Fusarium incarnatum: a paradigm for One Health pathogen dynamics across humans, animals, and the environment
Ahmed Namisy, Ying-Chen Wu, Kuo-Hsi Lin, Wei-Che Hsu, Wen-Hsin Chung

TL;DR
This paper explores how a plant pathogen, Fusarium incarnatum-equiseti, can infect humans, animals, and plants, highlighting its public health and agricultural risks.
Contribution
The study identifies Fusarium pernambucanum and Fusarium irregulare as newly reported pathogens in Taiwan capable of cross-infection across species.
Findings
FIESC isolates belong to the Incarnatum clade and include F. irregulare and F. pernambucanum.
Isolates can infect human nails, animal skin, and plant fruits, showing cross-species adaptability.
Plant isolates PLMF1 and BDMF7 transition from plant pathogens to zoonotic pathogens.
Abstract
The Fusarium incarnatum-equiseti species complex (FIESC) is a harmful plant pathogen responsible for diseases, such as root rot, fruit rot, leaf blight, and wilt, in various crops. Recently, it has been increasingly associated with both superficial and invasive infections in humans and animals. Additionally, this pathogen produces mycotoxins, posing significant risks to public health. This study aims to characterize the molecular and morphological characteristics of FIESC isolates obtained from various sources, including human, veterinary, and plant samples. Molecular identification was performed using multilocus sequence typing (MLST), targeting the internal transcribed spacer (ITS) rDNA, the second-largest subunit of the RNA polymerase gene (RPB2), and translation elongation factor 1 alpha (TEF-1α). Based on MLST analysis, all FIESC isolates were found to belong to the Incarnatum…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1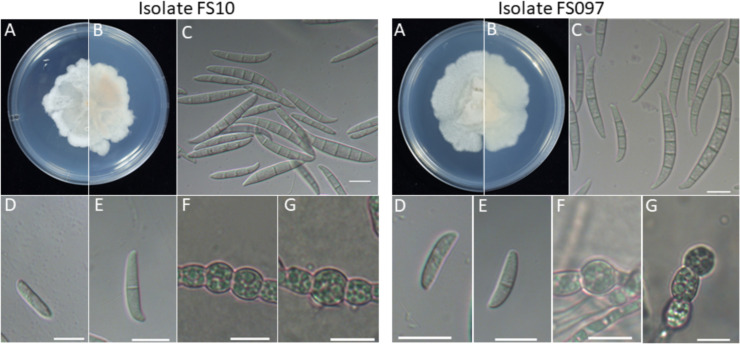 Fig 2
Fig 2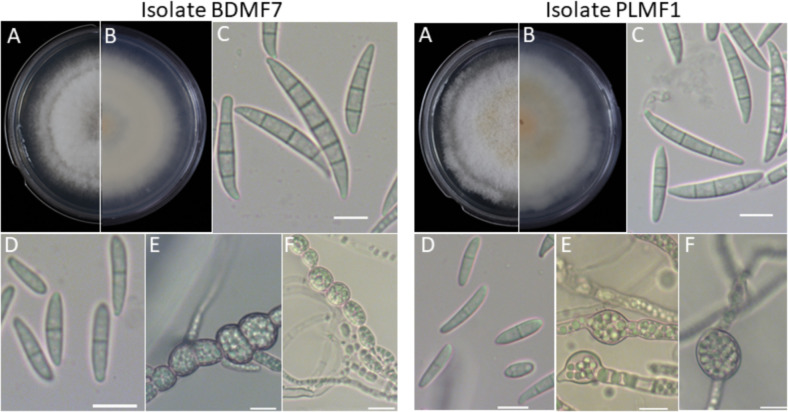 Fig 3
Fig 3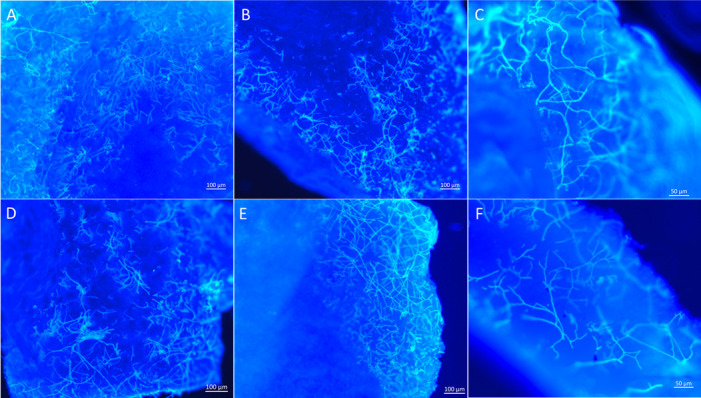 Fig 4
Fig 4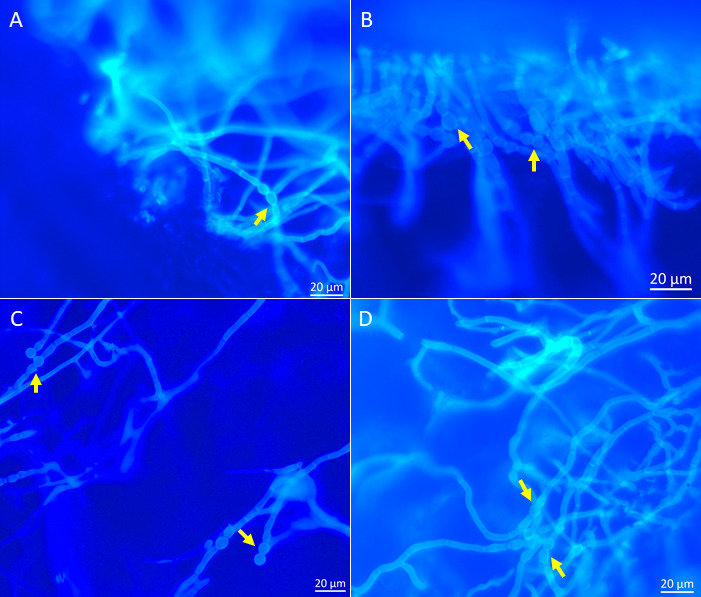 Fig 5
Fig 5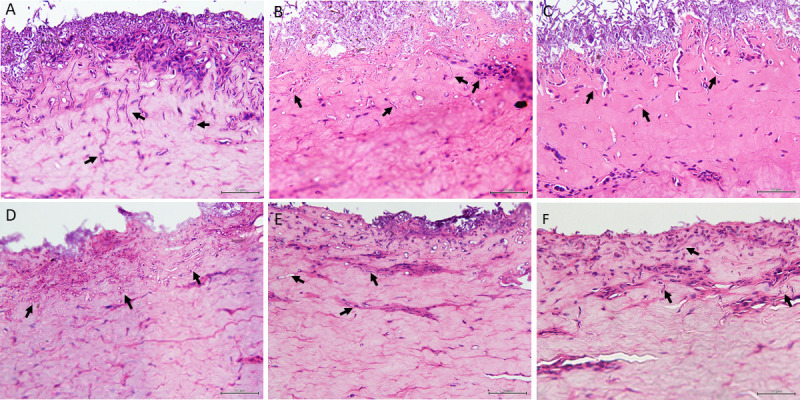 Fig 6
Fig 6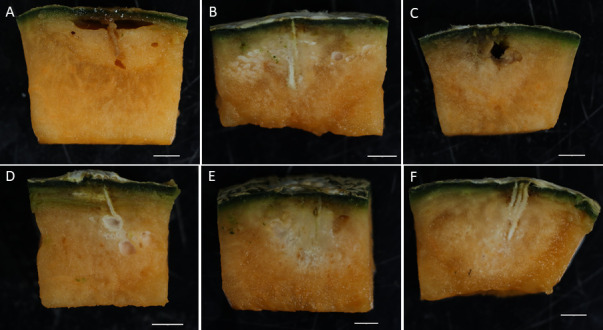 Fig 7
Fig 7| GenBank accession | ||||||
|---|---|---|---|---|---|---|
| Isolate | Host | Origin | ITS | TEF 1-α | RPB2 | |
|
| FS097 | Rabbit nose | Taiwan |
|
|
|
| LD1 | Cat lung | Taiwan |
|
|
| |
| LC7188 | Bamboo | China |
|
|
| |
| NRRL 34001 | Human foot wound | Texas |
|
|
| |
| PLMF1 | Melon | Taiwan |
|
|
| |
|
| FS405 | Turtles | Taiwan |
|
|
|
| FS10 | Human sputum | Taiwan |
|
|
| |
| NRRL 32864 | Human | Texas |
|
|
| |
| NRRL 36548 | Banana | Congo |
|
|
| |
| NRRL 34070 | Tortoise | Illinois |
|
|
| |
| BDMF7 | melon | Taiwan |
|
|
| |
|
| NRRL 34056 | Human bronchial wash | Illinois |
|
|
|
| NRRL 43730 | Contact lens | Mississippi |
|
|
| |
| NRRL 34059 | Human blood | Illinois |
|
|
| |
|
| NRRL 31167 | Human sputum | Texas |
|
|
|
| LC12167 | Luffa | China |
|
|
| |
|
| NRRL 36575 | Juniperus chinensis leaf | Hawaii |
|
|
|
| NRRL 34003 | Human sputum | Texas |
|
|
| |
|
| NRRL 43297 | Spartina rhizomes | Connecticut |
|
|
|
| NRRL 34005 | Human intravitreal fluid | Minnesota |
|
|
| |
|
| NRRL 32867 | Human | Texas |
|
|
|
| NRRL 13379 |
| India |
|
|
| |
|
| LC12168 |
| China |
|
|
|
| LC1385 | Saudi Arabia | Tomato |
|
|
| |
|
| NRRL 3214 | Unknown | Unknown |
|
|
|
| NRRL 3020 | Unknown | Unknown |
|
|
| |
|
| NRRL 36136 | Unknown | Unknown |
|
|
|
| NRRL 26419 | Soil | Germany |
|
|
| |
|
| NRRL 34032 | Human abscess | Texas |
|
|
|
| NRRL 32871 | Human abscess | Texas |
|
|
| |
|
| NRRL 31011 | Germany |
