An integrated phenotypic and genomic approach to characterize MBL-producing Enterobacterales strains circulating in a Sicilian transplant center
Claudia Vaiana, Carol Lee, Roberta Vazzana, Andrea Cona, Giovanni Mulè, Caterina Amato, Francesco Monaco, Daniele Di Carlo, Pier Giulio Conaldi, Valentina Agnese, Nicola Cuscino, Alessandra Mularoni, Alessia Gallo

TL;DR
This study combines phenotypic and genomic methods to analyze MBL-producing Enterobacterales strains in a Sicilian hospital, revealing high resistance and the need for surveillance.
Contribution
The paper introduces an integrated approach combining phenotypic and genomic data to characterize MBL-producing strains in a clinical setting.
Findings
New Delhi MBL was the most common (82%), followed by VIM (18%) in MBL-producing strains.
High resistance to amikacin (63%) and reduced susceptibility to cefiderocol (50%) were observed.
High-risk clones like K. pneumoniae ST147 and E. coli ST648 were identified, emphasizing the need for surveillance.
Abstract
Metallo-β-lactamases (MBLs)-producing Enterobacterales infections represent a serious threat in clinical practice due to high mortality rates associated and the limited therapeutic options available. Given their increased dissemination across Europe and their involvement in hospital outbreaks, further epidemiological investigations are required. Here, we provide a comprehensive characterization, by integrating clinical, genomic, and phenotypic data, of 22 MBL-carrying Enterobacterales strains (17 Klebsiella pneumoniae and 5 Escherichia coli), isolated from critically ill patients admitted to IRCCS ISMETT from 2021 to 2024. Bacterial antimicrobial susceptibility was evaluated using phenotypic methods. Using a whole-genome sequencing approach, we identified the sequence types (STs) and depicted the resistome and virulome content. This study revealed the circulation of alarming…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1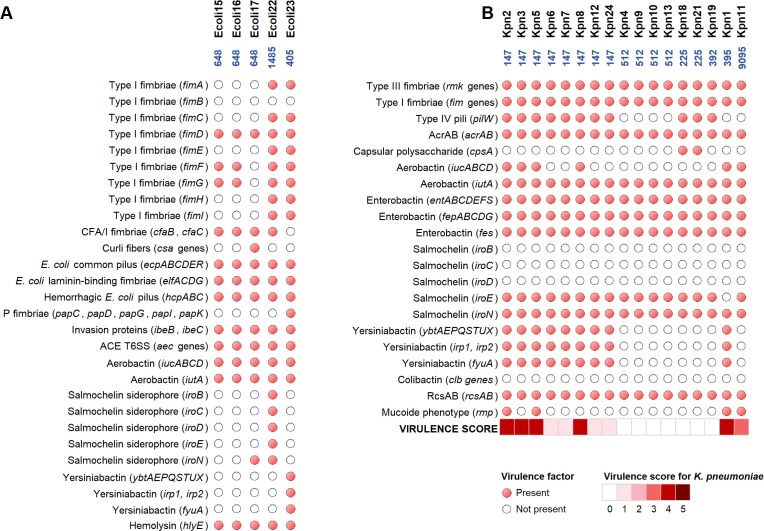 Fig 2
Fig 2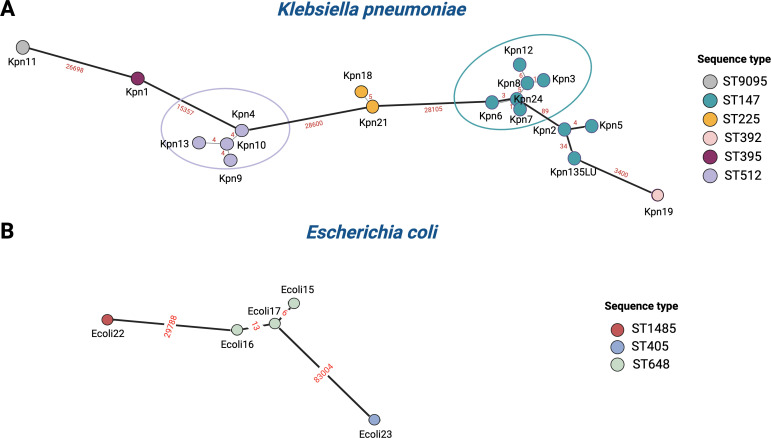 Fig 3
Fig 3| Patients’ characteristic | Infection ( |
|---|---|
| Age | 55 (2–80 years) |
| Male sex | 12 (54.5%) |
| ICU stay (>2 days) | 18 (81.8%) |
| Comorbid conditions | |
| Cardiovascular disease | 12 (54.5%) |
| Diabetes mellitus | 3 (13.6%) |
| Respiratory tract disease | 2 (9%) |
| Chronic kidney disease | 5 (22.7%) |
| Chronic liver disease | 6 (27.7%) |
| Malignancy | 4 (18.1%) |
| Organ solid transplant | 11 (50%) |
| Liver | 4 (36.3%) |
| Kidney | 2 (18.1%) |
| Lung | 2 (18.1%) |
| Heart | 2 (18.1%) |
| Kidney and liver | 1 (9%) |
| RT or CT last 90 days | 1 (4.5%) |
| Immunosuppression | 11 (50%) |
| Previous hospitalization in the last 3 months | 17 (77.2%) |
| Transfer patient (>2 days institutionalized in another facility) | 11 (50%) |
| Use of antibiotic in the last 90 days | 16 (72.7%) |
| Carbapenem | 4 (25%) |
| Third-generation cephalosporin | 3 (18.7%) |
| BL/BLI combinations | 11 (68.7%) |
| Colistin | 3 (18.7%) |
| Quinolones | 2 (12.5%) |
| Cefiderocol | 2 (12.5%) |
| LOS | 48 days (5–232 days) |
| Invasive devices | 19 (76.4%) |
| Abdominal surgery | 6 (31.5%) |
| Cardiac surgery | 4 (21%) |
| Thoracic surgery | 2 (10.5%) |
| Endoscopic procedure | 7 (36.8%) |
| Use of invasive diapositives (>2 days) | 20 (90.9%) |
| Co-colonization with KPC enzyme | 15 (68.1%) |
| Co-producer of KPC and MBL | 5 (33.3%) |
| Year | Sex | Age | Length of stay | Isolate | Species | Carbapenemases | Source of infection | Treatment | Subsequent infection/Site | 30-day mortality |
|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | M | 74 | 49 | Ecoli15 |
| NDM-5 | Abdominal | AMK, TGC, FOS | Yes/Abdominal | Alive |
| 2021 | F | 67 | 18 | Ecoli16 |
| NDM-5 | Abdominal | AMK, TGC, FOS | No | Alive |
| 2021 | M | 59 | 99 | Ecoli17 |
| NDM-5 | Blood | CZA + ATM, AMK | Yes/Abdominal | Alive |
| 2022 | F | 2 | 5 | Ecoli22 |
| VIM-1 | Abdominal | CZA + ATM, CST | No | Alive |
| 2022 | M | 62 | 232 | Kpn19 |
| VIM-1 | Blood | CZA + ATM | No | Deceased |
| 2022 | F | 70 | 106 | Kpn21 |
| VIM-1 | Blood | CZA + ATM | No | Deceased |
| 2022 | M | 46 | 47 | Kpn18 |
| VIM-1 | Blood | FDC | No | Transferred |
| 2023 | M | 48 | 87 | Kpn6 |
| NDM-1 | Abdominal | CZA + ATM | No | Deceased |
| 2023 | M | 54 | 25 | Ecoli23 |
| NDM-5 | Blood | CZA + ATM | No | Deceased of infection |
| 2023 | M | 69 | 123 | Kpn7 |
| NDM-1 | Respiratory | CST | No | Transferred |
| 2023 | F | 38 | 27 | Kpn24 |
| NDM-1 | Skin and soft tissue | CZA + ATM | No | Alive |
| 2023 | F | 67 | 171 | Kpn8 |
| NDM-1 | Urinary | FOS | Yes/Respiratory | Deceased of infection |
| 2024 | F | 63 | 8 | Kpn1 |
| NDM-1 | Abdominal | CZA + ATM | No | Alive |
| 2024 | M | 66 | 35 | Kpn3 |
| NDM-1 | Blood | FDC | Yes/Urinary | Alive |
| 2024 | M | 51 | 7 | Kpn5 |
