Nasal delivery of Lacticaseibacillus rhamnosus GG modulates respiratory immune responses and attenuates Mycoplasma pneumoniae pneumonia in a murine model
Jiarong He, Wei Tian, Pengxiao Feng, Tingfeng Du, Huanbing Long, Guiting He, Hongjiang Wei, Xinyan Zhu, Xinyue Jiang, Cuiming Zhu

TL;DR
Intranasal administration of LGG reduces lung inflammation and bacterial load in a mouse model of Mycoplasma pneumoniae pneumonia.
Contribution
This study shows that intranasal LGG is safe and effective in both preventing and treating MPP in mice.
Findings
Intranasal LGG reduces lung inflammation and pathogen load in MPP-infected mice.
LGG promotes immune defenses through increased IgA, IgG, and anti-inflammatory cytokines.
LGG suppresses pro-inflammatory cytokines and neutrophil levels in the lungs.
Abstract
Mycoplasma pneumoniae pneumonia (MPP) management presents significant challenges due to the lack of an effective vaccine and the widespread occurrence of macrolide resistance. Recent studies investigating the use of oral or intranasal probiotics have shown promising results in reducing respiratory tract infections. In this study, BALB/c mice were initially given intranasal doses of Lacticaseibacillus rhamnosus GG (LGG), Limosilactobacillus reuteri F275, or a combination of these two strains, called MIX, to evaluate the safety of probiotic administration by examining lung tissue inflammation. Our results suggest that LGG is relatively safe, with minimal lung inflammatory responses, unlike the mild inflammation seen with L. reuteri F275 and MIX treatments. Next, LGG was administered to the mice via the respiratory tract on two consecutive days before or after infection with M. pneumoniae.