Novel lytic phages improve the antibiofilm activity of dalbavancin, daptomycin, and fosfomycin against vancomycin-resistant enterococci
Rima Fanaei Pirlar, Alvaro Irigoyen-von-Sierakowski, Jeroen Wagemans, Yu Ning, Rob Lavigne, Andrej Trampuz, Svetlana Karbysheva

TL;DR
Two new phages work with antibiotics to effectively kill biofilms of drug-resistant enterococci, offering a potential treatment for difficult joint infections.
Contribution
The study introduces two novel lytic phages that synergize with antibiotics to combat vancomycin-resistant enterococci biofilms.
Findings
Phage-antibiotic combinations, especially with dalbavancin and fosfomycin, significantly enhanced antibiofilm activity against VRE.
CUB-FM and CUB-FS phages showed dose-dependent antibiofilm effects and were effective at specific concentrations.
Genomic analysis confirmed the phages are safe, lacking virulence or resistance genes.
Abstract
Periprosthetic joint infections (PJI) caused by Enterococcus spp., especially vancomycin-resistant strains (VRE), are challenging to treat due to biofilm tolerance and limited antibiotic options. Bacteriophages offer a promising adjunct through targeted and biofilm-disrupting activity. This study evaluated two novel lytic phages, alone and combined with last-line antibiotics, for their ability to eradicate VRE biofilms in vitro. Two novel lytic phages, CUB-FM (E. faecium) and CUB-FS (E. faecalis), were isolated from hospital sewage and characterized via whole-genome sequencing and transmission electron microscopy. Antibiofilm efficacy of phages alone and in combination with dalbavancin, daptomycin, and fosfomycin was assessed against biofilm-embedded VRE strains using isothermal microcalorimetry. Synergy was defined as a combined effect exceeding the sum of individual activities.