| NDM-1 | Blood | CZA + ATM | Yes/Abdominal | Alive |
| 2024 | F | 80 | 83 | Kpn4 |
| NDM-1, KPC-3 | Blood | CZA + ATM | No | Deceased |
| 2024 | M | 62 | 131 | Kpn2 |
| NDM-1 | Blood | FDC | Yes/Abdominal | Deceased of infection |
| 2024 | M | 61 | 31 | Kpn9 |
| NDM-1 | Blood | CST, CZA | No | Transferred |
| 2024 | F | 21 | 28 | Kpn13 |
| NDM-1, KPC-3 | Respiratory | FDC | No | Transferred |
| 2024 | M | 61 | 142 | Kpn10 |
| NDM-1, KPC-3 | Skin and soft tissue | CZA + ATM, AMK | No | Alive |
| 2024 | F | 62 | 36 | Kpn12 |
| NDM-1 | Skin and soft tissue | N/A | No | Transferred |
| 2024 | M | 45 | 55 | Kpn11 |
| NDM-1, KPC-3 | Urinary | CZA + ATM | No | Alive |
| Isolate | Collection date (day/mo/yr) | Specimen source | Carbapenemases | ST | Predicted serotype | MIC (µg/mL) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMK | FEP | CTX | CAZ | CZA | C/T | CIP | CST | GEN | IMR | MEM | MEV | TZP | TMP/SMX | TOB | ATM | TGC | FDC | AZA | ||||||
| Ecoli15 | 16/09/2021 | Biliary liquid | NDM-5 | 648 | H4-O8 | 4 (S) | 8 (R) | 32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | ≤20 (S) | >8 (R) | >32 (R) | ≤0.5 | 2 (S) | 1.5 (S) |
| Ecoli16 | 20/09/2021 | Rectal swab | NDM-5 | 648 | H4-O8 | 4 (S) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | ≤20 (S) | >8 (R) | >32 (R) | ≤0.5 | 2 (S) | 1.5 (S) |
| Ecoli17 | 12/10/2021 | Rectal swab | NDM-5 | 648 | H4-O8 | 8 (S) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | ≤20 (S) | >8 (R) | >32 (R) | ≤0.5 | 2 (S) | 1.5 (S) |
| Ecoli22 | 10/10/2022 | Rectal swab | VIM-1 | 1485 | H42-O83 | 4 (S) | 16 (R) | 32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | ≤0.5 (S) | ≤1 (S) | 4 (R) | 2 (I) | 2 (S) | >64 (R) | >160 (R) | 8 (R) | ≤1 (S) | ≤0.5 | 0.25 (S) | 0.19 (S) |
| Ecoli23 | 30/04/2023 | Rectal swab | NDM-5 | 405 | H6-O102 | >32 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | >160 (R) | >8 (R) | >32 (R) | ≤0.5 | 2 (S) | 3 (S) |
| Kpn18 | 23/01/2022 | Rectal swab | VIM-1 | 225 | K3-O2afg | ≤1 (S) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | >8 (R) | ND | 4 (R) | >8 (R) | 32 (R) | >64 (R) | >160 (R) | 8 (R) | >32 (R) | >4 | 1 (S) | 0.25 (S) |
| Kpn19 | 17/03/2022 | Blood | VIM-1 | 392 | K27-O4 | 8 (I) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | >8 (R) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | >160 (R) | >8 (R) | >32 (R) | 1 | 0.5 (S) | 0.25 (S) |
| Kpn21 | 23/04/2022 | Rectal swab | VIM-1 | 225 | K3-O2afg | ≤1 (S) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | >8 (R) | 2 (S) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | >160 (R) | 8 (R) | >32 (R) | >4 | 1 (S) | 0.25 (S) |
| Kpn24 | 05/08/2023 | Rectal swab | NDM-1 | 147 | K64-O2a | 32 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | ≤1 (S) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | 80 (I) | >8 (R) | >32 (R) | 4 | 0.5 (S) | 0.125 (S) |
| Kpn6 | 18/11/2023 | JP fluid | NDM-1 | 147 | K64-O2a | 16 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | ≤1 (S) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | >160 (R) | ND | >32 (R) | 4 | 1 (S) | 0.25 (S) |
| Kpn7 | 26/11/2023 | Tracheal aspirate | NDM-1 | 147 | K64-O2a | 16 (R) | >16 (R) | ND | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | ≤1 (S) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | ND | ND | >32 (R) | ND | 0.25 (S) | 0.094 (S) |
| Kpn8 | 12/12/2023 | Urinary tract | NDM-1 | 147 | K64-O2a | 16 (R) | >16 (R) | ND | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | ND | ND | >32 (R) | ND | 2 (S) | 0.38 (S) |
| Kpn9 | 05/01/2024 | Blood | NDM-1 | 512 | unk-O2afg | 32 (R) | >16 (R) | ND | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | ND | ND | >32 (R) | ND | 2 (S) | 0.5 (S) |
| Kpn10 | 10/01/2024 | Wound swab | NDM-1, KPC-3 | 512 | unk-O2afg | 32 (R) | >16 (R) | ND | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | ND | ND | >32 (R) | ND | 2 (S) | 0.75 (S) |
| Kpn11 | 11/01/2024 | Urinary tract | NDM-1, KPC-3 | 9095 | unk-O2afg | 32 (R) | >16 (R) | ND | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 2 (S) | >8 (R) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | ND | ND | >32 (R) | ND | 1 (S) | 0.25 (S) |
| Kpn12 | 17/01/2024 | Rectal swab | NDM-1 | 147 | K64-O2a | 16 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | ≤1 (S) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | 160 (R) | >8 (R) | >32 (R) | 4 | 2 (S) | 0.25 (S) |
| Kpn13 | 18/01/2024 | Rectal swab | NDM-1, KPC-3 | 512 | unk-O2afg | 32 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | >160 (R) | >8 (R) | >32 (R) | 2 | 2 (S) | 0.5 (S) |
| Kpn3 | 10/04/2024 | Rectal swab | NDM-1 | 147 | K64-O2a | 16 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | >32 (R) | >64 (R) | >160 (R) | >8 (R) | >32 (R) | 4 | 1 (S) | 0.38 (S) |