|
|
| |
| NRRL 26921 | Wheat | Germany |
|
|
| |
|
| NRRL 43638 | Manatee | Florida |
|
|
|
| NRRL 43694 | Human eye | Texas |
|
|
| |
|
| NRRL 36448 | Phaseolus vulgaris seed | Sudan |
|
|
|
| NRRL 36401 | Cotton | Mozambique |
|
|
| |
|
| NRRL 36323 | Cotton yarn | England |
|
|
|
| NRRL 28029 | Human eye | California |
|
|
| |
|
| NRRL 13459 | Plant debris | South Africa |
|
|
|
| Penetration depth | ||||
|---|---|---|---|---|
| Isolates | Host | Porcine skin (µm) | Muskmelon fruit (mm) | |
| PLMF1 |
| Muskmelon fruit | 109.5 ± 7.6 d | 20.1 ± 0.4 a |
| LD1 |
| Cat | 146.8 ± 2.8 c | 17.7 ± 0.3 b |
| FS097 |
| Rabbit | 190.6 ± 3.5 b | 17.0 ± 0.6 b |
| FS405 |
| Turtle | 191.4 ± 5.1 b | 20.8 ± 0.4 a |
| FS10 |
| Human | 223.7 ± 4.8 a | 18.4 ± 0.6 b |
| BDMF7 |
| Muskmelon fruit | 84.90 ± 2.3 e | 21.5 ± 0.2 a |
- —National Science and Technology Council, Taiwan, R.O.C.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycotoxins in Agriculture and Food · Plant-Microbe Interactions and Immunity
INTRODUCTION
The fungal genus Fusarium is one of the most significant groups of plant-pathogenic fungi, affecting a wide range of crops worldwide (1). These fungi are soil-borne diseases and can persist for long periods in the soil, surviving on crop residues and the roots of non-host plants as chlamydospores (2). Fusarium is widespread due to its ability to grow on various substrates and effective dispersal mechanisms (3). Additionally, Fusarium species have been identified in the water distribution systems of hospitals and residences, where they can survive for years (4). Currently, Fusarium is ranked among the 10 most destructive fungal plant pathogens (5). With a broad host range, Fusarium spp. can cause various diseases in many plant species. The pathogen infects certain parts of plants, such as grains, seedlings, heads, roots, and stems, leading to various diseases that reduce product quality and diminish crop yields (6). The genus Fusarium comprises at least 300 species grouped into approximately 23 species complexes based on multi-locus phylogenetic analyses (7). Fusarium species complexes, associated with human and animal infections, include the F. incarnatum-equiseti species complex (FIESC), F. tricinctum species complex (FTSC), F. sambucinum species complex (FSAMSC), F. solani species complex (FSSC), F. fujikuroi species complex (FFSC), F. oxysporum species complex (FOSC), F. dimerum species complex (FDSC), and F. chlamydosporum species complex (FCSC) (8).
In humans, Fusarium species is recognized as the second most common mold infection after Aspergillus (8). The most common infections caused by Fusarium are superficial, localized diseases such as keratitis and onychomycosis in immunocompetent patients (9). However, in severely immunocompromised patients, Fusarium can cause locally invasive or disseminated infections (9), particularly in those who have undergone allogeneic hematopoietic stem cell transplantation (HSCT) due to significant immunosuppression and prolonged neutropenia (8). Invasive disease caused by Fusarium spp. includes cutaneous infection, chronic rhinosinusitis, pneumonia, brain abscess, and fatal fungemia (10). Fusarium species commonly cause opportunistic infections in animals, including seahorses, turtles, dolphins, dogs, horses, and pigs (6, 11). It is responsible for various animal mycotic infections, such as keratitis and dermatitis (12). In addition, Fusarium spp. have been reported to cause systemic infection in goats and dogs, including meningoencephalitis, chronic ulcerative dermatitis, and lesions in the kidneys, abdominal lymph nodes, and lungs (11, 13, 14).