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
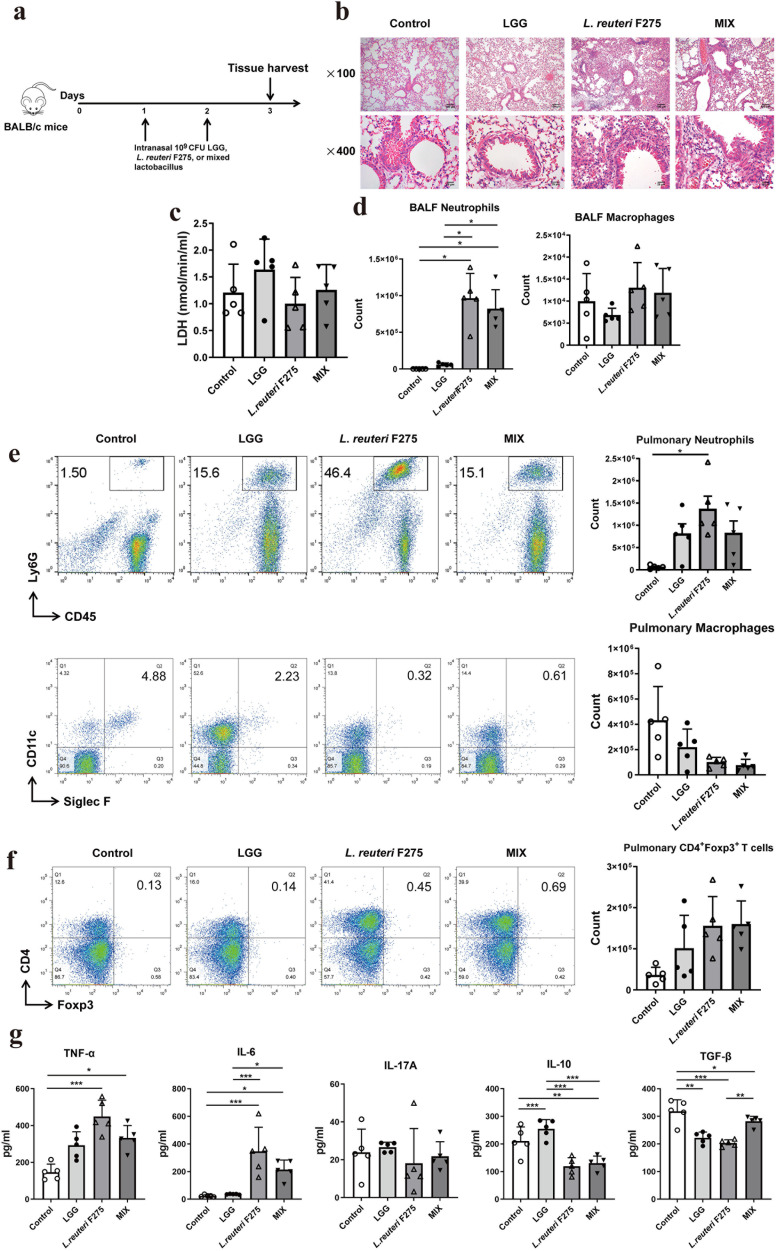 Fig 1
Fig 1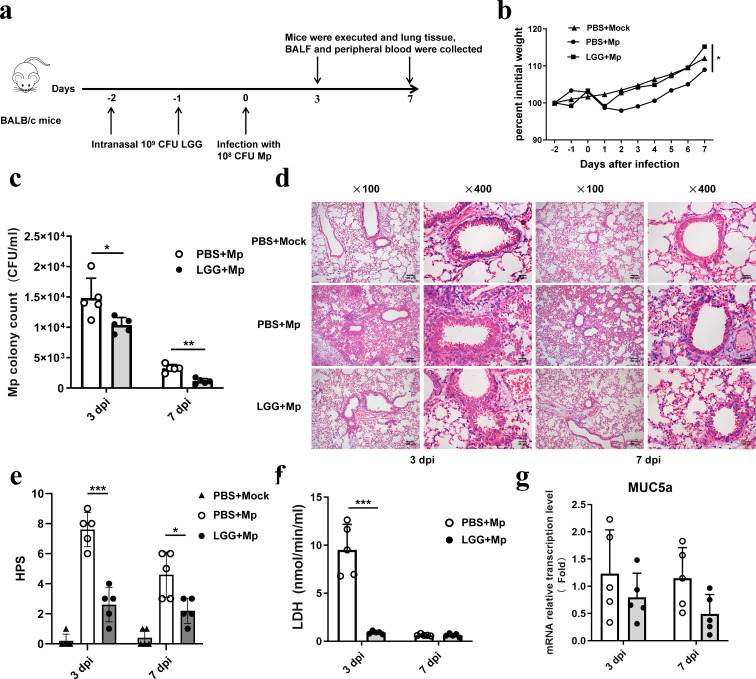 Fig 2
Fig 2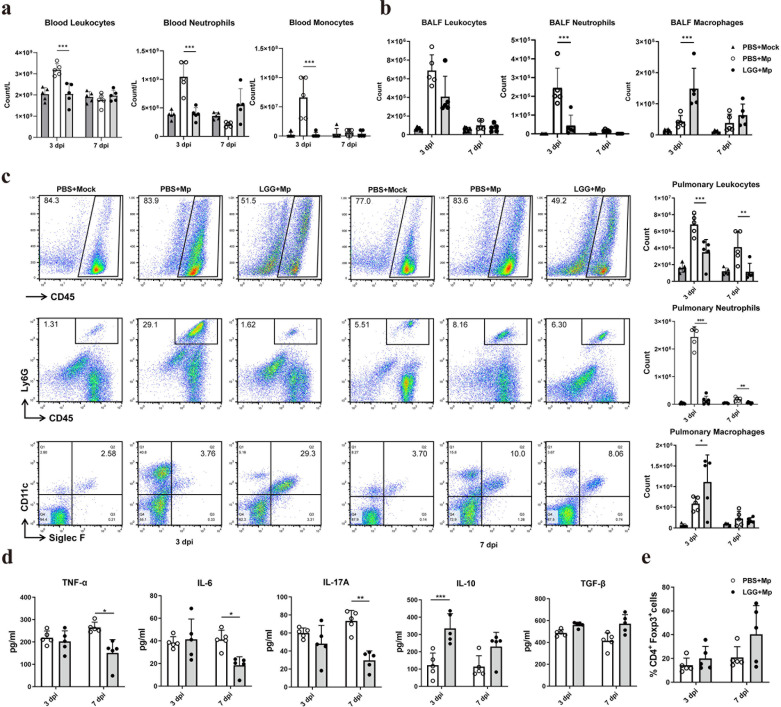 Fig 3
Fig 3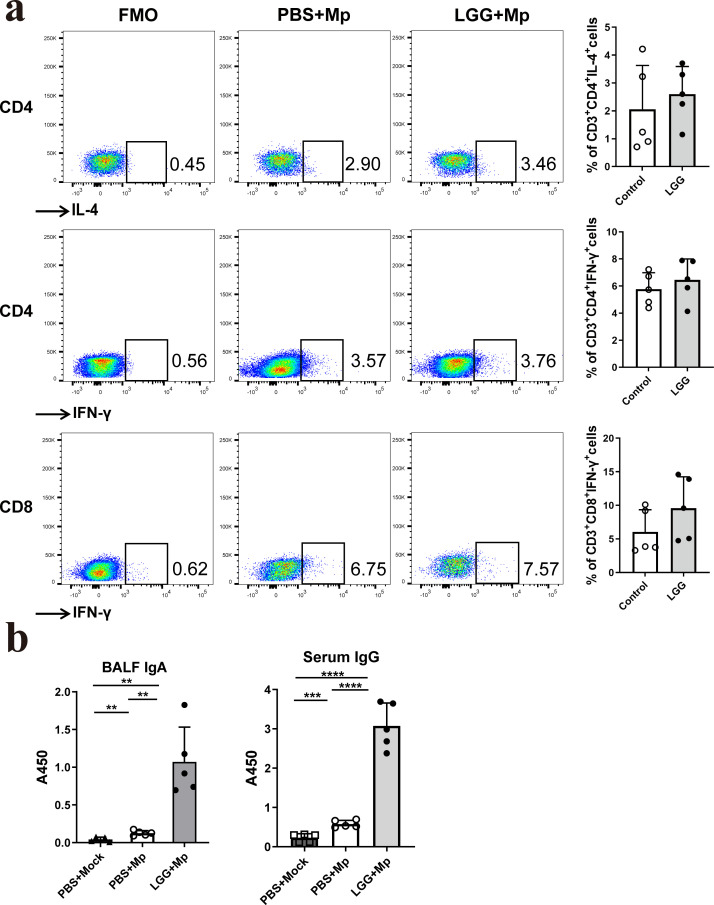 Fig 4
Fig 4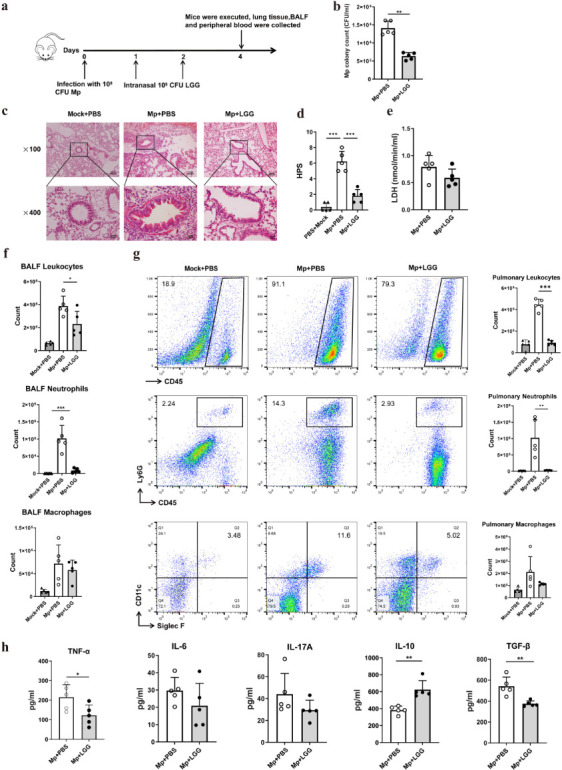 Fig 5
Fig 5- —Natural Science Foundation of Hunan Province
- —Research Project of Hunan Health Commission
- —Hunan Provincial College Students' Innovation and Entrepreneurship Training Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPneumonia and Respiratory Infections · Microbial infections and disease research · vaccines and immunoinformatics approaches
INTRODUCTION
Probiotics are “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” (1). Probiotic benefits are specific to certain strains and do not apply universally to the entire genus or all its species. Research indicates that many probiotic strains provide health benefits to humans. A well-known group with recognized probiotic potential is the genus Lactobacillus, which consists of gram-positive bacteria commonly found in various parts of the human body, including the gastrointestinal, oral, vaginal, and respiratory tracts. Some specific strains within this genus are widely used as probiotics in commercial products and as fermenting agents in dairy production.