| Kpn1 | 12/04/2024 | Rectal swab | NDM-1 | 395 | K2-O1ab | 32 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | >160 (R) | >8 (R) | >32 (R) | 1 | 4 (R) | 0.125 (S) |
| Kpn2 | 12/04/2024 | Blood | NDM-1 | 147 | K64-O2a | ND | >16 (R) | ND | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | ≤1 (S) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | ND | ND | >32 (R) | ND | 0.5 (S) | 0.19 (S) |
| Kpn5 | 19/04/2024 | Rectal swab | NDM-1 | 147 | K64-O2a | 16 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | ≤1 (S) | >8 (R) | 8 (I) | 32 (R) | >64 (R) | >160 (R) | 8 (R) | >32 (R) | ND | 1 (S) | 0.125 (S) |
| Kpn4 | 09/05/2024 | Rectal swab | NDM-1, KPC-3 | 512 | unk-O2afg | 32 (R) | >16 (R) | >32 (R) | >32 (R) | >8 (R) | >16 (R) | >2 (R) | 0.5 (S) | >8 (R) | >8 (R) | >8 (R) | 32 (R) | >64 (R) | >160 (R) | >8 (R) | >32 (R) | 4 | 2 (S) | 0.38 (S) |
- —Ministero della Salutehttp://dx.doi.org/10.13039/501100003196
- —Ministero dell'Università e della Ricercahttp://dx.doi.org/10.13039/501100021856
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotics Pharmacokinetics and Efficacy · Antibiotic Use and Resistance
INTRODUCTION
Carbapenem-resistant Enterobacterales (CRE) constitute a major global health concern due to their rapid dissemination in healthcare settings and their ability to cause severe infections, which are associated with substantial morbidity and mortality (1). The primary mechanism driving carbapenem resistance in CRE is the production of β-lactamases (2). Among these, metallo-β-lactamases (MBLs) are of particular concern, with New Delhi MBL (NDM), Verona integron-encoded MBL (VIM), and imipenemase (IMP) being their most epidemiologically important members (3). MBLs are endemic in India and Southeast Asia, with the largest burden of cases (4). However, in recent years, clinical events involving these enzymes have been increasingly reported worldwide, especially in Europe (4, 5). This growing prevalence is alarming due to their ability to hydrolyze nearly all β-lactams (BLs), including penicillins, cephalosporins, carbapenems, and novel BL/β-lactamase inhibitor (BLI) combinations, and their frequent association with other resistance mechanisms, including co-production of other β-lactamases, resulting in a highly multidrug-resistant (MDR) phenotype (6, 7). In Italy, VIM has historically been the most prevalent MBL among Enterobacterales and also the predominant carbapenemase in Pseudomonas aeruginosa (4). However, a significant outbreak of NDM-producing Klebsiella pneumoniae ST147 in Tuscany may signal a shift in this distribution (8). Since this event, an increasing number of NDM-producing isolates have been reported across other Italian regions (9, 10), raising concerns about the further spread of this resistance mechanism. Compounding this issue, some strains showed reduced susceptibility to cefiderocol, a critical last-line agent for these infections (11). The acquisition of MBLs is strongly associated with factors such as prior antimicrobial exposure, intensive care unit (ICU) admission, prolonged hospitalization, comorbidities, and invasive devices (12). Importantly, previous colonization significantly increases the risk of subsequent infection (13), as demonstrated by the higher probability of bloodstream infection development in rectal carriers of NDM-producing K. pneumoniae than in those colonized with KPC-producing strains (14), an event most likely linked to the specific clonal lineage rather than the carbapenemase type. These findings underscore the importance of active surveillance and infection control measures to limit intra-hospital horizontal transmission (15). Mortality associated with MBL-producing Enterobacterales infections is high, ranging from 21% to 44%, and up to 55% attributable (16). Emerging data indicate a strong correlation between the selected antibiotic therapy and patient outcomes. The combination of ceftazidime-avibactam and aztreonam is currently regarded as the preferred treatment for MBL-producing Enterobacterales, showing superior efficacy compared to alternative regimens (17). The expanding distribution of MBL-harboring Enterobacterales and the limited treatment arsenal highlight the urgent need for robust infection control and optimized therapeutic strategies (18). In this context, our study aimed to comprehensively analyze MBL-producing Enterobacterales isolated from critically ill patients. We emphasized the genomic characterization of these strains as a pivotal tool to (i) identify the genetic determinants of antimicrobial resistance and virulence, (ii) define circulating sequence types (STs), and (iii) assess clonal relationships and identify potential transmission clusters. To the best of our knowledge, this is the first study that integrates patients’ clinical data with genomic profiling of MBL-producing organisms using this approach.