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
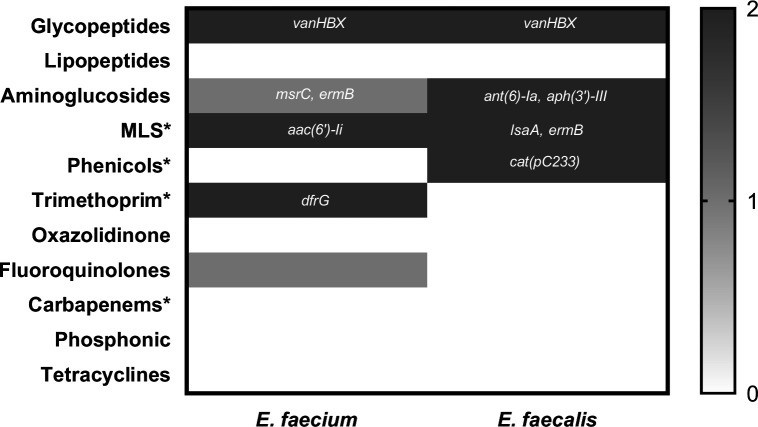 Fig 1
Fig 1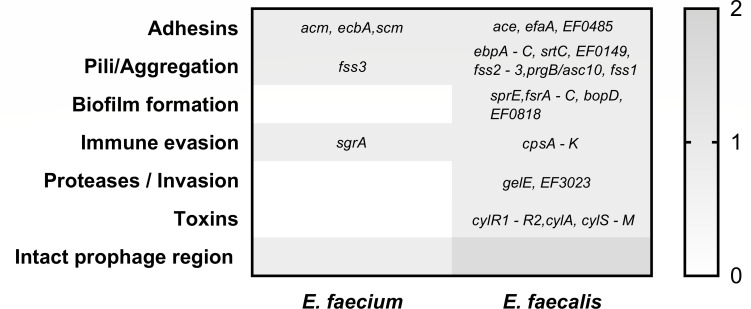 Fig 2
Fig 2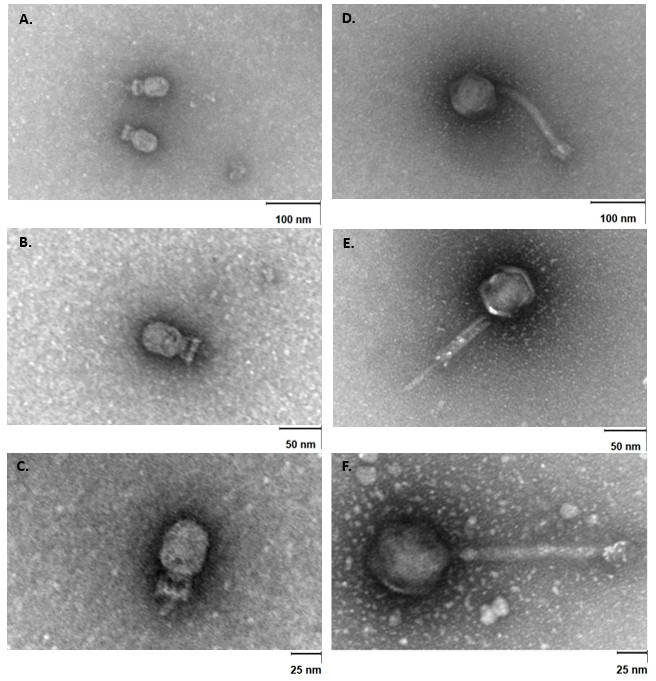 Fig 3
Fig 3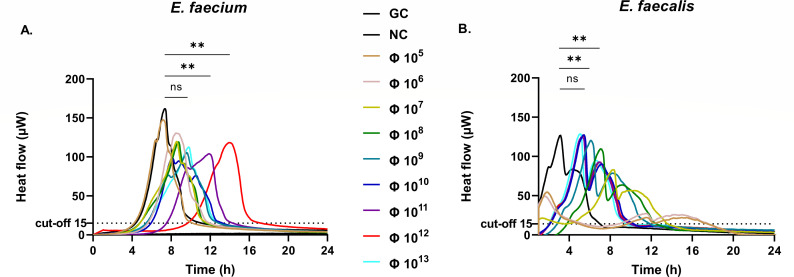 Fig 4
Fig 4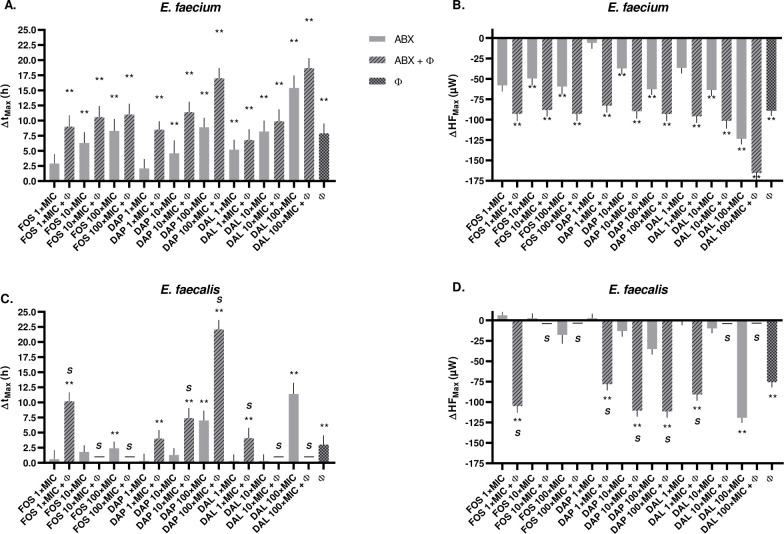 Fig 5
Fig 5| Strain | Vancomycin, | Teicoplanin, | HLG, | Tigecycline | Linezolid, | Levofloxacin, | Fosfomycin, µg/mL | Daptomycin, µg/mL | Dalbavancin, µg/mL |
|---|---|---|---|---|---|---|---|---|---|
|
| ≥32 (R/R) | <0.05 (S/S) | <500 (S/S) | <0.12 (S/S) | 2.0 (S/S) | ≥8 (R/R) | 64.0 (S/S) | 0.5 (S/S) | 0.023 (S/S) |
|
| ≥32 (R/R) | <0.05 (S/S) | >500 (R/R) | <0.12 (S/S) | 1.0 (S/S) | 0.5 (S/S) | 48.0 (S/S) | 0.5 (S/S) | 0.125 (S/S) |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Orthopedic Infections and Treatments · Fecal contamination and water quality
INTRODUCTION
Periprosthetic joint infections (PJI) represent a severe complication in orthopedic surgery, significantly affecting patient outcomes and healthcare systems. Among the diverse array of pathogens implicated, Enterococcus spp. account for 2 to 15% of cases and pose notable challenges due to their ability to form biofilms on prosthetic surfaces (1, 2). Biofilms not only shield bacteria from the host immune response but also significantly reduce the efficacy of conventional antibiotic therapies (3).