Besides the direct infection of plants, humans, and animals, Fusarium spp. can also produce mycotoxins, including trichothecenes and zearalenone (15, 16). Fusarium mycotoxins can infiltrate the food chain through contamination during crop production and food processing, ultimately posing a threat to human and animal health (17). These mycotoxins contribute to worldwide losses amounting to millions of dollars annually due to their impact on human health, animal health, and condemned agricultural products (16). The most significant effect of Fusarium species on livestock arises from feed contaminated with mycotoxins (18). Poultry, pigs, and aquatic vertebrates are highly sensitive to mycotoxins due to their high consumption rates and chronic exposure to cereal mycotoxins (18).
Currently, the cross-kingdom infection of Fusarium spp. in plants and humans has been reported in previous studies. Fusarium spp. isolated from keratitis patients retain their infective capacity during the cross-kingdom reinfection transmission from humans to plants (19). For example, members of F. solani are host-specific pathogens with an extensive host range of agriculturally important crops, such as potato, tomato, pepper, and eggplant (20). However, they are also increasingly recognized as causative agents of human and animal mycoses (21). Moreover, FIESC has been reported to cause infections in humans, animals, and plants (12, 22, 23). These cross-infections pose serious threats to food safety and public health. In this study, FIESC was successfully isolated from various sources, including human, veterinary, and agricultural environments in Taiwan. These isolates were analyzed phylogenetically using multilocus sequence typing (MLST), targeting the internal transcribed spacer (ITS) rDNA, the second-largest subunit of the RNA polymerase gene (RPB2), and translation elongation factor 1 alpha (TEF-1α). Additionally, we investigated the potential for cross-infection of these isolates in plants, human nails, and porcine skin.
MATERIALS AND METHODS
Pathogen isolates
This study analyzed six Fusarium isolates belonging to FIESC that were obtained from human, veterinary, and plant sources (Table 1). The isolate FS10 was recovered from a sputum sample of a patient diagnosed with fungal pneumonia at Tungs’ Taichung MetroHarbor Hospital, Taichung, Taiwan. The patient had a history of liver transplantation, and after antifungal treatment, the symptoms resolved. Veterinary isolates LD1, FS097, and FS405 were recovered from three different animal species, all presenting with respiratory clinical signs, at the clinical microbiology laboratory of the Research Center for Animal Medicine in Taichung, Taiwan. Specifically, isolate LD1 was cultured from a cat’s lung via bronchoalveolar lavage, isolate FS097 from a rabbit’s purulent nasal discharge, and isolate FS405 from a turtle’s (Chelonoidis denticulata) tracheal discharge via tracheal wash. Two plant isolates, PLMF1 and BDMF7, obtained from rotted melon fruits, were generously provided by the Plant Parasites and Pesticide Resistance Molecular Diagnosis Laboratory (PRMD), National Chung Hsing University, Taichung, Taiwan. The PRMD lab previously identified PLMF1 and BDMF7 as Fusarium incarnatum-equiseti species complex, based on their morphological and molecular characteristics.