MATERIALS AND METHODS
Patient data collection and bacterial strains isolation
During the period from January 2021 to September 2024, 127 patients admitted at IRCCS ISMETT, a solid organ transplant center, located in Palermo (Italy), were found to have positive cultures for MBL-producing Enterobacterales. Among them, 101/127 patients were colonized without signs of infection, while 26/127 patients developed infection. Of these, only four patients were excluded from this study because clinical data or bacterial isolates were missing. Patients were followed up until death or 30 days from infection. The last follow-up date was 20 April 2025, used as the censoring date for patients still alive.
We retrospectively collected patient demographic and clinical data, including age, sex, admission and discharge dates, underlying conditions, antibiotic treatments, and clinical outcomes from the hospital electronic records. Five of these isolates were collected from clinically relevant samples, three in blood, one in bile fluid, and one in the respiratory tract, and were directly linked to infectious episodes. The remaining 17 isolates, initially identified during colonization, were linked to subsequent infections using the concordance criteria, established by Falcone et al. (14). Based on these criteria, we confidently assumed that these colonization-related isolates, showing identical resistance markers and antimicrobial profiles, were responsible for the subsequent infectious events. Bacterial identification and antimicrobial susceptibility tests were performed by the hospital diagnostic laboratory as part of the routine management of the patients. Subsequently, these isolates were cryopreserved at −80°C in Luria-Bertani broth with 20% glycerol for future analyses, including additional antibiotic susceptibility testing and genomic investigations.
Bacterial identification, antimicrobial susceptibility testing, and MBL detection
Bacterial identification was performed by matrix-assisted laser desorption ionization–time-of-flight mass spectrometry Vitek MS system (bioMérieux, Marcy-l'Étoile, France). Antimicrobial susceptibility to most of the antibiotics of interest was investigated using the VITEK 2 system (bioMérieux, Marcy-l'Étoile, France), while resistance to colistin was confirmed using Sensititre (Thermo Fisher Scientific, MA, USA). Susceptibility to cefiderocol was tested with the ComASP cefiderocol microdilution panel (0.008 µg/mL–128 µg/mL) (Liofilchem, Roseto degli Abruzzi, Italy). Minimum inhibitory concentration (MIC) results were interpreted according to the EUCAST clinical breakpoints (EUCAST clinical breakpoint guidance v.15.0). Susceptibility to aztreonam-avibactam was tested using the ETEST Aztreonam/Avibactam (AZA) (0.016/4 µg/mL–256/4 µg/mL) (bioMérieux, Marcy-l'Étoile, France). Production of class A and B carbapenemases was verified using carbapenemase inhibition tests with boronic acid derivates and dipicolinic acid/EDTA (Rosco Diagnostica A/S, Taastrup, Denmark). Identification of the five major carbapenemases (KPC, NDM, VIM, IMP, and OXA-48-like) from bacterial isolates was performed using immunochromatographic tests, including NG-Test Carba 5 (NG Biotech, Guipry, France) or the lateral flow immunoassay (Medomics), according to the manufacturer’s instructions.
DNA extraction and whole-genome sequencing (WGS)
Pure bacterial colonies were grown on MacConkey agar plates, and then genomic DNA was extracted using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany), by following the manufacturer’s instructions. DNA was subsequently quantified using the Qubit 2.0 fluorometer (Invitrogen, Carlsbad, CA, USA) with the Qubit dsDNA Broad Range Assay Kit. DNA library preparation was performed using the Illumina DNA Prep Kit (Illumina, San Diego, CA, USA) according to the manufacturer’s instructions. Nextera DNA CD Indexes (96 Indexes–96 Samples; Illumina, San Diego, CA, USA) were ligated after amplification. DNA libraries were evaluated on the Agilent 4200 TapeStation System (Agilent Technologies Ltd., USA), while library concentration was checked using the Qubit 2.0 fluorometer (Invitrogen, MA, USA). Whole-genome sequencing was performed according to the manufacturer’s instructions on the NextSeq 550 System using a 2 × 150 bp paired-end library (Illumina Inc., San Diego, CA, USA).
Bioinformatic analysis
The draft genome and plasmids were assembled using SPAdes v.3.15.12 software (https://github.com/ablab/spades) (19), and automatic annotation was performed with Prokka v.1.14.5 (https://github.com/tseemann/prokka) (20). The MLST STs were identified on the MLST database hosted by the CGE (https://cge.food.dtu.dk/services/MLST/) (21). Subsequently, the Kaptive Web algorithm (22) and SerotypeFinder 2.0 (23) were employed for in silico serotyping of K. pneumoniae and Escherichia coli isolates, respectively. Antimicrobial resistance genes (ARGs) and virulence factors were detected using CARD (24) and VFDB (25), respectively. K. pneumoniae strain cohort was additionally assessed for their virulence score using Kleborate (26). Plasmid incompatibility types were identified using PlasmidFinder on the CGE (https://cge.food.dtu.dk/services/PlasmidFinder/) website (27). Contigs containing MBL genes were initially aligned to plasmids identified using PlasmidFinder with the BLASTn algorithm (BLAST+ v.2.16.0;) (28). Contigs showing less than 80% identity and coverage with the reference plasmids were subsequently re-aligned against the NCBI nt database for further characterization. Kpn1 and Ecoli23 isolates were used as reference genomes for the core-genome SNP (cgSNP) analysis. The number of SNPs in the core genome was calculated from the de novo assembled genomes for isolates with the same ST (ST147, 512, 648, 225) using the Harvest software suite (Parsnp), including a cgSNP-based phylogenetic tree. A minimum spanning tree was constructed using R software (v.4.2.2) (https://www.R-project.org/).