The clinical management of enterococcal-related PJI is further complicated by the emergence of vancomycin-resistant Enterococcus spp. (VRE), which restricts treatment options to reserve antibiotics, such as glycopeptides, oxazolidinones, or lipopeptides (4, 5). This growing resistance highlights an urgent need for alternative therapeutic approaches.
Bacteriophages (phages), viruses that specifically target bacteria, have re-emerged as a promising tool against biofilm-associated infections (6–8). Their specificity, self-amplifying nature, and biofilm-degrading capacity make them an innovative therapeutic tool for treating persistent infections associated with medical devices. Recent studies have demonstrated that lytic phages can penetrate and disrupt biofilms on orthopedic implants, reducing bacterial load and delaying regrowth (8, 9). In the context of PJI, phage therapy may offer a minimally invasive adjunct or salvage option when surgical debridement and systemic antibiotics fail (8). Importantly, phages can also act synergistically with antibiotics, offering the potential to further improve treatment efficacy (10).
Although phage-antibiotic combinations have shown promise against VRE biofilms, for instance, complete eradication of E. faecalis biofilms with phage ɸEFP01 plus vancomycin (11), comprehensive data remain limited. However, most studies have focused on planktonic cultures or a limited set of antibiotics (10), leaving a gap in knowledge regarding phage synergy with clinically relevant agents, such as dalbavancin, daptomycin, and fosfomycin, against biofilm-forming VRE.
In this context, our study aims to address this gap by isolating novel VRE-targeting phages and evaluating their antibiofilm activity in combination with last-line antibiotics, exploring viable therapeutic strategies for challenging multidrug-resistant enterococcal infections in orthopedic settings.
RESULTS
MIC determination of antibiotics and genomic analysis of Enterococcus strains
The phenotypic and genotypic profiles of both strains demonstrated that all were MDR, showing non-susceptibility to at least one agent in three or more antimicrobial categories. The MIC profiles of both isolates reflected their genotypic resistance backgrounds. Vancomycin resistance (≥32 µg/mL) corresponded with the presence of the vanHBX operon in both strains, while their susceptibility to teicoplanin (MIC < 0.05 µg/mL) aligns with the known phenotype of vanB-type resistance, which typically confers resistance to vancomycin but not teicoplanin. Additionally, E. faecalis displayed high-level gentamicin resistance, consistent with the presence of aminoglycoside-modifying enzymes (ant (6)-Ia and aph(3′)-III), whereas E. faecium, which carried aac(6′)-Ii, remained below the high-level resistance breakpoint. Levofloxacin resistance in E. faecium could not be attributed to acquired resistance genes, indicating the likely involvement of chromosomal mutations. In contrast, all other tested agents (linezolid, tigecycline, fosfomycin, dalbavancin, and daptomycin) displayed phenotypic susceptibility consistent with genomic predictions (Table 1; Fig. 1). Whole-genome sequencing (WGS) revealed distinct virulence gene repertoires in E. faecium and E. faecalis (Fig. 2). The E. faecium isolate carried a relatively limited set of virulence determinants, including adhesins (acm, ecbA, scm), the aggregation factor fss3, and the immune evasion gene sgrA. In contrast, E. faecalis harbored a broader spectrum of virulence-associated genes across multiple functional categories. These included adhesins (ace, efaA, EF0485), pili/aggregation genes (ebpA- C, srtC, EF0149, fss2-3, prgB/asc10, fss1), and biofilm-related loci (sprE, fsrA-C, bopD, EF0818). Additional determinants comprised immune evasion genes (cpsA - K), proteases (gelE, EF3023), and the cytolysin operon (cylR1-R2, cylA, cylS-M). PHASTER analysis further identified one intact prophage region in E. faecium and two intact prophage regions in E. faecalis. Together, these findings highlight the higher virulence potential of the E. faecalis isolate, particularly in traits linked to biofilm formation, toxin production, and immune evasion.
*Correlation of genomic predictions with phenotypic resistance profiles in VRE strains. Heatmap illustrating the relationship between WGS-predicted resistance determinants and MIC-based susceptibility results for E. faecium and E. faecalis. The scale denotes the level of correlation: 2 = gene present and phenotype confirmed; 1 = inconsistent results between genotype and phenotype; and 0 = gene absent and phenotype confirmed. Both strains carried the vanHBX operon, correlating with vancomycin resistance and teicoplanin and dalbavancin susceptibility consistent with a vanB-type phenotype. High-level gentamicin resistance was observed in E. faecalis, matching the presence of ant (6)-Ia and aph(3′)-III, whereas E. faecium carried aac(6′)-Ii but remained phenotypically susceptible. Levofloxacin resistance in E. faecium was not explained by acquired resistance genes, suggesting chromosomal mutations. All other agents (linezolid, tigecycline, fosfomycin, dalbavancin, and daptomycin) showed phenotypic susceptibility in line with genomic predictions. , MIC was not determined; MLS, macrolides/lincosamides/streptogramins.