Phylogenetic analysis
The genomic DNA was extracted from mycelium grown on PDA medium for one week using the DNeasy Blood and Tissue Kit (QIAGEN GmbH, Hilden, Germany). The internal transcribed spacer (ITS) rDNA, translation elongation factor 1-α (TEF 1-α) gene, and RNA polymerase II second largest subunit (RPB2) gene were PCR amplified and sequenced using primers of ITS1/ITS4 (24), EF1/EF2 (25), and 5F2/7CR (26), respectively. The PCR was performed in a 25 µL reaction volume, consisting of 0.5 µL (10 µmol/L) of each forward and reverse primer, 5 µL of PCR Master Mix (BioKit, Taiwan), 18 µL of nuclease-free water, and 1 µL of DNA template. PCR amplifications were conducted using a Labcycler 48 (SensoQuest GmbH, Göttingen, Germany) under the conditions described by White et al. (24), Carbone and Kohn (25), and Liu et al. (26) for ITS, TEF 1-α, and 5F2/7CR, respectively. The PCR products were analyzed via 1.5% agarose gel electrophoresis, visualized, and photographed under UV light. The phylogeny of six FIESC isolates obtained in this study, along with 33 reference sequences from GenBank (Table 1), was analyzed using multi-locus sequence typing (MLST). Sequences were assembled using the BioEdit Sequence Alignment Editor (27) and aligned using Clustal W. Minor gaps were manually removed using Molecular Evolutionary Genetics Analysis software version 7.0 (MEGA 7.0). Phylogenetic analysis was performed using the maximum likelihood method with 1,000 bootstrap replicates.
Morphological characteristic
A single-spore culture was used to examine the morphological characteristics of FIESC isolates. Colony pigmentation was assessed after culturing the isolates on Potato Dextrose Agar (PDA) plates and incubating at 28°C for one week. Approximately 30 individual conidia were measured for length, width, and number of septa in microconidia and macroconidia, cultured on Spezieller Nahrstoffarmer Agar (SNA) medium, and incubated in the dark at 28°C for one week. Conidial characteristics were analyzed from images captured using an Axioplan 2 imaging microscope (Zeiss, Germany).
Inoculum preparation
Six Fusarium isolates collected from human, veterinary, and plant sources were used to investigate their ability to cross-infect, invade, and colonize human nails, porcine skin, and muskmelon fruit. Conidial suspensions were prepared as described by Namisy et al. (28). Briefly, a single spore from each isolate was cultured on PDA medium and incubated at 28°C for one week. The conidia were then washed once with sterile distilled water (SDW). Mycelia were removed by filtration through a Miracloth filter (MilliporeSigma, Burlington, MA, USA) to prepare the conidial suspension, and the concentration was adjusted to 10⁶ conidia/mL for all inoculation treatments.
Inoculation assay
The ex vivo infection model was used to study the colonization of Fusarium isolates in human nails. Nails were collected from male and female volunteers, disinfected with 70% ethanol for 1 min, and rinsed three times with sterile distilled water (SDW). After drying on sterile paper under laminar airflow, the sterilized nails were soaked in an Eppendorf tube containing conidial suspension for 30 s. Mock controls were inoculated with SDW. The nails were then placed in sterile Petri dishes and incubated at 28°C for 4 days. Fusarium invasion in infected nails was assessed at 4 days post-inoculation (dpi) using an Axio Imager A1 microscope (Carl Zeiss AG, Jena, Germany) equipped with an X-Cite 120Q fluorescence illuminator system (Excelitas Technologies, USA) and an Axiocam 506 color camera. Nail sections were manually prepared, placed on glass slides, and stained with 10 µL of Calcofluor White solution (Sigma) and 10 µL of 10% potassium hydroxide solution to enhance resolution. The slides were incubated at room temperature in the dark for 1 min. Fluorescence expression of Fusarium isolates was visualized using a DAPI filter with excitation at 320–390 nm and emission at 430–490 nm. The experiment was performed in triplicate, repeated twice, and approximately 10 slices per replication were observed.
In the animal tissue inoculation assay, an ex vivo infection model using porcine skin was employed. Fresh skin samples were excised from euthanized animals, cut into small pieces (~2 cm²), disinfected with 70% ethanol for 5 min, and then rinsed three times with sterile distilled water (SDW) before drying under a biosafety cabinet. The samples were wounded using a sterile 24G needle, placed in Petri dishes, and inoculated with 20 µL of conidial suspension from each Fusarium isolate on the upper layer of the porcine skin epidermis. Mock controls were inoculated with SDW. The infected tissues and controls were incubated at 28°C for 4 days. The histological examination was performed according to standard protocols as described by Feldman and Wolfe (29). Infected tissues were fixed in 10% formaldehyde for 24 h, embedded in paraffin, sectioned into 3 μm thick slices using a microtome, and stained with hematoxylin and eosin (H&E). Fungal invasion in stained sections was observed using an Axio Imager A1 microscope (Carl Zeiss AG, Jena, Germany).
