microenvironments by adapting its role to promote tissue regeneration.
Clinically, these insights pioneer two transformative approaches: ecological engineering to enhance P. copri colonization through competitive microbiome modulation and metabolite-based therapies utilizing sphingosine or stabilized ceramide derivatives. By bridging microbial ecology with precision metabolic intervention, this work shifts wound management from passive antimicrobial regimens to active microecosystem reprogramming, offering a blueprint for microbiome-guided regenerative medicine. The ability of a single commensal species to functionally replace complex community interactions challenges conventional paradigms, opening new frontiers in chronic wound therapy and host-microbe interface engineering.
Limitations and methodological considerations
The role of antibiotics in wound healing is complex and context-dependent. While some regimens, particularly targeted topical applications, can improve outcomes by reducing pathogen load or via direct anti-inflammatory effects (42), broad-spectrum systemic antibiotics often disrupt microbial ecology and impair healing. Our model, which examines recovery from such disruption, reveals that the specific compositional shifts post-antibiotics are critical: enrichment of a beneficial commensal like P. copri can rescue healing, whereas dominance of pathobionts would likely exacerbate delay.
Our reliance on bacterial conditioned media (CM), rather than viable P. copri, represents a deliberate strategy to isolate secreted bioactive factors; however, this approach also presents certain limitations. While effective for identifying soluble mediators such as sphingosine, the CM method may overlook contributions from non-secreted or cell-associated components—such as LPS—which could elicit divergent immune and metabolic responses. LPS derived from P. copri is of particular interest: although typically pro-inflammatory, its effects are highly context-dependent and may contradict the pro-regenerative outcomes observed in this study. Thus, the absence of whole-cell or specific component treatments precludes a comprehensive understanding of the full scope of P. copri-host interactions.
We cannot rule out the possibility that other species may also be involved and produce sphingolipid metabolites. In the future, we can use correlation analysis between species and metabolites to clarify the relationship and then verify it using germ-free mice.
Future directions
While our CD31 data demonstrate enhanced angiogenesis, a key component of wound healing, future studies will benefit from the inclusion of specific immunohistochemical markers such as keratin 14 to directly quantify re-epithelialization dynamics.
A more detailed kinetic analysis beyond the endpoint assessment would provide deeper insights into the healing dynamics. While the present study focused on the standard 7-day post-wounding time point—commonly used to evaluate mature granulation tissue and angiogenesis in murine models—future studies incorporating multi-time-point analyses will be essential to fully elucidate the temporal sequence and regulatory mechanisms underlying the pro-healing effects induced by P. copri metabolites.
The exclusive use of male mice in this study minimized the confounding effects of hormonal fluctuations associated with the estrous cycle (43); however, future investigations in female models are essential to confirm the generalizability and sexual dimorphism of the observed healing effects.
While our murine data establish a proof-of-concept mechanism, a critical future goal will be to test the effect of P. copri-derived sphingosine on wound healing in ex vivo human skin models and primary human keratinocytes to confirm its translational potential.
Future studies employing genetically engineered P. copri strains will be crucial to definitively establish the role of bacterial ceramide synthesis in mediating host-microbe interactions in vivo. To further validate the causal contribution of P. copri to wound healing, key genes involved in glycosphingolipid metabolism could be knocked out via genetic editing, followed by functional assessment of wound repair outcomes. Additionally, the development of bacteriophage-based targeting strategies against Prevotella could offer a selective microbial clearance tool to corroborate its therapeutic function. Complementing these approaches, the use of purified microbial molecules—such as LPS and outer membrane vesicles—will help dissect their individual contributions and clarify whether their effects align with or antagonize the pro-healing activities observed in this study.
MATERIALS AND METHODS
Establishment of a mouse skin injury model
Eight-week-old male C57BL/6 mice were anesthetized using avertin (250 mg/kg, intraperitoneal injection) and subjected to dorsal hair removal with electric clippers. Full-thickness excisional wounds (6 mm diameter) were created using a sterile biopsy punch (RWD Life Science) and surgical scissors to ensure complete removal of epidermal and dermal layers. Wound progression was documented daily under standardized imaging conditions with a fixed camera height and LED ring light. Mice were individually housed in ventilated cages and received daily topical treatments. Wound closure rates were quantified using ImageJ software by calculating the percentage reduction in wound area relative to the initial size. Only male mice were used in this study to minimize variability in wound healing rates and immune responses influenced by the fluctuating estrogen levels of the estrous cycle in females (43).