Lacticaseibacillus rhamnosus GG (LGG) and Limosilactobacillus reuteri F275 (or Lactiplantibacillus plantarum NCIMB 8826) are common lactobacilli that are generally considered safe (2–4). The immunomodulatory properties of LGG and L. reuteri F275 have garnered significant attention due to their beneficial effects on the gastrointestinal mucosa (5, 6). Multiple clinical studies have demonstrated that taking LGG orally reduces the risk or decreases the duration of respiratory infections, including those induced by the H1N1 influenza virus and Pseudomonas aeruginosa (7–10). L. reuteri F275 has been demonstrated to enhance survival rates and reduce lung viral load and pulmonary inflammation in cases of pneumonia virus infection (11, 12). Nevertheless, certain studies have indicated that Lactobacillus species, whether alive or heat-inactivated, when administered to the respiratory tract, can elicit a more effective immunomodulatory response and mitigate the symptoms of respiratory pathogenic infections or allergic diseases (13–16). According to the research conducted by Percopo CM et al., the administration of either live or heat-inactivated L. reuteri F275 via intranasal delivery to mice before exposure effectively mitigated pulmonary inflammation, reduced viral load, and significantly increased survival rates in mice, resulting in a significant and sustained protective effect against subsequent lethal respiratory virus infections (12). The intranasal administration of LGG was found to upregulate the activity of lung natural killer (NK) cells and to stimulate the production of pro-inflammatory cytokines, including tumor necrosis factor α (TNF-α), interleukin (IL)−1β, and monocyte chemoattractant protein-1 (MCP-1), within the pulmonary tissues of BALB/c mice. This immunological response enhanced survival rates among adult mice infected with H1N1 (13). Similarly, neonatal C57BL/6 mice exhibited significantly improved survival rates and alleviated symptoms when administered intranasal LGG, leading to elevated levels of type I IFN (16). Notably, intranasal LGG administration demonstrated greater efficacy than oral supplementation in combating influenza virus infection, and live bacteria were more effective than inactivated bacteria (14, 17).
Mycoplasma pneumoniae constitutes a significant etiological agent in community-acquired pneumonia, particularly among school-aged children and young adults (17, 18). In recent years, the emergence of antibiotic resistance and the lack of efficacious vaccines have led to increasingly significant treatment challenges (19, 20). Our previous studies have shown that the oral administration of the probiotics Lacticaseibacillus casei CNRZ1874, LGG, and L. reuteri F275 can mitigate M. pneumoniae pneumonia (MPP) (21, 22). In this study, we demonstrated that in a murine model, pre- and post-intranasal inoculation with live LGG effectively reduced pathogen loads and mitigated respiratory infections following exposure to M. pneumoniae. The observed effects may be attributed to the immune regulatory properties of LGG, which decrease the expression of pro-inflammatory cytokines induced by the pathogen. Furthermore, LGG appears to promote the recruitment of alveolar macrophages and enhance the production of anti-inflammatory cytokine IL-10 and mycoplasma-specific immunoglobulin IgA (IgA) in bronchoalveolar lavage fluid (BALF) and IgG in serum.
MATERIALS AND METHODS
Mice
BALB/c mice, 6 weeks old, were obtained from Jiangsu Jicui Yaokang Biotechnology Co., Ltd. These animals were housed in a specific pathogen-free environment at the University of South China’s animal facility. Before initiating the experiment, the mice underwent a 7-day adaptation period in their housing environment.
Lactobacilli intranasal inoculations
The LGG (ATCC 53103) strain was generously provided by Professor Xiaohua Chen of Hengyang Normal University, China. L. reuteri F275 (ATCC23272, DSM20016, or CIP109823) was acquired from BeiNuo Biological Company, Shanghai, China. The LGG and L. reuteri F275 bacterial strains were cultivated in Man, Rogosa, and Sharpe (MRS) medium (Solarbio, China) at 37°C for 12–16 h. Subsequently, the cultures were centrifuged at 3,000 × g for 10 min. The supernatant was removed, and the bacterial pellet was rinsed once with sterile phosphate-buffered saline (PBS). The bacteria were then resuspended in sterile PBS to achieve a final concentration of 2.5 × 10^10^ colony-forming units (CFUs)/mL. After anesthetization with inhaled isoflurane, 10^9^ CFUs of LGG, 10^9^ CFUs of L. reuteri F275, or a combination of both (5 × 10^8^ CFUs of LGG and 5 × 10^8^ CFUs of L. reuteri F275) in 40 µL of PBS was intranasally inoculated into mice, with PBS delivery as a control. Respiration, weight loss, and locomotion were assessed daily.