To investigate the colonization of Fusarium isolates in plants, healthy muskmelon fruits were surface-disinfected with 70% ethanol for 30 s, followed by 1% NaOCl for 5 min, and then rinsed three times with sterile distilled water (SDW). The fruits were wounded using a sterile needle and inoculated with 10 µL (concentration 10^6^ conidia/mL) of conidial suspension, while mock controls were inoculated with SDW. The fruits were monitored for symptoms 10 days after incubation at 28°C.
Statistical analysis
Data analyses were conducted using the R Statistical Software v4.1.3 (https://www.r-project.org/). To analyze the statistical significance of penetration depth of each Fusarium isolate, we performed a one-way analysis of variance followed by Fisher’s least significant difference multiple comparisons test at (P < 0.05).
RESULTS
Phylogenetic analysis
The phylogenetic analysis was conducted based on the combined sequences of the ITS region, TEF 1-α, and RPB2 to classify six Fusarium isolates belonging to the Fusarium incarnatum-equiseti species complex (FIESC), which were isolated from human, veterinary, and plant sources. These sequences were compared with 33 reference sequences from GenBank, with Fusarium concolor (NRRL 13459) used as the outgroup (Table 1). The results indicated that the six isolates in this study were divided into two groups. Two isolates, LD1 and FS097, obtained from veterinary sources, and one isolate PLMF1 from a plant source clustered with F. irregulare, with 99% maximum likelihood (ML) bootstrap support. Similarly, two isolates, FS405 and FS10, obtained from veterinary and human sources, respectively, along with one isolate BDMF7 from a plant source, clustered with F. pernambucanum, also supported by 99% mL bootstrap values (Fig. 1).
Phylogenetic tree of Fusarium incarnatum-equiseti species complex (FIESC) based on sequencing analysis of ITS, TEF 1-α, and RPB2 using the maximum likelihood method. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) is shown.
Morphological characteristic
To further investigate their morphological characteristics, the FS097 and FS10 were selected as representative isolates and compared their morphology with same species of PLMF1 and BDMF7, isolates obtained from plant sources. Single-spore isolates assessed morphological characteristics, including the shape, size, and septation of microconidia and macroconidia. The colony of F. irregulare veterinary isolate FS097 exhibited cottony white mycelium, which developed white-brown pigmentation over time. Microconidia were ovoid to fusiform, or slightly curved, with 0 to 1 septum, measuring 2.4–3.7 µm × 10.1–16.5 µm. Macroconidia were 3 to 6-septate, with a slightly curved apical cell that ended bluntly, ranging in size from 3.2 to 4.8 µm × 24.6–43.0 µm. Spherical or oval chlamydospores with thick walls were abundant, occurring singly or in chains and produced in a terminal or intercalary position. These chlamydospores were developed through the modification of hyphal and conidial cells (Fig. 2).
General morphology of Fusarium pernambucanum human isolate FS10 and Fusarium irregulare veterinary isolate FS097. (A, B) Forward and reverse colony morphology on PDA medium; (C) macroconidia; (D, E) microconidia; and (F, G) chlamydospores. Scale bar: 10 µm.
Additionally, the F. irregulare plant isolate PLMF1 formed a colony with a white cottony appearance and developed pale yellow pigmentation. Microconidia were ovoid or slightly curved, with 0 to 1 septum, ranging in size from 2.1 to 3.7 µm × 12.0–17.9 µm. Macroconidia were mostly 3-septate and rarely 5 and 6-septate, with a slightly curved apical cell that ended bluntly, measuring 3.4–4.8 µm × 25.2–38.3 µm. Oval chlamydospores were abundant, occurring singly and produced in either terminal or intercalary positions, developing from hyphae (Fig. 3).
General morphology of Fusarium pernambucanum plant isolate BDMF7 and Fusarium irregulare plant isolate PLMF1. (A, B) Forward and reverse colony morphology on PDA medium; (C) macroconidia; (D) microconidia; and (E, F) chlamydospores. Scale bar: 10 µm.
Similarly, the colony of the F. pernambucanum human isolate FS10 exhibited similarities to the veterinary isolate FS097. Microconidia were ovoid or slightly curved, with 0 to 1 septum, ranging in size from 2.0 to 3.6 µm × 11.5–16.8 µm. Most macroconidia were 3 to 5-septate, with rare occurrences of 7- and 11-septate. They featured a slightly curved apical cell that ended bluntly and measured 3.3–4.5 µm × 22.0–39.0 µm. Spherical chlamydospores occurred singly or in chains, produced in an intercalary position, and developed from hyphae (Fig. 2).
The colony of the F. pernambucanum plant isolate BDMF7 was cottony white with pale yellow pigmentation. Microconidia were ovoid or slightly curved, with 0 to 1 septum, ranging in size from 2.0 to 3.1 µm × 11.0–16.9 µm. Macroconidia were primarily 3-septate, with a few exhibiting 5 septa. The apical cell was blunt at the apex and measured 3.5–5.5 µm × 22.3–37.0 µm. Spherical chlamydospores were developed from hyphae and produced singly in terminal or intercalary positions (Fig. 3).
Inoculation assay
In this study, we evaluated the potential for cross-infection in human nails, porcine skin tissue, and muskmelon fruits using F. irregulare and F. pernambucanum isolates obtained from various sources, including the human isolate FS10; veterinary isolates FS097, LD1, and FS405; and plant isolates PLMF1 and BDMF7.