Preparation of ABX solution
Topical administration of the classical quadruple antibiotic regimen—comprising ampicillin (β-lactam), vancomycin (glycopeptide), metronidazole (nitroimidazole), and neomycin (aminoglycoside)—was employed to broadly deplete commensal microbiota. This combination synergistically targets diverse bacterial groups: ampicillin eliminates gram^+^/gram^−^ facultative anaerobes but inherently enriches β-lactamase-producing strains like Prevotella copri, which expresses chromosomally encoded cfxA penicillinases. Vancomycin suppresses gram^+^ competitors (e.g., Firmicutes), while neomycin depletes gram^−^ facultative anaerobes, indirectly promoting anaerobic expansion. Metronidazole exerts pressure on obligate anaerobes, though ~30% of P. copri strains evade this via nim gene-mediated resistance. Ampicillin (A5354), vancomycin (V0045000), metronidazole (M3761), and neomycin (33492) were procured from Sigma-Aldrich (St. Louis, MO, USA). The ABX solution was composed of 100 mg/kg ampicillin, 50 mg/kg vancomycin, 100 mg/kg metronidazole, and 100 mg/kg neomycin, administered twice daily by syringe drip.
ELISA detection of growth factors
Skin tissues from designated time points were homogenized in RIPA buffer containing protease inhibitors (Beyotime) and centrifuged to obtain supernatants. Growth factor concentrations (VEGF, EGF, and KGF) were measured using species-specific ELISA kits (Jiangsu Jingmei Biotech) according to manufacturer protocols. Absorbance at 450 nm was recorded with a microplate reader (SpectraMax iD5, Molecular Devices), and values were interpolated from standard curves.
Tissue processing and paraffin embedding
Excised wound tissues were fixed in 4% paraformaldehyde for 24 h at 4°C, followed by sequential dehydration in graded ethanol solutions (70%, 80%, 90%, 95%, 100%), clearing in xylene, and paraffin embedding using a histology embedding station (Leica EG1150H). Paraffin blocks were sectioned at 5 µm thickness with a rotary microtome (Leica RM2235), mounted on charged slides, and dried at 42°C for subsequent staining.
H&E staining
Deparaffinized sections were rehydrated through xylene and graded ethanol series. Nuclei were stained with Harris hematoxylin for 5 min, followed by cytoplasmic counterstaining with eosin Y for 1 min. Sections were dehydrated in ethanol, cleared in xylene, and coverslipped with resinous mounting medium. Tissue architecture was analyzed using bright-field microscopy.
Masson’s trichrome staining
Following deparaffinization and rehydration, sections were stained with Weigert’s hematoxylin for 10 min to visualize nuclei. Subsequent immersion in Ponceau S/acid fuchsin solution for 5 min highlighted connective tissue, followed by differentiation in 1% phosphomolybdic acid. Collagen fibers were selectively stained with aniline blue for 5 min, and sections were dehydrated, cleared, and mounted for analysis of matrix deposition patterns.
Immunohistochemical staining
Antigen retrieval was performed by heating sections in citrate buffer (pH 6.0) at 95°C for 20 min. Endogenous peroxidase activity was quenched with 3% hydrogen peroxide. After blocking with 3% BSA (Solarbio), sections were incubated overnight at 4°C with anti-CD31 primary antibody (1:200, Abcam), followed by HRP-conjugated secondary antibody (DAKO) for 30 min. DAB chromogen (DAKO) was applied for signal development, and nuclei were counterstained with Mayer’s hematoxylin. Sections were imaged using a high-resolution slide scanner (3DHISTECH PANNORAMIC 250) at 20× magnification.
Microbial sampling and 16S rRNA sequencing analysis of cutaneous wounds
To characterize the wound microbiota dynamics, sterile cotton swabs were rigorously rotated 60 times across wound beds at days 1 and 3 post-injury, ensuring comprehensive microbial collection. Swabs were immediately transferred to pre-chilled cryotubes, homogenized in sterile PBS with vortexing, and flash-frozen at −80°C until processing. Microbial genomic DNA was extracted using the MagPure Stool DNA KF Kit B (Magen Biotech), followed by PCR amplification of the 16S rRNA V3–-V4 hypervariable regions with universal primers 338F (5´- ACTCCTACGGGAGGCAGCAG-3´) and 806R (5´- GGACTACHVGGGTWTCTAAT-3´). Amplified libraries were purified using Agencourt AMPure XP beads (Beckman Coulter), quantified via Agilent 2100 Bioanalyzer (Agilent Technologies), and sequenced on the BGISEQ platform (BGI-Shenzhen).