Distribution of virulence-associated genes in E. faecium and E. faecalis. Heatmap representation of virulence determinants identified by WGS in the two VRE isolates. Genes are grouped by functional categories: adhesins, pili/aggregation, biofilm formation, immune evasion, proteases/invasion, and toxins. The scale bar indicates: 1 = gene presence, 0 = gene absence. E. faecium harbored adhesin genes (acm, ecbA, scm), the aggregation factor (fss3), and immune evasion gene (sgrA), whereas E. faecalis carried a broader set of virulence determinants, including multiple adhesins (ace, efaA, EF0485), pili genes (ebpA-C, srtC, EF0149, fss2-3, prgB/asc10, fss1), biofilm-associated factors (sprE, fsrA-C, bopD, EF0818), immune evasion (cpsA-K), proteases (gelE, EF3023), and cytolysin toxin genes (cylR1-R2, cylA, cylS-M). PHASTER analysis identified one intact prophage region in E. faecium and two intact prophage regions in E. faecalis.
Genomic characterization of isolated phages
The isolated phages, CUB-FM targeting E. faecium and CUB-FS targeting E. faecalis, were characterized by WGS to determine their suitability for phage therapy. CUB-FM has a short dsDNA genome of 18,153 bp and is closely related to Enterococcus phage Athos (National Center for Biotechnology Information [NCBI] accession number LR990834; BLASTn analysis: 99% coverage and 92.65% sequence identity). The intergenomic similarities were calculated, revealing that CUB-FM belongs to the same unclassified genus within the Salasmaviridae family (Fig. S1). After genome annotation, neither indications for a lysogenic lifecycle nor antibiotic resistance and virulence genes were found. The strictly lytic nature of this phage was further validated using Phage.AI. Phage CUB-FS contains a 142,310 bp dsDNA genome and is related to Enterococcus phage phiEF24C (NCBI accession number NC_009904; BLASTn analysis: 91% coverage and 98.56% sequence identity). The intergenomic distance between these related viruses showed CUB-FS represents a new species within the Kochikohdavirus of the Brockvirinae within the Herelleviridae family (Fig. S1). It also does not contain any unwanted genes and can be concluded as a lytic phage.
Transmission electron microscopy (TEM) visualization of isolated phages
TEM analysis of phage CUB-FM revealed a slightly elongated icosahedral head approximately 47 nm in diameter and a short tail measuring about 19 nm in length (Fig. 3A through C). The tail appeared non-contractile and lacked visible tail fibers, consistent with members of the Salasmaviridae family. The CUB-FS displays an icosahedral head (77 nm in diameter) and a contractile tail (197 nm in length, 20 nm in width) with no visible tail fibers (Fig. 3D through F). A baseplate structure was observed at the distal end of the tail, supporting classification within this family. These features, together with genomic analysis, confirm that CUB-FS belongs to the Herelleviridae family.
TEM of the lytic enterococcal phages characterized in this study. (A–C) show phage CUB-FM infecting E. faecium at different magnifications with scale bars of 100 (A), 50 (B), and 25 nm (C). (D–F) show phage CUB-FS infecting E. faecalis at 100 (D), 50 (E), and 25 nm (F). Phage suspensions were applied to carbon-coated, glow-discharged Ni-mesh grids and negatively stained with 1% (w/v) uranyl acetate. Images were acquired using a Zeiss EM 906 operating at 80 kV. Intact virions with icosahedral heads and visible tails are clearly discernible.
Phage efficacy testing
Phages propagated on E. faecium as well as E. faecalis consistently reached exceptionally high concentrations of around 4 × 10^13^ PFU/mL. For E. faecium, 10^12^ PFU/mL was identified as the most effective concentration (Fig. 4A). The mean bacterial heat detection time following treatment with 10^12^ PFU/mL of phages was significantly prolonged (15.6 h) compared to GC (7.4 h), P = 0.008. Further increasing the phage titer to 10^13^ PFU/mL did not provide additional benefit, as the heat detection time decreased to 9.1 h. In contrast, for E. faecalis, at lower phage concentrations (10^5^–10^7^ PFU/mL), heat flow curves displayed an early metabolic peak (~1.8 h), followed by regrowth (~11.6 h), reflecting partial suppression and rebound of the biofilm population. In contrast, higher titers (10^8^ and 10^9^ PFU/mL) suppressed bacterial metabolism from the start (near-baseline heat flow) and significantly delayed peak activity (7.0 and 6.3 h, respectively) compared to GC (3.3 h), P < 0.01 for both, indicating the most effective antibiofilm activity at these concentrations. There was no significant difference when the phage titer was further increased to 10^10^ PFU/mL (4.7 h), P = 0.08 (Fig. 4B). Therefore, based on previous studies and commonly used phage titers in therapeutic applications (6, 12), a dose of 10^8^ PFU/mL was selected for further analysis.