M. pneumoniae preparation and infection
The standard strain of M. pneumoniae type 1, M129 (ATCC 29342), was maintained at the Institute of Pathogen Biology, Hengyang Medical College, University of South China. This strain was cultivated in a pleuropneumonia-like organism (PPLO) medium (BD Biosciences, USA). Cultivation occurred in a 225 cm^2^ cell culture flask at 37°C until the medium’s color changed from red to orange. Subsequently, the medium was carefully removed by gentle decantation, and the bottom of the vial was gently scraped with a cell scraper for several minutes. The sample was centrifuged for 15 min at 10,000 × g to harvest mycoplasma cells, and the cells were washed once with sterile PBS and then resuspended in the same medium. The CFUs of M. pneumoniae were assessed using the following protocol: After freezing for several days, a tube of mycoplasma was retrieved from the freezer and subjected to a single rapid freeze-thaw cycle. CFUs were then determined by plating serial dilutions of the organism onto CM401 solid medium (Oxoid) and counting colonies under 100× magnification after approximately 7 days of incubation. Additional M. pneumoniae aliquots stored at −80°C were used for immunological testing or challenge experiments, all of which were performed within 1 month of storage. All M. pneumoniae-infected mice received intranasal inoculation with 10^8^ CFUs of this organism in 40 µL of PBS under isoflurane anesthesia. Mice were euthanized at 3 and 7 days post-infection (dpi) via carbon dioxide inhalation. The 3 dpi time point was selected to assess LGG’s protective efficacy against an acute M. pneumoniae challenge and its modulation of innate immunity, specifically neutrophil and macrophage activation (23, 24). The 7 dpi time point was chosen to evaluate the adaptive immune response, characterized by significant increases in pathogen-specific IgM antibodies (22, 24) and markedly elevated lung T-cell counts, as previously demonstrated. Cardiac blood, BALF, and lung samples were collected for further analysis.
The number of M. pneumoniae colonies
The collected BALF was centrifuged at 900 × g for 10 min. Subsequently, 50 µL of the resulting supernatant was uniformly distributed on a solid PPLO medium. The prepared medium was then placed in a temperature-controlled incubator maintained at 37℃, with an atmosphere of 95% N2 and 5% CO2, for 7–10 days. The solid plate was then placed under a standard inverted microscope for observation, and the number of mycoplasma colonies was quantified under low magnification.
Histopathology scoring
The right lower lobe of the lung was fixed in a 4% paraformaldehyde solution (Biosharp, China) for 48 h. Subsequently, paraffin embedding, sectioning, and hematoxylin and eosin staining were performed, and pathological inflammatory damage to the lung tissue was observed utilizing a double-blind method. Histopathology scoring (HPS) was assessed by analyzing the quantity and infiltration area of inflammatory cells in the alveoli and bronchi, the extent of proliferation of blood vessels and bronchial tube walls, and the delineation of borders (25).
Lactate dehydrogenase production
Following the manufacturer’s protocol, lactate dehydrogenase (LDH) production in the BALF supernatant, an indicator of lung tissue cytotoxicity, was quantified using an LDH assay kit (Solarbio, China).
Cell sorting and counting
200 µL of cardiac blood was transferred to an EDTA-K2 anticoagulant tube and gently inverted to ensure thorough mixing. Cell analysis was performed using an XN-1000V Animal Hematology Analyzer (Xisenmecan).
Cells in BALF were isolated by centrifugation at 900 × g for 10 min, after which they were resuspended in PBS for subsequent analysis. Lung tissue specimens were excised and homogenized in 1 mL of sterile PBS. The homogenate was filtered through a 40-micron cell strainer and then centrifuged at 1,200 × g for 5 min. The resultant pellet was resuspended in a 35% PERCOLL solution and subjected to density gradient centrifugation at 700 × g for 15 min. The upper liquid and impurities were carefully removed. Subsequently, ACK lysis buffer was applied to the sediment to lyse erythrocytes at room temperature for 5 min. Finally, pulmonary cells were harvested by centrifugation at 1200 × g for 5 min (26). The cells from BALF and pulmonary tissue were then stained with anti-mouse CD45-FITC, anti-mouse Ly-6G-PE, anti-mouse CD11c-PerCP-Cy5.5, and anti-mouse Siglec-F-A647(APC) antibodies. Following immunostaining, cells were analyzed using flow cytometry. Specifically, CD45+ cells were identified as leukocytes, CD45+Ly6G+ cells as neutrophils, and CD45+CD11c+Siglec-F+ cells as alveolar macrophages. Absolute count of pulmonary neutrophils or macrophages = total cells in the sample × percentage of target cells (%).
Cytokines in BALF
Under the manufacturer’s protocol, an indirect enzyme-linked immunosorbent assay (ELISA) was conducted to quantify the cytokines TNF-α, IL-6, IL-10, IL-17A, and transforming growth factor-beta (TGF-β) in the supernatant obtained from BALF. The absorbance was measured at A450, and the concentrations of the cytokines were calculated based on the established standard curve.
Reverse transcription-quantitative PCR
The total RNA from pulmonary cells was isolated using TRIzol reagent (TIANGEN, China) and subsequently converted to cDNA using FastKing One-Step Genomic cDNA First-Strand Synthesis Premix Reagent (KR118) (TIANGEN, China). Subsequently, the Muc5a gene was detected through quantitative reverse transcription PCR (qRT-PCR) utilizing a SYBR Green Premix Ex Taq kit (Bio-Rad, 1725121). The analysis was conducted using the 2^−ΔΔCt^ method, with β-actin as the internal control. The sequences of the primers are presented as follows: Muc5a forward, 5′-CAGGACTCTCTGAAATCGTACCA-3′; Muc5a reverse, 5′-AAGGCTCGTACCACAGGGA-3′; β-actin forward, 5′-TGCTGTCCCTGTATGCCTCT-3′; β-actin reverse, 5′-AGGTCTTTACGGATGTCAACG-3.'