RESULTS
Clinical characteristics and cases’ description
During the study period, a total of 22 patients with infection due to MBL-producing organisms were included. The median age of the study population was 61.5 years (IQR, 48.8–66.8 years), and the majority were male (54.5%). Pre-existing comorbidities were frequent, with cardiovascular disease (54.5%), chronic liver disease (27.7%), and chronic kidney disease (22.7%) being the most prevalent. Patients’ characteristics of the study cohort are detailed in Table 1. Half of the patients (50%) were solid organ transplant recipients: liver (36.3%) and kidney (18.1%) transplants being the most common. Seventeen patients (77.2%) had been hospitalized for more than 2 days in the previous 3 months, and half of the included patients had been transferred from another healthcare facility. Prior antibiotic use within 90 days before isolation of the MBL-producing organism, whether from surveillance or clinical specimens, was reported in 72.7% (16/22) of patients. Among those, BL/BLI combinations were the most frequently used agents (68.7%), followed by carbapenems (25%), third-generation cephalosporins and colistin (each 18.7%), quinolones, and cefiderocol (each 12.5%). The median length of hospital stay (LOS) was 48 days (IQR, 27–104 days). Co-colonization with KPC-producing organisms was observed in 68.1% (15/22) of patients, with 33.3% (5/15) of these harboring isolates co-producing both KPC and MBL enzymes.
The most common sources of infection were bloodstream (45.4%) and intra-abdominal site (22.7%). Clinical infection characteristics and treatment data are summarized in Table 2. Among the 10 bloodstream infections, only two were caused by E. coli, harboring NDM-5, and the remaining eight were caused by K. pneumoniae. Interestingly, 27.2% of patients (6/22) developed a subsequent infection at a different anatomical site during follow-up; notably, only one case involved the same anatomical site as the initial infection. The primary treatment regimen across the cohort was the combination of ceftazidime-avibactam and aztreonam, used in 54.4% (12/22) of cases, followed by cefiderocol in 18.1% (4/22) and colistin in 13.6% (3/22). Among patients with bloodstream infections, ceftazidime-avibactam plus aztreonam was the main regimen in 60% (6/10), while cefiderocol was used in 30% of cases (3/10). For intra-abdominal infections, a combination of tigecycline and an aminoglycoside played a role in 40% (2/5) of the cases. Among patients analyzed, the 30-day mortality rate was 31.8% (7/22). At the last follow-up in March 2025, 12 patients (54.5%) were alive, 7 (31.8%) had died, and for 3 (13.6%), follow-up information was not available. Of the seven deceased patients, five died during hospitalization, with MBL infection identified as the primary cause in three cases. The remaining two deaths occurred during the study follow-up and were attributed to causes unrelated to the MBL infection.
Identification, antimicrobial susceptibility profile, and MBL detection
The study population included 17 K. pneumoniae and 5 E. coli isolates. The most prevalent MBL was NDM, detected in 82% of the isolates (18/22), and followed by VIM, present in 18% of the isolates (4/22). Among the NDM-positive isolates, 14 out of 18 carried the NDM-1 variant, while 4 out of 18 had NDM-5, which was exclusively found in E. coli. In contrast, just one VIM variant, VIM-1, was detected.
All isolates were examined for susceptibility to multiple antimicrobial agents, with particular focus on those frequently used for treatment of MBL-specific infections. As depicted in Table 3, consistent resistance was noted, as expected, across all isolates to fluoroquinolones (ciprofloxacin), last-generation cephalosporins (cefepime, ceftriaxone, and ceftazidime), and BL/BLI combinations (ceftolozane-tazobactam, ceftazidime-avibactam, and imipenem-relebactam), while susceptibility to meropenem-vaborbactam was observed only in 1 out of the 22 isolates. No isolate was susceptible to meropenem. Thus, isolates showed a typical antimicrobial resistance profile of MBL-producing bacteria. Nevertheless, almost all isolates (21/22) were also resistant to aztreonam, likely due to the presence of other β-lactamases not detected with phenotypic tests. Regarding aminoglycosides, all tested isolates were resistant to tobramycin, while 63.6% (14 out of 22) and 59% (13 out of 22) of isolates were resistant also to amikacin and gentamicin, respectively. The highest susceptibility rates were observed for colistin and cefiderocol, with 19/22 (86%) and 21/22 (95%) susceptible isolates, respectively. Interestingly, all isolates were susceptible to aztreonam-avibactam combination (Table 3).
Some notable differences in antibiotic susceptibility were observed both between K. pneumoniae and E. coli isolates and within isolates of the same species. All E. coli isolates harboring either NDM-5 or VIM-1 exhibited susceptibility to colistin, tigecycline, and cefiderocol. Compared to other STs, the ST1485 isolate was unique in its susceptibility to gentamicin, aztreonam, and meropenem-vaborbactam. Unlike ST648 isolates, which were susceptible to trimethoprim-sulfamethoxazole, ST405 and ST1485 isolates displayed a high level of resistance to this antibiotic combination. Similar to E. coli, K. pneumoniae strains carrying an MBL, alone or with KPC-3, demonstrated broad resistance to β-lactams and fluoroquinolones. Of note, all isolates resistant to colistin were VIM-carrying K. pneumoniae, belonging to ST225 and ST392, while NDM-1-positive Kpn11 showed reduced susceptibility (MIC = 2 mg/L). Although cefiderocol susceptibility was widely reported, nearly half of isolates exhibited an MIC value of 2 mg/L, which is extremely close to the EUCAST breakpoint (resistant phenotype with MIC >2 mg/L), indicating a significantly reduced susceptibility margin.
Genomic analysis: sequence type, resistance, and virulence
Based on MLST analysis, a heterogeneous ST distribution was observed. In particular, among the 17 K. pneumoniae strains, ST147 was the most prevalent ST detected in our investigation (eight isolates), followed by ST512 (four isolates), ST225 (two isolates), ST392 (one isolate), and ST395 (one isolate). Furthermore, one isolate belonged to an ST not previously identified. Among the five E. coli isolates, three isolates were assigned to ST648, while the remaining two isolates belonged to ST1485 and ST405, respectively. Furthermore, plasmid content was investigated in each isolate. Among K. pneumoniae isolates, the most prevalent plasmid Inc type is IncFIB. In E. coli isolates, the most frequently observed Inc types appear to be IncFIB(AP001918) and IncFII(pRSB107), often co-occurring with other plasmids like Col-types (File S1).
Genomic characterization of the isolates revealed a complex resistance landscape, characterized by the occurrence of porins, efflux pumps, and enzyme variants responsible for their altered functional profiles (File S2). The ARGs responsible for the observed resistance to the antibiotics tested in this study are summarized in Fig. 1.