The nail infection assay results demonstrated that all Fusarium isolates could grow on the nail surface within 4 days of incubation at 28°C. Additionally, histological analysis confirmed that all isolates could invade the nail plate (Fig. 4). However, the penetration rate varied among the different Fusarium isolates. The human and veterinary isolates exhibited a higher penetration density at 4 days post-inoculation compared to the plant isolates. Thick hyphae and chlamydospores were observed throughout the nail (Fig. 5). However, microconidia and macroconidia were not detected at 4 days post-inoculation.
Histopathological characteristics of Fusarium isolates in human nail at 4 dpi. (A) Fusarium pernambucanum human isolate FS10; (B) Fusarium pernambucanum veterinary isolate FS405; (C) Fusarium irregulare veterinary isolate FS097; (D) Fusarium irregulare veterinary isolate LD1; (E) Fusarium irregulare plant isolate PLMF1; and (F) Fusarium pernambucanum plant isolate BDMF7.
Chlamydospores in the human nail infected with Fusarium isolates at 4 dpi. (A) Fusarium irregulare veterinary isolate FS097; (B) Fusarium pernambucanum veterinary isolate FS405; (C) Fusarium pernambucanum plant isolate BDMF7; and (D) Fusarium irregulare plant isolate PLMF1.
In the porcine skin infection assay, an ex vivo infection model using porcine skin was employed to study the invasion and colonization of Fusarium isolates. The results indicated that all Fusarium isolates could colonize the surface of tissue samples within 2 days post-inoculation. The human isolate FS10 and veterinary isolates FS097, LD1, and FS405 exhibited abundant growth, whereas the plant isolates BDMF1 and PLMF1 showed lower growth, with BDMF1 displaying the least. Additionally, at 2 days post-inoculation, histological analysis confirmed that the pathogen could penetrate the epidermis layer of porcine skin. By 4 days post-inoculation, the penetration of all isolates had extended into the dermis layer (Fig. 6).
Histopathological characteristics of Fusarium isolates in porcine skin at 4 dpi. (A) Fusarium pernambucanum human isolate FS10; (B) Fusarium pernambucanum veterinary isolate FS405; (C) Fusarium irregulare veterinary isolate FS097; (D) Fusarium irregulare veterinary isolate LD1; (E) Fusarium irregulare plant isolate PLMF1; and (F) Fusarium pernambucanum plant isolate BDMF7.
However, significant differences in penetration depth were observed among the Fusarium isolates. The human isolate FS10 displayed the most significant penetration depth, measuring 223.77 µm, followed by the veterinary isolates FS405 (191.43 µm), FS097 (190.65 µm), and LD1 (146.86 µm) (Table 2).
Similar to infections observed in human nails and porcine skin, the plant pathogenicity assay also confirmed that all Fusarium isolates were pathogenic to muskmelon fruits at 10 days post-inoculation (dpi) (Fig. 7). Symptoms initially appeared as water-soaked lesions at the inoculation site. Over time, white, cotton-like mycelia developed on the fruit’s surface, progressively expanding and invading internal tissues, ultimately leading to discoloration and fruit breakdown. Additionally, the pathogen spread and colonized seeds within infected fruits. Pathogens were successfully reisolated from infected fruit and confirmed through morphological and molecular analysis. Among the isolates, F. pernambucanum plant isolates BDMF7 and veterinary isolate FS405 exhibited the greatest penetration depths, measuring 21.58 mm and 20.85 mm, respectively. Similarly, the F. irregulare plant isolate PLMF1 showed a significant penetration depth of 20.1 mm. In contrast, the F. pernambucanum human isolate FS10 and F. irregulare veterinary isolates LD1 and FS097 demonstrated the lowest penetration depths, measuring 18.43 mm, 17.70 mm, and 17.00 mm, respectively (Table 2).
Invasion and colonization of Fusarium isolates in muskmelon fruit at 10 dpi. (A) Fusarium pernambucanum human isolate FS10; (B) Fusarium pernambucanum veterinary isolate FS405; (C) Fusarium irregulare veterinary isolate FS097; (D) Fusarium irregulare veterinary isolate LD1; (E) Fusarium irregulare plant isolate PLMF1; and (F) Fusarium pernambucanum plant isolate BDMF7.
DISCUSSION
In this study, we identified FIESC as a pathogenic fungus capable of causing cross-infection in various hosts, including humans, animals, and plants. Notably, this pathogen was first reported in Taiwan in 2023, causing severe fruit rot in muskmelon fruit in different locations (30). Simultaneously, we successfully isolated the pathogen from human and veterinary sources. In humans, the pathogen was isolated from the sputum of a patient diagnosed with fungal pneumonia, who had a history of liver transplantation. Fusarium species have been reported to cause pneumonia, particularly in immunocompromised patients (10). Often manifesting as nodular and cavitary lesions, this condition is associated with increased mortality, even after controlling for immune status (31). Additionally, the pathogen has been isolated from a rabbit’s purulent nasal discharge, cat lung, and turtle. Similarly, Fusarium has been identified in the skin, nasal cavity, and lungs of Boer goats (13). In this study, the isolation of FIESC from diverse hosts and locations suggests that this fungus is widespread throughout Taiwan and capable of cross-infection across different species.
The commonly sequenced genes used for Fusarium identification include translation elongation factor-1α (TEF-1α), RNA polymerase 1 and 2 (RPB1 and RPB2), and β-tubulin (tub) (1). In this study, we used a combination of sequences of three genes, ITS, TEF 1-α, and RPB2, for identifying the FIESC isolates from plant, veterinary, and human sources. Based on the MLST results, all isolates belonging to the Incarnatum clade include two veterinary isolates, FS097 and LD1, and plant isolate PLMF1 which clustered with F. irregulare. Meanwhile, the veterinary isolate FS405, human isolate FS10, and plant isolate BDMF7 are clustered with F. pernambucanum. Previous multi-locus phylogenetic analyses identified 32 phylogenetic species within FIESC, which are divided into two major clades: the Equiseti clade and the Incarnatum clade (32, 33).