Intracellular cytokines and Foxp3
Pulmonary cells were stimulated with 50 ng/mL PMA, 1 µg/mL ionomycin, and 1 µmol/mL brefeldin A for 4 h. After which, the cells were blocked with rat anti-mouse Fc receptor (CD16/CD32) antibody (BD Biosciences), and then incubated with anti-mouse CD3e-PE-Cy7 (BD PharMingen, USA), anti-mouse CD4-PE (BioLegend, USA), and anti-mouse CD8a-FITC (BioLegend) for 30 min at 4°C for surface molecules immunostaining. Subsequently, dead cells were distinguished by staining with the anti-mouse dye eflour@560. Following this, the cells were fixed and permeabilized using a buffer set (BD Biosciences) and subsequently incubated with anti-mouse IFN-γ-APC and anti-mouse IL-4-PerCP-Cy5.5 (BioLegend, USA) for 30 min. For Treg analysis, pulmonary cells were stained with rat anti-mouse Fc receptor (CD16/CD32) antibody and anti-mouse CD4-PE. After fixation and permeabilization, cells were incubated with anti-mouse Foxp3-Alexa Fluor (APC) (BD PharMingen, USA) for 30 min. After staining, the cells were collected by centrifugation at 12,000 × g for 5 min and resuspended in FACS buffer. Finally, the samples were examined using an LSRII flow cytometer (BD Biosciences), and the resulting data were analyzed using FlowJo software. The cells underwent two washes at each procedural step using a fluorescence-activated cell sorting (FACS) buffer; however, sterile PBS was used for washing before staining the dead cells with the anti-mouse dye eFlour@560.
Antibodies in serum and BALF
To prepare the antigen, M. pneumoniae was collected and centrifuged at 10,000 × g for 15 min. Next, the cells were washed with sterile PBS before being harvested by centrifugation. The resulting pellet was suspended in 10 mL of PBS and disrupted using ultrasonic waves. The sonication process consisted of 3-second bursts followed by 10-second intervals, continuing for 1 h. The protein content of the samples was determined by a BCA kit (Epizyme, China).
ELISA was used to detect the production of M. pneumoniae-specific serum IgG and BALF IgA. In summary, M. pneumoniae antigens were suspended in a carbonate coating buffer at a concentration of 50 mmol/L, with a pH of 9.6, to achieve a final concentration of 15 µg/mL. The antigen solution was subsequently applied to 96-well microtiter plates and incubated overnight at 4°C. Following incubation, the plates were thoroughly washed with buffer and blocked with 200 µL of 5% skim milk powder for 2 h at 37°C. After an additional washing step, 100 µL of diluted serum or BALF supernatant was added to the designated test wells, and the plates were incubated for a further 2 h at 37°C. The plates were rewashed, after which horseradish peroxidase-conjugated goat anti-mouse IgG or IgA antibodies (Proteintech, USA) were added at dilutions of 100 µL per well. The plates were incubated for 1 h at 37°C. Finally, 100 µL of TMB solution was added to each test well, and the plates were incubated in the dark at 37°C for 15 min. The chromogenic reaction was halted with a stop solution, and the optical density was measured using an ELISA instrument (PerkinElmer, USA) to quantify IgG and IgA levels.
Statistical analyses
Statistical analyses were performed using GraphPad Prism 8 software (GraphPad Software, Inc., La Jolla, CA, USA). Results are presented as mean ± standard deviation (SD). Differences were assessed using a two-way ANOVA with the Sidak multiple comparison test or a Student’s t-test. Statistical significance was defined as P < 0.05.
RESULTS
Impact of nasal delivery of Lactobacillus on mouse pulmonary tissue
To evaluate the safety of intranasal Lactobacillus administration, alterations in the lung tissues of mice were examined. The experimental procedure was illustrated in Fig. 1a. Although the alveolar structure remained relatively intact, the number of inflammatory cells and area of infiltration of inflammatory cells into the alveoli of mice intranasally administered live L. reuteri F275 and MIX increased (Fig. 1b). Although no significant differences were observed in LDH production in BALF or the population of regulatory T cells (Tregs) in the lungs of mice inoculated intranasally with PBS, LGG, L. reuteri F275, and MIX (Fig. 1c and g), the MIX formulation and PBS control, nasal administration of L. reuteri F275, and the MIX formulation significantly increased neutrophil counts within pulmonary tissue and elevated levels of TNF-α and IL-6 in BALF in the murine model (Fig. 1b through g). At the same time, a reduction in IL-10 and TGF-β production was observed (Fig. 1g). Conversely, LGG administration enhanced IL-10 secretion without affecting neutrophil influx or the levels of TNF-α, IL-6, and TGF-β (Fig. 1g).
Effects of nasal administration of Lactobacillus in mice lungs. On the first day following Lactobacillus supplementation, mice were sacrificed, and the lungs and BALF were collected for further analysis. (a) Experimental procedure for intranasal Lactobacillus supplementation in mice. (b) Lung histopathology (representative images at ×100 and ×400 magnification for each experimental group). (c) LDH content in BALF. (d and e) Neutrophils and alveolar macrophages were detected and quantified in both lung tissue and BALF using flow cytometry. (f) CD4+Foxp3+ cells in the mouse lung tissue were analyzed by flow cytometry. (g) Secretion of pro-inflammatory cytokines in the BALF was quantified using ELISA. N = 20, n = 5. Each value indicates mean ± SD. * P < 0.05, ** P < 0.01, *** P < 0.001.