Raw sequencing data underwent rigorous quality control: low-quality reads (Q-score <20), adapter contaminants, and ambiguous sequences (>5% N bases) were filtered using Readfq v.8. Paired-end reads were merged with FLASH (v.1.2.11) using a 10 bp minimum overlap. Chimeric sequences were identified and removed via UCHIME algorithm within USEARCH (v.11.0.667). Operational taxonomic units were clustered at 97% similarity using UPARSE, with taxonomic annotation performed against the Greengenes database (v.13_8) via the RDP classifier (v.2.13) with an 80% confidence threshold. Microbial community structure was visualized through principal coordinates analysis (ade4 package, R v.4.1.2) and hierarchical clustering heatmaps (ggplot2, R v.4.1.2). Species-level compositional profiles were generated to compare microbiota shifts between experimental groups. This integrated workflow ensures high-resolution characterization of wound microbiota ecology, linking taxonomic succession patterns with healing phenotypes through standardized bioinformatics pipelines.
Bacterial strain activation and culture techniques
Strain revival and subculturing
Lyophilized Prevotella copri stocks (BeNa Culture Collection, China) were rehydrated with 500 µL anaerobic sterile water within an anaerobic chamber (Coy Laboratory Products). Two hundred microliters of the suspension was spread onto pre-reduced Columbia blood agar plates (Hopebio) using sterile glass beads, followed by anaerobic incubation at 37°C for 5 days (AnaeroPack System, Mitsubishi Gas Chemical). Single colonies were isolated through quadrant streaking and subcultured under identical conditions to ensure purity.
Liquid culture protocol
A modified BHI broth was prepared containing 38.5 g/L brain heart infusion (Huankai Microbial, China), 1 g/L L-cysteine hydrochloride (MCE), 10 mg/L hemin chloride (MCE), and 1 mg/L vitamin K1 (MCE). The medium was adjusted to pH 6.9 ± 0.1, aliquoted into anaerobic tubes, and autoclaved at 121°C for 20 min. Primary plate cultures were inoculated into 10 mL broth and incubated statically at 37°C for 72 h under anaerobic conditions. Optical density at 600 nm (OD_600_) was monitored to confirm logarithmic growth phase (OD_600_ = 0.6–0.8).
Metabolomic profiling of bacterial secretions
Metabolites extraction
One hundred microliters of sample was taken, mixed with 400 µL of extraction solution (MeOH:ACN, 1:1 [vol/vol]), which contained deuterated internal standards. The mixed solution was vortexed for 30 s, sonicated for 10 min in a 4°C water bath, and incubated for 1 h at −40°C to precipitate proteins. Then the samples were centrifuged at 12,000 rpm (RCF = 13,800 (× g), R = 8.6 cm) for 15 min at 4°C. The supernatant was transferred to a fresh glass vial for analysis.
LC-MS/MS analysis
For polar metabolites, LC-MS/MS analyses were performed using an UHPLC system (Vanquish, Thermo Fisher Scientific) equipped with a Waters ACQUITY UPLC BEH Amide (2.1 mm × 50 mm, 1.7 µm) column coupled to Orbitrap Exploris 120 mass spectrometer (Thermo Fisher Scientific). The mobile phase consisted of 25 mmol/L ammonium acetate and 25 ammonia hydroxide in water (pH = 9.75) (A) and acetonitrile (B). The auto-sampler temperature was 4°C, and the injection volume was 2 µL. Mass spectrometry detection was operated in information-dependent acquisition (IDA) mode controlled by Xcalibur software (Thermo Fisher Scientific) to acquire MS/MS spectra. In this mode, the software continuously evaluates the full-scan MS spectrum. The ESI source conditions were set as follows: sheath gas flow rate 50 Arb, aux gas flow rate 15 Arb, capillary temperature 320°C, spray voltage 3.8 kV (positive) or −3.4 kV (negative), full MS resolution at 60,000, MS/MS resolution at 15,000. A stepped normalized collision energy (NCE) of 20, 30, and 40 was applied.
Data preprocessing and annotation
The raw data were converted to the mzXML format using ProteoWizard and processed with an in-house program, which was developed using R and based on XCMS, for peak detection, extraction, alignment, and integration. The R package and the BiotreeDB (v.3.0) were applied in metabolite identification.
Transcriptomic sequencing and bioinformatics
Total RNA was extracted from wound tissues using TRIzol (Invitrogen), with quality verified by Bioanalyzer 2100 (RIN > 8.0). Libraries were prepared using NEBNext Ultra II RNA Kit and sequenced on BGISEQ-500. Raw reads were quality-trimmed using Trimmomatic (SLIDINGWINDOW:4:20) and aligned to mm10 genome via STAR (v.2.7.9a). Gene expression quantification employed RSEM (v.1.3.3), with differential expression analysis using DESeq2 (|log2FC| > 1, Padj < 0.05). Functional enrichment of DEGs was performed through clusterProfiler (v.4.2.2) using GO databases, with hypergeometric testing and Benjamini-Hochberg correction (FDR < 0.05).