*Microcalorimetry analysis of E. faecium (A) and E. faecalis (B) treated with corresponding phages at different concentrations, from 105 to 1013 PFU/mL. Dashed lines represent time to 15 µW heat flow (i.e., thermal growth onset time). For E. faecalis at lower phage concentrations (105–107 PFU/mL), heat flow curves displayed an early metabolic peak (~1.8 h), followed by regrowth (~11.6 h), reflecting partial suppression and rebound of the biofilm population. In contrast, higher titers (10⁸ PFU/mL) suppressed bacterial metabolism from the start (near-baseline heat flow) and markedly delayed peak activity, indicating the most effective antibiofilm activity at these concentrations. Φ, phage (CUB-FM for E. faecium and CUB-FS for E. faecalis). *, P < 0.01.
Phage-antibiotic synergistic effect against biofilm bacteria
Antibiotics alone exhibited dose-dependent antibiofilm activity. For E. faecium, dalbavancin at 100× MIC achieved the strongest effect compared to the growth control, reducing HF_Max_ from 210.4 ± 6.5 µW (growth control) to 86.8 ± 1.7 µW and increasing t_Max_ from 5.2 ± 1.0 h to 19.0 ± 1.8 h (P < 0.001). Daptomycin and fosfomycin at 100× MIC also led to significant regrowth delays (t_Max_: 12.5 ± 1.2 and 11.9 ± 1.7 h, respectively; both P < 0.001). In contrast, lower concentrations (1× MIC) of fosfomycin or daptomycin showed limited efficacy, with t_Max_ values not significantly different from the control. In E. faecalis, only high concentrations of antibiotics demonstrated notable activity. Dalbavancin at 100× MIC delayed t_Max_ from 3.6 ± 1.1 to 15.0 ± 1.5 h (P < 0.001), while fosfomycin and daptomycin at the same concentration yielded t_Max_ values of 6.0 ± 0.0 h (P = 0.007) and 10.6 ± 1.2 h (P = 0.002), respectively (Fig. 5; Table S1; Fig. S2).
*Effect of combined antibiotic-phage treatments on E. faecium and E. faecalis biofilms. FOS, fosfomycin; DAP, daptomycin; DAL, dalbavancin; Φ, phage (CUB-FM for E. faecium and CUB-FS for E. faecalis); ABX, antibiotics; S, synergy; −, complete biofilm eradication (no heat flow detected); *, P < 0.01. Bar graphs show mean ± SD changes in (A) tMax and (B) HFMax in E. faecium and (C) tMax and (D) HFMax in E. faecalis relative to untreated growth controls following combined antibiotic-phage treatment at indicated multiples of MIC. Conditions include fosfomycin + phage, daptomycin + phage, and dalbavancin + phage or corresponding phage monotherapies. Positive ΔtMax values indicate delayed time to maximum heat flow; negative ΔHFMax values indicate reduced metabolic activity. Data are from biological triplicates. One-way ANOVA with Tukey’s post-hoc test was used. Synergy is defined as a combined effect exceeding the sum of the effects of the individual agents, whereas additive effects represent improved activity over monotherapy but within the expected sum of individual effects.