Pre-intranasal administration of LGG alleviated MPP
The mycoplasma cell load and inflammatory pathology in the lungs of mice were determined to evaluate the effect of the pre-intranasal administration of LGG on M. pneumoniae infection. The experimental design is illustrated in Fig. 2a. Intranasal administration of LGG before the infection with M. pneumoniae did not yield a statistically significant difference in the weight of the mice when compared to the control group (Fig. 2b). Three days after infection, pre-treatment with LGG substantially lowered the pathogen burden (Fig. 2c). Additionally, a reduced presence of inflammatory cells, smaller areas of infiltration, and more intact alveolar structures were observed in the lungs of LGG-administered mice, along with a significant decrease in HPS and LDH compared to the PBS pre-treated control group (Fig. 2d through f), providing further evidence for the mitigation of lung tissue damage. MUC5a is upregulated in response to inflammatory conditions, and it has been reported that excessive production of MUC5a is a major contributor to airway obstruction. It was reported that probiotic and prebiotic therapy could significantly reduce the transcription of the MUC5a gene in asthmatic mice (27, 28); however, our study showed that the administration of LGG did not alter the transcription levels of MuC5a in the lungs (Fig. 2g). Seven days post-infection, the LGG-pretreated group demonstrated a reduction in M. pneumoniae burden compared to the control group, whereas no significant differences were observed between the two groups regarding pulmonary histopathology, inflammatory cell counts, HPS, and MUC5a expression (Fig. 2c through g).
LGG pre-treatment reduces lung tissue damage caused by M. pneumoniae infection. Mice were nasally pre-administered with LGG and then infected with M. pneumoniae via the respiratory tract. The animals were euthanized at 3 and 7 dpi. (a) Experimental procedure for nasal pretreatment with Lactobacillus before M. pneumoniae infection, (b) Body weight, (c) M. pneumoniae colony counts in mouse BALF, (d) lung histopathology (representative images at ×100 and ×400 magnification for each experimental group), (e) HPS, (f) LDH content in BALF, and (g) mRNA relative transcription of MUC5a in mouse lungs, n = 5. Each value shows the mean ± SD. * P < 0.05, ** P < 0.01, *** P < 0.001.
Pre-treatment with LGG altered innate cellular profiles and cytokine responses
To investigate the effects of intranasal LGG on the modulation of innate immunity in the context of mitigating MPP, this study analyzed the classification and enumeration of cells in the peripheral blood, BALF, and lung tissue, as well as the expression of cytokines in BALF of mice infected with M. pneumoniae. Three days after infection with M. pneumoniae, the intranasal administration of LGG led to a significant decrease in the total leukocyte count, along with reductions in the numbers of neutrophils and monocytes in the peripheral blood (Fig. 3a). In addition, there was a decrease in leukocytes and neutrophils and an increase in alveolar macrophages in the lung tissue of mice (Fig. 3a). In the BALF, the changes in leukocytes, neutrophils, and alveolar macrophages mirrored those in the lungs (Fig. 3b). Seven days after M. pneumoniae infection, mice that received LGG pre-treatment showed a significant decrease in lung leukocytes and neutrophils, as well as neutrophils in BALF, compared to those pretreated with PBS (Fig. 3c). On day 3 post-infection, nasal administration of LGG before M. pneumoniae infection increased the anti-inflammatory cytokine IL-10. At the same time, there were no changes in the levels of TNF-α, IL-6, and IL-17 (Fig. 3d). However, on day 7 post-infection, levels of the pro-inflammatory cytokines TNF-α, IL-6, and IL-17A significantly decreased (Fig. 3d). No significant differences were observed in TGF-β levels or the percentage of Tregs between the two experimental groups on days 3 and 7 post-infection (Fig. 3e).
Intranasal pre-treatment with LGG altered innate cellular profiles and cytokine responses following M. pneumoniae infection. Mice were pre-administered LGG nasally and then subjected to M. pneumoniae infection via the respiratory tract. The animals were euthanized at 3 and 7 dpi. (a–c) Leukocytes, neutrophils, and monocytes (or alveolar macrophages) were analyzed and quantified in peripheral blood, BALF, and pulmonary cells by flow cytometry. (d) Cytokine concentrations in BALF were determined using ELISA. (e) The percentage of pulmonary Tregs was assessed by flow cytometry. N = 30, n = 5. Each value represents the mean ± SD. * P < 0.05, ** P < 0.01, *** P < 0.001.
Pre-treatment with LGG regulated the adaptive immunity triggered by M. pneumoniae
This study examined the impact of nasal pre-administration of LGG on the adaptive immune response in mice infected with M. pneumoniae. To gain insights into these effects, the activation of T cells and the production of specific antibodies were analyzed. The results of our study indicated that the pre-administration of LGG did not significantly alter the profiles of specific immune cells in the pulmonary tissue of infected mice. This includes immune cell populations such as CD3+CD4+IL-4+, CD3+CD4+IFN-γ+, and CD8+IFN-γ+ cells (Fig. 4a). Nevertheless, LGG resulted in a significant increase in pathogen-specific IgA levels in the BALF and enhanced IgG levels in the serum (Fig. 4b). These findings demonstrate that LGG effectively modulates the humoral immune response.
*Intranasal pretreatment with LGG regulated adaptive immunity triggered by M. pneumoniae. Mice were pre-administered LGG nasally and then subjected to M. pneumoniae infection via the respiratory tract. The animals were euthanized at 7 dpi. (a) The percentage of T cells was analyzed by flow cytometry, and (b) the levels of M. pneumoniae-specific IgA in BALF and IgG in serum were analyzed using ELISA. N = 15, n = 5. Each value represents the mean ± SD. **P < 0.01, *** P < 0.001, ***P < 0.0001.
Post-treatment with LGG enhanced the resistance to M. pneumoniae
To evaluate the potential of LGG as an adjunct therapy for MPP, intranasal administration was performed following M. pneumoniae infection. The experimental protocol is illustrated in Fig. 5a. Intranasal LGG treatment after M. pneumoniae infection significantly decreased pathogen colonies compared to the control group (Fig. 5b). Participants who received LGG exhibited reduced inflammatory infiltration in the peribronchial and perivascular regions, along with a decrease in HPS (Fig. 5c and d), while there was no effect on LDH levels (Fig. 5e). Additionally, LGG administration markedly lowered total leukocyte and neutrophil counts in lung tissue and BALF; however, alveolar macrophage populations remained unchanged (Fig. 5f and g). Cytokine analysis revealed that LGG therapy decreased TNF-α and TGF-β secretion and increased IL-10 production in BALF (Fig. 5h).