RNA isolation and quantitative PCR analysis
Total RNA was extracted from wound tissues using TRIzol (Invitrogen) following a standardized phenol-chloroform protocol. Briefly, homogenized tissues underwent phase separation with chloroform, and RNA was precipitated with isopropanol followed by 75% ethanol washes. RNA purity and concentration were quantified via NanoDrop 2000 spectrophotometry (A260/A280 > 1.8, A260/A230 > 2.0; Thermo Fisher Scientific). Reverse transcription of 1 µg total RNA was performed using the PrimeScript RT Reagent Kit (Takara Bio Inc., Japan) with oligo(dT) and random hexamer priming.
qPCR reactions were conducted in 20 µL volumes containing SYBR Green Premix Ex Taq II (Takara Bio) and gene-specific primers (sequences provided in Table S2) on a QuantStudio 7 Flex Real-Time PCR System (Thermo Fisher Scientific). Thermal cycling conditions included initial denaturation (95°C, 30 s), followed by 40 cycles of 95°C (5 s) and 60°C (30 s). Melt curve analysis (60–95°C, 0.3°C/s increments) confirmed primer specificity. Gene expression levels were normalized to 18S rRNA using the 2^−ΔΔCt^ method, with technical triplicates ensuring intra-assay precision.
Cell viability assay
Cells were seeded in 96-well plates at a density of 5 × 10^3^ cells/well and allowed to adhere for 12 h. After treatment with ceramide (0.781 µM) (Fig. S1), 10 µL CCK-8 reagent (Hanbio Biotechnology) was added to each well, and the plates were subsequently incubated for 3 h. The absorbance was measured at 450 nm with a reference wavelength of 650 nm using a microplate reader (SpectraMax iD5, Molecular Devices). Cell viability was calculated using the following formula: [(A_test_ – A_blank_) / (A_control_ – A_blank_)] × 100%, where A_blank_ represents the absorbance of the blank wells (containing culture medium and CCK-8 reagent only).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1James GA, Swogger E, Wolcott R, Pulcini ED, Secor P, Sestrich J, Costerton JW, Stewart PS. 2008. Biofilms in chronic wounds. Wound Repair Regen 16:37–44. doi:10.1111/j.1524-475X.2007.00321.x 18086294 · doi ↗ · pubmed ↗
- 2Malone M, Bjarnsholt T, Mc Bain AJ, James GA, Stoodley P, Leaper D, Tachi M, Schultz G, Swanson T, Wolcott RD. 2017. The prevalence of biofilms in chronic wounds: a systematic review and meta-analysis of published data. J Wound Care 26:20–25. doi:10.12968/jowc.2017.26.1.2028103163 · doi ↗ · pubmed ↗
- 3Kirketerp-Møller K, Jensen PØ, Fazli M, Madsen KG, Pedersen J, Moser C, Tolker-Nielsen T, Høiby N, Givskov M, Bjarnsholt T. 2008. Distribution, organization, and ecology of bacteria in chronic wounds. J Clin Microbiol 46:2717–2722. doi:10.1128/JCM.00501-0818508940 PMC 2519454 · doi ↗ · pubmed ↗
- 4Bjarnsholt T. 2013. The role of bacterial biofilms in chronic infections. APMIS 121:1–58. doi:10.1111/apm.1209923635385 · doi ↗ · pubmed ↗
- 5Grice EA, Segre JA. 2011. The skin microbiome. Nat Rev Microbiol 9:244–253. doi:10.1038/nrmicro 253721407241 PMC 3535073 · doi ↗ · pubmed ↗
- 6San Miguel AJ, Grice EA. 2015. Interactions between host factors and the skin microbiome. Cell Mol Life Sci 72:1499–1515. doi:10.1007/s 00018-014-1812-z 25548803 PMC 4376244 · doi ↗ · pubmed ↗
- 7Byrd AL, Deming C, Cassidy SKB, Harrison OJ, Ng W-I, Conlan S, NISC Comparative Sequencing Program, Belkaid Y, Segre JA, Kong HH. 2017. Staphylococcus aureus and Staphylococcus epidermidis strain diversity underlying pediatric atopic dermatitis. Sci Transl Med 9:eaal 4651. doi:10.1126/scitranslmed.aal 465128679656 PMC 5706545 · doi ↗ · pubmed ↗
- 8Smith SA, Travers RJ, Morrissey JH. 2015. How it all starts: initiation of the clotting cascade. Crit Rev Biochem Mol Biol 50:326–336. doi:10.3109/10409238.2015.105055026018600 PMC 4826570 · doi ↗ · pubmed ↗