ARG distribution among the isolates analyzed. Each column represents a single bacterial isolate, with its ST indicated at the top, and each row corresponds to a specific resistance gene. A colored cell indicates the presence of that particular gene in the corresponding isolate. The identified ARGs have been classified based on their respective gene family.
WGS showed that genes encoding various β-lactamases appear to be highly prevalent and that all isolates carried MBL genes located on plasmids (File S3). Among MBLs, NDM-1 was detected in all NDM-producing K. pneumoniae, while NDM-5 was limited to E. coli ST648 and ST405. In contrast, VIM-1 was found both in E. coli (one ST1485) and K. pneumoniae (two ST225, one ST392) isolates. Of note, five K. pneumoniae isolates, belonging to ST512 and ST9095, were characterized by the concomitant presence of NDM-1 and KPC-3. Additionally, we identified other β-lactamases, with the most common being OXA (found in 18/22 isolates), followed by SHV (17/22), TEM (18/22), and CTX-M-15 (14/22). Carbapenemase-encoding blaKPC and β-lactamase-encoding blaEC were less common. A notable finding was the presence of a remarkably high number of β-lactamase-encoding genes in 11 K. pneumoniae strains, ranging from five to six per isolate (Fig. 1). Moreover, the co-occurrence of distinct variants of the same β-lactamase (OXA-1 and OXA-9) was also observed. Genes associated with aminoglycoside resistance were found. Specifically, genes encoding aminoglycoside-modifying enzymes (AAC3′, AAC6′, ANT(2), ANT(3′′), APH(3), APH(6)) showed varying and wide distribution among the isolates. Instead, armA and rmtB appeared to be less common. Among the determinants related to sulfamethoxazole and trimethoprim resistance, sul genes (sul1, sul2) and dfrA genes (dfrA1, dfrA5, dfrA12, dfrA14) were identified in 19/22 isolates and 13/22 isolates, respectively. Additionally, most isolates exhibited genes related to quinolone resistance, qnrS1, either alone or in combination with qnrB1. Genes related to polymyxin resistance, including pmrF and arnT, were detected in all isolates. An ept gene (eptA or eptB), implicated in colistin resistance, was widely distributed, while the ugd gene was present in two isolates. Finally, other genes like catA, pmrA, and pmrB exhibited limited distribution in the analyzed isolates. Focusing on the most prevalent STs for each species, we discovered that K. pneumoniae ST147 exhibited a wide range of resistance determinants associated with resistance to β-lactams (SHV variants, TEM-1, CTX-M-15, OXA variants), aminoglycosides, fluoroquinolones, and sulfonamides-trimethoprim. E. coli ST648 carried multiple genes conferring resistance to aminoglycosides and fluoroquinolones, in addition to NDM-5 and CTX-M-15.
Virulome analysis demonstrated a heterogeneous distribution of virulence factor-encoding genes across bacterial species and sequence types, as illustrated in Fig. 2.
Virulence factor-encoding gene distribution among the E. coli (A) and K. pneumoniae isolates (B). Red circles indicate the presence of the gene. Virulence score (0–5) was assigned to K. pneumoniae strains according to Kleborate analysis.
Among them, iron uptake systems are of great concern. Particularly, genes for enterobactin (entABCDEFS, fepABCDG, fes) were widely distributed in all K. pneumoniae isolates, while aerobactin-encoding genes have a variable distribution, with iucABCD operon found in all E. coli isolates and in 6 out of 17 K. pneumoniae isolates, and iutA was present in all the analyzed isolates. Moreover, all K. pneumoniae isolates and two E. coli isolates (Ecoli17 and Ecoli22) carried at least one gene from the salmochelin-encoding iroBCDEN operon. Furthermore, yersiniabactin-encoding genes (ybtAEPQSTUX, irp1, irp2, fyuA) were detected in K. pneumoniae ST147 and ST395 isolates, and in E. coli ST405. All K. pneumoniae isolates carried acrAB, fim, and rmk genes, encoding the AcrAB efflux pump, type I and type III fimbriae, respectively. Additionally, ST147 and ST225 isolates also harbored genes for type IV pili (pilW) and capsule polysaccharide (cpsA). According to Kleborate analysis, virulence scores, ranging from 0 to 5, were assigned to assess virulome content. Specifically, the highest virulence score was exhibited by five ST147 and one ST395, while ST9095 and four ST147 were assigned medium and low virulence scores, respectively. Finally, genes associated with capsule polysaccharide synthesis and hypermucoidy (rmpA, rmpA2, and rmpC) were detected in Kpn1, Kpn2, Kpn5, and Kpn11 strains. In contrast, all E. coli isolates harbored additional virulence factors, including genes for type I fimbriae, E. coli common pilus (ECP), hemorrhagic E. coli common pilus (HCP), hemolysin (hlyE), and invasion proteins (ibeB, ibeC, and tia).
Core-genome SNP analysis
The genetic relationships among the isolates were investigated in detail using core-genome SNP analysis, as shown in Fig. 3. As expected, isolates belonging to different STs generally showed greater genetic diversity. Among K. pneumoniae strains, all ST512 isolates demonstrated high genetic similarity, with only four SNPs separating them, suggesting a tightly linked cluster. An additional cluster comprising six isolates, with a median SNP difference of 3, was revealed within the ST147 group. In contrast, Kpn5 and Kpn2, though also ST147, appeared to form a distinct cluster, evidenced by a substantial genetic difference from Kpn24. Furthermore, Kpn2 shows significant genetic relatedness with an NDM-1-producing K. pneumoniae clone accountable for an outbreak in Italy (29), named Kpn135LU, diverging by 34 SNPs. Additionally, a high degree of similarity was observed between the two ST225 isolates. Notably, the novel ST9095 isolate displayed considerable genetic diversity compared to the others (Fig. 3A). Regarding E. coli ST648, Ecoli15 and Ecoli17 exhibited strong genetic relatedness (6 SNPs different), while Ecoli16 showed greater genetic divergence (13 SNPs different) (Fig. 3B).
Minimum spanning tree of K. pneumoniae (A) and Escherichia coli (B) isolates based on cgSNP analysis. Each node represents a distinct isolate and the colors of the nodes denote different STs. Numbers on lines represent the number of allelic differences between isolates. This figure was created using BioRender (https://www.biorender.com/).