*Post-treatment with LGG enhanced resistance to M. pneumoniae. 24 h following intranasal infection with M. pneumoniae, the mice were nasally treated with LGG for two consecutive days and subsequently sacrificed. (a) Experimental procedure for the LGG treatment, (b) the M. pneumoniae colony counts in mouse BALF, (c) lung histopathology (representative images at ×100 and ×400 magnification for each experimental group), (d) HPS, (e) LDH content in BALF, (f and g) Flow cytometry detection of leukocytes, neutrophils, and alveolar macrophages in BALF and lung tissue, (h) Cytokines secretion in the BALF quantified using ELISA. N = 15, n = 5. Each value represents the mean ± SD. *P < 0.05, **P < 0.01, **P < 0.001.
DISCUSSION
Although LGG and L. reuteri F275 are generally considered safe intestinal probiotics in the gastrointestinal tract, their non-native status in the respiratory tract warrants examination of histopathological changes following intranasal administration. Previous research has evidenced that 24 hours following intranasal administration, L. reuteri F275 is no longer detectable in lung tissue and is associated with temporary, mild, heterogeneous inflammation (11). Our results revealed that intranasal administration of 10^9^ CFUs of L. reuteri F275 or MIX resulted in an elevated presence of neutrophils in lung tissue and the upregulation of pro-inflammatory cytokines TNF-α and IL-6 and led to minor pulmonary damage, which aligns with findings from previous research (11). In contrast to the effects observed in the lungs of mice treated with L. reuteri F275, our study indicates that intranasal administration of LGG is a relatively safe inoculation method. The treatment did not significantly alter the number of immune cells, cytokine release, or LDH production in the lung tissues of BALB/c mice. Furthermore, it did not cause any inflammatory pathological damage to the lung tissue. These findings are consistent with prior studies in this field (11, 12, 14, 16).
M. pneumoniae infection is principally responsible for developing bronchitis, tracheitis, and pneumonia (18). Although the pathogenesis of M. pneumoniae infection remains to be elucidated, neutrophil recruitment and the induction of pro-inflammatory and other cytokines under both acute and chronic conditions are essential components of this infection (18, 29). Accordingly, the primary objective in preventing MMP is to diminish the burden on the organism and the ensuing inflammatory response. The results of our study demonstrated that intranasal administration, either before or after infection, effectively reduced the number of mycoplasma cells and mitigated MPP in a murine model. These effects were comparable to those observed when LGG was administered orally, whether in its viable form or after heat inactivation (22).
The clearance of pathogens following M. pneumoniae infection primarily depends on alveolar macrophage recruitment and activation, as murine models with elevated alveolar macrophage populations are essential for resistance to this pathogen (29–31). Conversely, rather than facilitating M. pneumoniae clearance, excessive neutrophil presence can result in severe tissue damage (29, 32, 33), and the suppression of neutrophil itaconate production can mitigate MPP (34). Furthermore, in the development and progression of MPP, particularly in refractory cases, the equilibrium between Th17 and Treg cells plays a crucial role (35). IL-17 plays a pivotal role in M. pneumoniae infections and is a significant predictor of refractory MPP in pediatric patients (36). The abnormal secretion of IL-17 in pulmonary tissue can trigger the activation of epithelial cells to produce chemokines, leading to ongoing neutrophil influx, tissue damage, and potentially asthma (37–39). In contrast to Th17 cells, Tregs are generally recognized as potent suppressors of mycoplasma respiratory infections, and Tregs and IL-10 are involved in suppressing IL-17A production (40). During the acute phase of MPP, Treg functionality is notably compromised. Curiously, as the condition progresses to the resolution stage, the impairment of Treg function coincides with an elevation in Th17 cell numbers (41). The Th17/Treg ratio strongly correlates with the severity of MPP and plays a significant role in the pathogenesis of refractory MPP (40, 41).
LGG modulates innate immunity through TLR interactions, the induction of antimicrobial peptides, and the strengthening of the epithelial barrier. It influences the maturation of dendritic cells, the polarization of macrophages, and the function of neutrophils, while promoting anti-inflammatory effects. Competitive exclusion and immunomodulatory components further reduce bacterial load (42). The investigation demonstrated that mice pretreated with LGG before M. pneumoniae infection exhibited an increased population of alveolar macrophages, suggesting enhanced clearance of the pathogen. Furthermore, LGG pretreatment resulted in reduced leukocyte and neutrophil counts in both peripheral blood and lung tissue, along with lower levels of the pro-inflammatory cytokines IL-17A, TNF-α, and IL-6. Concurrently, IL-10 levels were elevated in the lungs, indicating reduced pulmonary inflammation. Although LGG pretreatment could reduce M. pneumoniae load in the lungs of mice, the decrease was not substantial, as the bacterial count in the lungs did not change by even a logarithmic unit. One reason might be that, although LGG administration can activate immune cells such as macrophages, this activation was not strong enough to produce a robust immune response that clears the pathogen. Additionally, on day 3 post-infection, the levels of pro-inflammatory cytokines in BALF were similar between mice pretreated with LGG and those with PBS, indicating an inadequate immune response to suppress and eliminate the pathogen. Furthermore, in the experiment, the interval between LGG administration and infection was short (only 2 days). LGG may not have had sufficient time to multiply and exert its immunomodulatory effects before the pathogen attack, thus failing to reduce the pathogen load significantly.