We examined the morphological characteristics of Fusarium isolates obtained from various host sources within the same species and incubated at 28°C, which has been reported as the optimal condition for observing FIESC isolate (30). Our findings revealed that differences among these isolates were observed in colony morphology and growth rate. Additionally, our results revealed that all F. irregulare and F. pernambucanum isolates from different hosts could produce an abundance of microconidia and chlamydospores on potato dextrose agar (PDA). However, in contrast to our findings, microconidia and chlamydospores were not observed in F. irregulare isolated from bamboo on PDA media (22). In F. pernambucanum isolate from humans, the predominant macroconidia exhibited 3 to 5 septa, whereas in plant isolate, most exhibited 3 septa, with a few displaying 4 or 5 septa. Both plant and human isolates demonstrated the ability to produce microconidia and chlamydospores on PDA. Additionally, Zhang et al. (34) reported that macroconidia of F. pernambucanum isolated from muskmelon had 3 to 5 septa on carnation leaf agar (CLA); however, no microconidia were observed. These morphological variations between our results and previous studies may be attributable to differences in cultural conditions. Various factors, including culture media, pH, and temperature, can significantly influence the mycelial growth and sporulation of Fusarium (35). Additionally, Fusarium isolates exhibit considerable variability in phenotypic characteristics, including colony color, pigmentation, sporulation, conidial size, and shape, even for the same species isolates (36). In this study, the morphological characteristics appeared to correlate more closely with host origin, which may indicate a degree of phenotypic plasticity. However, further investigation involving a larger number of isolates is necessary to validate these findings and assess the extent of host-driven morphological variation.
In this study, we examined the ability of Fusarium isolates obtained from various sources to penetrate and colonize the human nail plate, porcine skin, and muskmelon fruits. For all inoculation treatments, the spore suspension was standardized to a single concentration of 10⁶ conidia/mL, and SDW was used as a mock control due to the difficulty in identifying and sourcing a suitable negative control species. Notably, a previous study employed a single inoculum concentration to evaluate the pathogenicity of Fusarium species in plants and animals (30, 37). In the human nail infection test, results revealed that F. pernambucanum and F. irregulare isolates, regardless of their source, exhibited a remarkable ability to penetrate and colonize the nail plate within 4 days post-inoculation (dpi). This finding highlights their potential to cause onychomycosis in humans and underscores the adaptability of Fusarium across various environments and host types. Onychomycosis is the most common chronic fungal infection of the nail, accounting for approximately 50% of all nail diseases (38, 39). The most common fungi causing onychomycosis worldwide include Fusarium species, Aspergillus species, Scopulariopsis brevicaulis, Neoscytalidium dimidiatum, and Acremonium species (40).
Microscopic examination of Fusarium isolates colonized in the nail revealed that all isolates could produce chlamydospores at 4 dpi, indicating that the Fusarium isolates could establish an infection in a short time. However, no micro- or macroconidia were observed in infected nails. Previous studies reported the presence of chlamydospores and microconidia of Fusarium spp. throughout infected nails (41, 42) and in the reticular dermis, particularly associated with dermal blood vessels (43). The formation of chlamydospores in infected nails may aid in distinguishing dermatophytes from non-dermatophyte fungal causes of onychomycosis. Unlike previous studies, we observed no microconidia in infected nails. This discrepancy may be attributed to environmental factors or the specific Fusarium isolate examined.
Porcine skin was used as an ex vivo model to examine the ability of Fusarium isolates to infect human and animal skin. This model is frequently utilized due to its structural similarities to human skin, including thickness, hair follicle content, pigmentation, collagen, and lipid composition (44, 45). The results demonstrated that all Fusarium isolates could penetrate the dermal layer within 4 days post-inoculation. These findings suggest that FIESC isolates from various sources, including plant isolates PLMF1 and BDMF7, may cause both superficial and invasive diseases in humans and animals. This highlights Fusarium’s ability to transition from a plant pathogen to a zoonotic pathogen, underscoring its significance as a concern for ecosystems and public health. Similarly, Tava et al. (46) reported that strains of F. musae isolated from banana fruits and human patients can cause significant diseases in the banana as a plant host and Galleria mellonella larvae as a human proxy for the investigation of cross-kingdom pathogenicity. However, the severity of Fusarium infections in humans and animals is influenced by factors such as the Fusarium species and the host’s immune competence, with immunocompromised individuals being particularly susceptible to severe disease (31).
Beyond causing onychomycosis, Fusarium species can lead to invasive infections in humans, affecting the skin, lungs, blood vessels, and sinuses, often resulting in positive blood cultures (10). The skin is the most frequently affected organ in invasive infection, either as the primary site or through hematogenous spread, serving as an essential and early diagnostic clue for cutaneous lesions (10, 47). In addition to human infection, Fusarium has been reported to induce cutaneous, nasal, and systemic disease in a goat; the systemic infection was characterized by multifocal nodular lesions throughout the kidneys, abdominal lymph nodes, and lungs (13). Furthermore, Fusarium spp. have been associated with meningoencephalitis and chronic ulcerative dermatitis in dogs (11, 14).