DISCUSSION
MBL-harboring Enterobacterales represent a concerning and growing issue, especially due to their limited treatment options. The epidemiology of carbapenem-resistance mechanisms is shifting, with a progressive spread of NDM-carrying bacteria across Europe, particularly in Italy, where an increasing number of NDM-producing isolates has been reported (9, 10). In this work, conducted in our transplant center, we characterized the clinical, epidemiological, and genomic features of resistance associated with MBL-producing strains isolated from critically ill patients, using phenotypic tests combined with WGS analysis. The most frequently detected MBL was NDM (82% of isolates), with NDM-1 being the predominant variant, followed by NDM-5 found exclusively in E. coli isolates. VIM-1 was the only VIM variant detected (18% of isolates). These results align with existing literature that highlights NDM-1 as the most prevalent NDM variant in Enterobacteriaceae across European countries, including Italy (30), and NDM-5 variant, exhibiting notable geographic variations, with an endemic presence observed in China (31), and cases sporadically reported in Europe (32, 33). The presence of VIM-producing Enterobacteriaceae in our cohort is particularly concerning, as two of these strains were associated with fatal infections. This finding is significant given their reported association with hospital outbreaks (34, 35) and their relatively uncommon occurrence in Enterobacteriaceae, compared to P. aeruginosa (36). Interestingly, in all the strains, MBL genes had a plasmid localization.
Beyond the expected resistance to cephalosporins, BL/BLI combinations, and carbapenems, due to MBL production, many other antimicrobial agents demonstrated no in vitro effectiveness, reducing further therapeutic options. Although MBLs are not able to hydrolyze monobactams, a significant proportion (95%) of the analyzed isolates exhibited aztreonam resistance, likely mediated by class A β-lactamases, including extended-spectrum β-lactamase (ESBLs) and KPCs, which are able to easily inactivate this antibiotic (37). The in vitro effectiveness shown by aztreonam-avibactam suggests this antibiotic combination as a promising therapeutic strategy. Resistance to aminoglycosides was also prevalent, with 63% of the isolates resistant to amikacin. Amikacin is often used for MDR Gram-negative bacterial infections treatment, including urinary tract infections (18), particularly when alternative therapeutic agents like cefiderocol are not available. In our study, for instance, 31.8% of patients received an aminoglycoside as part of their empirical antibiotic regimen.
Despite high susceptibility rates being limited to colistin and cefiderocol, we observed the emergence of colistin resistance in all VIM-1-producing K. pneumoniae and a relevant number of K. pneumoniae isolates showing cefiderocol MICs very close to the EUCAST breakpoint, indicating a reduced susceptibility to this antibiotic. Our results reflect a phenomenon observed in several studies, which report a higher prevalence (42%–59%) of NDMs among cefiderocol-resistant isolates (38, 39). Furthermore, MBLs, such as IMP, VIM, and NDM, can marginally hydrolyze cefiderocol and lead to an increased MIC (40, 41). This is supported by the fact that inhibition of MBL activity using dipicolinic acid prevented the emergence of cefiderocol-resistant mutants (42). On the other hand, we cannot exclude the involvement of other factors not investigated in this study, such as the overexpression of various class A β-lactamases, mainly KPC and SHV, which have been reported to contribute to cefiderocol resistance (43). Although we found only one strain (Kpn1) resistant to cefiderocol, it is important to highlight that cefiderocol resistance has been reported in a large outbreak in Tuscany, Italy, recently (8).
Genotypic analysis revealed a significant presence of concerning high-risk clones (44–46) among the isolates circulating in our hospital. MLST analysis revealed that the most frequent K. pneumoniae STs were ST147, known to be responsible for several outbreaks in European countries (Italy, Greece, and, more recently, Spain [47]) and North Africa and India (44, 48), followed by ST512. The most prevalent E. coli ST was ST648, described to become a high-risk clonal lineage, alarmingly circulating worldwide (49, 50), and to worsen infection treatment possibilities (51). Besides them, we found other STs, including one not previously described. The substantial heterogeneity in ST distribution strongly suggests multiple independent acquisitions, and no evidence of a defined outbreak was identified during the investigation.
According to the phylogenetic analysis, two significant potential clusters within the K. pneumoniae group were present, one comprising all four ST512 isolates and another one consisting of six out of eight ST147 isolates, both showing strong genetic relatedness. Remarkably, one of the ST147 isolates (Kpn2) shows significant genetic relatedness with an NDM-1-producing K. pneumoniae clone responsible for a large outbreak in Tuscany, Italy (29). Notably, the significant genetic diversity shown by the novel ST9095 isolate suggests the new ST is unlikely to derive from other detected strains. Our data, especially the intra-ST147 diversity, show that MLST alone may not be sufficient for a detailed epidemiological tracing, while the cgSNP analysis appeared to be crucial.
The resistome analysis revealed a complex landscape of antibiotic resistance mechanisms among the isolates analyzed. Several ARGs appear to be particularly widespread. For instance, genes associated with resistance to β-lactams (e.g., SHV, TEM, CTX-M, KPC, and OXA members) are prevalent in many K. pneumoniae isolates. Among the genes mentioned above, blaSHV-11 encoding β-lactamases has been involved in drug resistance mechanisms (52) through enzymatic inactivation of antimicrobials (53). Similarly, the co-presence of genes conferring resistance to aminoglycosides (aac, ant, aph), fluoroquinolones (qnr), sulfonamide (sul), and trimethoprim (dfr) was often detected. In general, a higher number of genes conferring resistance to aminoglycosides was found in K. pneumoniae isolates, compared to E. coli.
Of note, among the NDM-producing K. pneumoniae (ST147, ST395) and E. coli (ST648 and ST404) strains examined in this study, the concomitant presence of ESBL, including CTX-M-15, OXA, SHV, and TEM variants, was observed. Overall, our findings suggest a general correspondence between genomic resistance profiles and phenotypic antibiogram. However, while the absence of resistance genes often predicts susceptibility, the mere presence of genetic determinants is not always sufficient to confer resistance to an antibiotic, and this is probably due to the complex and multifactorial mechanisms underlying antimicrobial resistance profiles. For instance, the molecular mechanisms leading to colistin resistance involve multiple interacting factors, mainly centered on LPS modification (54). One such factor is the plasmid-borne mobile colistin resistance gene (mcr-1), which was not detected in isolates analyzed here. Conversely, WGS revealed a widespread diffusion of pmrF, arnT, and ept gene variants, reported to potentially confer polymyxin resistance (55, 56). The low rate of colistin resistance observed in this study could be explained by their different expression levels, which were not tested. Furthermore, other resistance strategies, such as efflux pumps and capsule formation (57), may also be implicated in these isolates’ resistance mechanisms.