Although LGG treatment post-infection did not significantly affect alveolar macrophage populations, it decreased the number of mycoplasma colonies. This outcome may be attributed to reduced neutrophil infiltration, decreased TNF-α production, and increased IL-10 expression in the lungs. Earlier studies have shown that the oral administration of LGG results in an increase in lung Tregs in mice infected with P. aeruginosa (8); however, the present study did not observe an increase in Tregs or the expression of TGF-β following intranasal administration of LGG in mice infected with M. pneumoniae. Interestingly, administering LGG to mice, whether alone or following an infection with M. pneumoniae, resulted in a significant reduction in TGF-β secretion in the BALF. To build on these findings, further research is required to investigate the expression of the CD25 molecule in Tregs. Moreover, the processes responsible for producing Tregs and TGF-β remain unclear and require further investigation.
Undoubtedly, the course and development of M. pneumoniae infection are significantly modulated by adaptive immune responses. In addition to their critical role in controlling or preventing MPP, antibodies and effector T cells also inhibit the dissemination of mycoplasma infections from mucosal regions to other bodily tissues, which can potentially result in arthritis and other diseases. Earlier investigations have suggested that IgA may play a crucial role in suppressing M. pneumoniae’s ability to adhere during infection. IgG is vital in the body’s immune defense mechanism, particularly in the later phases of M. pneumoniae infections (25, 43). Our research revealed that intranasal pre-administration of LGG did not alter the number of pulmonary CD4+IL-4+ T cells, CD4+IFN-γ+ T cells, or C8+IFN-γ+ T cells in M. pneumoniae-infected mice. Nonetheless, our results showed that intranasal administration of LGG resulted in a significant elevation of IgA concentrations in the BALF and IgG levels in the serum. This effect may be linked to the increased number of alveolar macrophages, which act as antigen-presenting cells, enhancing the ingestion, processing, and presentation of M. pneumoniae antigens and subsequently activating T cells to stimulate humoral immunity. The roles of M. pneumoniae adaptive immunity present a dichotomy. Despite immune responses being advantageous in preventing infection, they often fail to eliminate mycoplasma and disease, resulting in the development of chronic inflammation (25, 43, 44). Furthermore, animal studies have demonstrated that the severity of the disease is exacerbated by the antibody response (45). Consequently, additional research is necessary to elucidate the impact of the heightened antibody levels induced by intranasal pre-administration of LGG.
To summarize, our findings suggest that intranasal administration of LGG is a promising alternative approach for preventing and managing MPP. Intranasal administration of LGG for two consecutive days before M. pneumoniae infection or 24 hours post-pathogen exposure significantly reduced the mycoplasma cell burden and ameliorated lung tissue damage in a murine model. The modulation of intranasal LGG may be responsible for this outcome, as it influences the innate and adaptive immune responses within the respiratory system when triggered by M. pneumoniae. These responses include increased alveolar macrophage counts, IL-10 production, and specific mycoplasma BALF IgA and serum IgG levels, concurrent with reduced neutrophil infiltration and decreased production of pro-inflammatory cytokines, such as TNF-α, IL-6, and IL-17A. However, additional research is necessary to fully understand the effects of intranasal LGG administration on M. pneumoniae infection, including studies with larger sample sizes, diverse animal models, and investigations into the long-term protective effects.
Conclusion
Our results demonstrated that both intranasal pre- and post-administration of LGG significantly ameliorated the inflammatory pathology of lung tissue induced by M. infection and reduced the pathogen burden. The protective effects were attributed to the immunomodulatory response of nasal LGG, which involved the recruitment of leukocytes, the release of cytokines, and the production of specific antibodies. Intranasal administration with LGG is a potentially productive and safe intervention for mitigating MPP.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Borchers AT, Selmi C, Meyers FJ, Keen CL, Gershwin ME. 2009. Probiotics and immunity. J Gastroenterol 44:26–46. doi:10.1007/s 00535-008-2296-019159071 · doi ↗ · pubmed ↗
- 2O’Callaghan J, O’Toole PW. 2013. Lactobacillus: host-microbe relationships. Curr Top Microbiol Immunol 358:119–154. doi:10.1007/82_2011_18722102141 · doi ↗ · pubmed ↗
- 3Heeney DD, Gareau MG, Marco ML. 2018. Intestinal Lactobacillus in health and disease, a driver or just along for the ride? Curr Opin Biotechnol 49:140–147. doi:10.1016/j.copbio.2017.08.00428866243 PMC 5808898 · doi ↗ · pubmed ↗
- 4Du T, Lei A, Zhang N, Zhu C. 2022. The beneficial role of probiotic Lactobacillus in respiratory diseases. Front Immunol 13:908010. doi:10.3389/fimmu.2022.90801035711436 PMC 9194447 · doi ↗ · pubmed ↗
- 5Vaarala O. 2003. Immunological effects of probiotics with special reference to lactobacilli. Clin Exp Allergy 33:1634–1640. doi:10.1111/j.1365-2222.2003.01835.x 14656348 · doi ↗ · pubmed ↗
- 6Kalyuzhin OV, Afanasyev SS, Bykov AS. 2016. Probiotics as stimulators of immune response against pathogens in the respiratory tract. Ter Arkh 88:118–124. doi:10.17116/terarkh 2016885118-12427458629 · doi ↗ · pubmed ↗
- 7Kawase M, He F, Kubota A, Harata G, Hiramatsu M. 2010. Oral administration of lactobacilli from human intestinal tract protects mice against influenza virus infection. Lett Appl Microbiol 51:6–10. doi:10.1111/j.1472-765X.2010.02849.x 20438618 · doi ↗ · pubmed ↗
- 8Khailova L, Baird CH, Rush AA, Mc Namee EN, Wischmeyer PE. 2013. Lactobacillus rhamnosus GG improves outcome in experimental Pseudomonas aeruginosa pneumonia: potential role of regulatory T cells. Shock 40:496–503. doi:10.1097/SHK.000000000000006624240593 PMC 5592098 · doi ↗ · pubmed ↗