Although all Fusarium isolates were capable of penetrating the dermal layer of porcine skin, our results indicated significant differences in penetration depth among isolates from various sources, including those within a single species. For instance, the human isolate FS10, which belongs to F. pernambucanum, exhibited a high penetration level, whereas the plant isolate BDMF7, also from the same species, demonstrated significantly lower penetration. These differences in penetration depth may reflect variations in pathogenicity. Our findings suggest that the pathogenicity of Fusarium isolates against porcine skin may depend more on their source than on their species. In contrast, a study examining the pathogenicity of Fusarium spp. isolates from clinical and environmental sources in a murine model found that virulence varied by species, with isolates belonging to F. solani being the most virulent (37). Additionally, virulence variations of F. musae strains in plant and animal hosts appear to depend more on the specific strain than on the host species (46). Based on our results and previous studies, the variance of Fusarium may be correlated to both the specific species and the individual isolate within that species. Furthermore, environmental conditions, such as temperature and humidity, can also influence the pathogenicity of Fusarium (48).
FIESC isolates from human, animal, and plant sources were evaluated for their pathogenicity against muskmelon fruits. The results indicated that all FIESC isolates were pathogenic to muskmelon, exhibiting different penetration depths at 10 days post-inoculation. FIESC has been reported to cause severe fruit rot disease in muskmelon in Taiwan (30). Furthermore, FIESC has been documented across various geographic regions as a causative agent of diseases in many crops, including tomato, eggplant, rockmelon, maize, soybean, rice, wheat, and barley (49–55). After inoculation with FIESC isolates from different sources, muskmelon fruits developed symptoms consistent with those observed on melon fruits under field conditions and as documented in previous reports (30, 56). Beyond its agricultural effects, FIESC poses a health risk to humans and animals. Transmission can occur through contaminated food, leading to either superficial or invasive infections. Additionally, these pathogens pose an indirect health hazard by producing a variety of mycotoxins (57). Currently, the pathogen has been reported in multiple regions, including Asia, Europe, and Africa, highlighting its global significance (58–60). Its wide distribution emphasizes the potential for cross-infection among plants, animals, and humans, as well as the associated risks of mycotoxin contamination.
Conclusion
In this study, we report that Fusarium isolates belonging to F. pernambucanum and F. irregulare obtained from various sources, including human, cat, rabbit, and turtle, along with plant samples from muskmelon fruits, represent newly identified pathogens in Taiwan. These isolates exhibit potential for cross-infection among plants, animals, and humans, highlighting their significance in both agricultural and medical contexts. Notably, our findings demonstrate the possibility of transmitting from a plant pathogen to a zoonotic pathogen of the plant isolates PLMF1 and BDMF7. To our knowledge, this is the first report of F. incarnatum cross-infection among humans, animals, and plants. The ability of this pathogen to adapt across hosts underscores the urgent need for comprehensive strategies to mitigate its impact on ecosystems and human health. In future research, we plan to investigate the mechanisms of cross-infection to better understand how fungal pathogens transition between biological systems. Additionally, it will be essential to evaluate their ability to produce mycotoxins within human and animal tissues to assess potential health risks.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Summerell BA. 2019. Resolving Fusarium: current status of the genus. Annu Rev Phytopathol 57:323–339. doi:10.1146/annurev-phyto-082718-10020431226019 · doi ↗ · pubmed ↗
- 2Gordon TR. 2017. Fusarium oxysporum and the Fusarium wilt syndrome. Annu Rev Phytopathol 55:23–39. doi:10.1146/annurev-phyto-080615-09591928489498 · doi ↗ · pubmed ↗
- 3Nelson P, Toussoun T, Cook R. 1981. Fusarium: diseases, biology, and taxonomy. Penn. State Univ. Press, University Park, PA.
- 4Anaissie EJ, Kuchar RT, Rex JH, Francesconi A, Kasai M, Müller FM, Lozano-Chiu M, Summerbell RC, Dignani MC, Chanock SJ, Walsh TJ. 2001. Fusariosis associated with pathogenic Fusarium species colonization of a hospital water system: a new paradigm for the epidemiology of opportunistic mold infections. Clin Infect Dis 33:1871–1878. doi:10.1086/32450111692299 · doi ↗ · pubmed ↗
- 5Dean R, Van Kan JAL, Pretorius ZA, Hammond-Kosack KE, Di Pietro A, Spanu PD, Rudd JJ, Dickman M, Kahmann R, Ellis J, Foster GD. 2012. The Top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol 13:414–430. doi:10.1111/j.1364-3703.2011.00783.x 22471698 PMC 6638784 · doi ↗ · pubmed ↗
- 6Sáenz V, Alvarez-Moreno C, Pape PL, Restrepo S, Guarro J, Ramírez AMC. 2020. A One Health perspective to recognize Fusarium as important in clinical practice. J Fungi (Basel) 6:235. doi:10.3390/jof 604023533092120 PMC 7711799 · doi ↗ · pubmed ↗
- 7Bansal Y, Singla N, Kaistha N, Sood S, Chander J. 2019. Molecular identification of Fusarium species complex isolated from clinical samples and its antifungal susceptibility patterns. Curr Med Mycol 5:43–49. doi:10.18502/cmm.5.4.2149 PMC 703478232104743 · doi ↗ · pubmed ↗
- 8Guarro J. 2013. Fusariosis, a complex infection caused by a high diversity of fungal species refractory to treatment. Eur J Clin Microbiol Infect Dis 32:1491–1500. doi:10.1007/s 10096-013-1924-723934595 · doi ↗ · pubmed ↗