All the isolates characterized in this study harbored several virulence factors, with significant clinical implications. Particularly, siderophore systems, involved in bacterial growth and pathogenesis (58, 59), were consistently detected, mainly in K. pneumoniae isolates and E. coli ST1485 and ST405, which harbored the highest number of virulence factors. All the E. coli isolates carried factors involved in colonization and biofilm formation (ECP and HCP) (60), determinants involved in adhesion (laminin-binding fimbriae) (60), proteins with toxic effects (hemolysin), and invasion proteins. In particular, the Ecoli23 strain was also characterized by 39P fimbria playing a role in ExPEC pathogenicity (61). Strikingly, this was the only E. coli strain in our study that resulted in patient death, whose infection was identified as the primary cause. The presence of these virulence determinants may explain why 40% (2/5) of the patients infected with MBL-producing E. coli developed a subsequent infection following the initial isolation.
All the K. pneumoniae isolates were positive for type III and I fimbriae, which facilitate bacterial adhesion (62), the AcrAB efflux pump involved in expelling antibiotics and antimicrobial peptides (63), and rcsAB, a regulator of capsule synthesis, as well as siderophores. Furthermore, the regulator of mucoid phenotype, rmp (64), was detected in some of the strains. Finally, through a bioinformatic tool, a very high virulence score was assigned to five strains (Kpn1, Kpn2, Kpn3, Kpn5, and Kpn8). Four of these high-virulence isolates (Kpn2, Kpn3, Kpn5, and Kpn8) belong to ST147, well known as a global MDR high-risk clone (44). These isolates were associated with subsequent infections in patients, resulting in two cases of patient death, highlighting the significant clinical impact of such strains on the clinical outcome.
Overall, our findings revealed an alarming co-presence of multidrug resistance and virulence among MBL-producing Enterobacterales, particularly for K. pneumoniae. Virulome profiling identified a broad repertoire of virulence factors, including siderophores and fimbriae, with several strains linked to subsequent infections and poor clinical outcomes. Notably, widespread resistance determinants, including those conferring resistance to aminoglycosides, fluoroquinolones, and last-resort agents like colistin and cefiderocol, severely narrow therapeutic options. In addition to colistin-resistant strains, we observed a worrying trend of reduced susceptibility to cefiderocol, which is particularly relevant given its growing role in the treatment of MBL-producing organisms, thus warranting close monitoring.
Although the limitation of the study resides in the relatively small sample size and the monocentric design, our findings offer valuable insights into the genomic and clinical landscape of MBL-producing pathogens in this specific patient population. Bigger and multicenter studies may be useful to better define genetic and virulence-associated factors linked to poor outcomes and ultimately guide earlier, more targeted interventions to improve patient survival.
In conclusion, genomic analyses are crucial for a prompt understanding of epidemiological dynamics and resistance mechanisms. Resistome and virulome information, obtained with WGS, could be used to develop new infection control protocols, able to track hypervirulent MDR strains associated with difficult-to-treat infections and hospital outbreaks.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Logan LK, Weinstein RA. 2017. The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of a global menace. J Infect Dis 215:S 28–S 36. doi:10.1093/infdis/jiw 28228375512 PMC 5853342 · doi ↗ · pubmed ↗
- 2Boyd SE, Livermore DM, Hooper DC, Hope WW. 2020. Metallo-β-lactamases: structure, function, epidemiology, treatment options, and the development pipeline. Antimicrob Agents Chemother 64:e 00397-20. doi:10.1128/AAC.00397-2032690645 PMC 7508574 · doi ↗ · pubmed ↗
- 3Meini MR, Llarrull LI, Vila AJ. 2014. Evolution of metallo-β-lactamases: trends revealed by natural diversity and in vitro evolution. Antibiotics (Basel) 3:285–316. doi:10.3390/antibiotics 303028525364574 PMC 4212336 · doi ↗ · pubmed ↗
- 4Logan LK, Bonomo RA. 2016. Metallo-β-lactamase (MBL)-producing Enterobacteriaceae in United States children. Open Forum Infect Dis 3:ofw 090. doi:10.1093/ofid/ofw 09027419164 PMC 4943557 · doi ↗ · pubmed ↗
- 5Brolund A, Lagerqvist N, Byfors S, Struelens MJ, Monnet DL, Albiger B, Kohlenberg A. 2019. Worsening epidemiological situation of carbapenemase-producing Enterobacteriaceae in Europe, assessment by national experts from 37 countries, July 2018. Euro Surveill 24. doi:10.2807/1560-7917.ES.2019.24.9.1900123 PMC 640217730862330 · doi ↗ · pubmed ↗
- 6Galani I, Souli M, Chryssouli Z, Katsala D, Giamarellou H. 2004. First identification of an Escherichia coli clinical isolate producing both metallo-beta-lactamase VIM-2 and extended-spectrum beta-lactamase IBC-1. Clin Microbiol Infect 10:757–760. doi:10.1111/j.1469-0691.2004.00913.x 15301681 · doi ↗ · pubmed ↗
- 7Lynch JP III, Clark NM, Zhanel GG. 2021. Escalating antimicrobial resistance among Enterobacteriaceae: focus on carbapenemases. Expert Opin Pharmacother 22:1455–1474. doi:10.1080/14656566.2021.190489133823714 · doi ↗ · pubmed ↗
- 8Tavoschi L, Forni S, Porretta A, Righi L, Pieralli F, Menichetti F, Falcone M, Gemignani G, Sani S, Vivani P, et al.. 2020. Prolonged outbreak of New Delhi metallo-beta-lactamase-producing carbapenem-resistant Enterobacterales (NDM-CRE), Tuscany, Italy, 2018 to 2019. Euro Surveill 25. doi:10.2807/1560-7917.ES.2020.25.6.2000085 PMC 702944732070467 · doi ↗ · pubmed ↗