Combining phages with antibiotics significantly enhanced antibiofilm activity across both species. In E. faecium, the combination of CUB-FM with all antibiotics produced an additive effect, with activity greater than either treatment alone but not exceeding the expected sum of their individual effects. The combination of dalbavancin at 100× MIC with CUB-FM yielded the most potent suppression (HF_Max_: 45.0 ± 7.6 µW; t_Max_: 22.3 ± 1.3 h; P < 0.001), exceeding either agent alone. Similar additive effects were observed with daptomycin and fosfomycin: t_Max_ increased to 20.6 ± 1.4 and 14.6 ± 1.5 h, respectively, when combined with CUB-FM (P < 0.001 for both). In E. faecalis, complete biofilm eradication (no heat flow detected) was achieved when CUB-FS was combined with dalbavancin or fosfomycin at 10× and 100× MIC. Notably, even at 1× MIC, these combinations showed synergistic effect (Δt_Max_ 4.1 h and 10.2, respectively), highlighting the potentiating effect of phage treatment. Daptomycin at 1× MIC combined with phage produced an additive effect, with activity greater than either treatment alone but not exceeding the expected sum of their individual effects, whereas at 10× and 100× MIC showed synergy (Δt_Max_ 7.4 h and 22.1, respectively), significantly reducing biofilm metabolic activity beyond either monotherapy. HF_Max_ values in combination therapy also showed consistent and significant reductions compared to monotherapies (Fig. 5; Table S1; Fig. S2).
DISCUSSION
This study addresses a critical need for novel strategies against vancomycin-resistant Enterococcus spp*.*, particularly in biofilm-associated infections, such as PJIs. We isolated two novel bacteriophages targeting E. faecium and E. faecalis and evaluated their therapeutic potential both alone and in combination with antibiotics commonly used for recalcitrant gram-positive infections: dalbavancin, daptomycin, and fosfomycin.
In selecting antibiotics for this study, we aimed to cover diverse mechanisms of action while including agents of clinical relevance in the management of multidrug-resistant gram-positive infections. Dalbavancin, daptomycin, and fosfomycin were chosen because they represent last-line therapeutic options in VRE-associated infections, have distinct targets (cell wall synthesis, membrane depolarization, and peptidoglycan precursor inhibition, respectively) (13), and are used in clinical scenarios where phage therapy might be considered as an adjunct (14, 15). Although dalbavancin as a lipoglycopeptide is mechanistically related to vancomycin, its inclusion allowed us to evaluate whether differences in binding affinity and spectrum could yield divergent activity against vanB-type VRE. As confirmed by MIC testing, both isolates demonstrated high-level vancomycin resistance consistent with their genotype, underscoring the need to explore alternative or combination strategies. In this study, antibiotic concentrations up to 100× MIC were applied in order to probe the upper range of antibiofilm activity. While such concentrations clearly exceed those achievable through systemic administration, they serve to illustrate the marked tolerance of biofilm-embedded bacteria compared to their planktonic counterparts. Importantly, these high concentrations are not without clinical relevance since comparable levels can be achieved through local delivery strategies, such as antibiotic-loaded spacers, beads, or bone cement, which provide high local drug release without systemic toxicity (16, 17). Thus, although 100× MIC data should be interpreted cautiously, they provide valuable insight into the potential limits of antibiotic efficacy within biofilms and emphasize the need for combination approaches, including phage therapy, to overcome biofilm-associated tolerance.
Antibiotic monotherapies exhibited variable antibiofilm efficacy with notable discrepancies compared to MIC-based susceptibility profiles. While MIC testing indicated susceptibility to fosfomycin, daptomycin, and dalbavancin under planktonic conditions, their activity was markedly reduced in the biofilm model. This highlights the known limitation of MIC assays in predicting efficacy against biofilm-associated infections, where bacterial communities exhibit increased tolerance due to physical barriers and altered metabolic states (18, 19). Among the agents tested, dalbavancin demonstrated comparable activity in both MIC and biofilm assays, where at 1× MIC, it significantly delayed bacterial growth of E. faecium compared to growth control. This aligns with previous studies reporting dalbavancin’s capacity to inhibit biofilm formation and reduce biomass in gram-positive pathogens (13, 20). Daptomycin and fosfomycin also exhibited dose-dependent antibiofilm activity against E. faecium, with significant growth delays observed starting from 10× MIC. However, both showed minimal antibiofilm activity against E. faecalis biofilms. While daptomycin is established in the treatment of bloodstream infections and endocarditis, its antibiofilm efficacy may be limited by its calcium-dependent mechanism of membrane depolarization (21). Fosfomycin monotherapy has also limited efficacy against mature biofilms potentially due to suboptimal biofilm matrix accumulation and resistance development when used alone (22). Nonetheless, its favorable pharmacokinetics, including low molecular weight and effective tissue diffusion, suggest its value as part of combination regimens, rather than a standalone agent (23).
Although certain antibiotics demonstrated measurable antibiofilm activity in our assays, their efficacy was incomplete and, in several cases, markedly reduced compared to planktonic conditions. The rationale for introducing phages lies in their complementary advantages: phages can actively degrade biofilm structure, replicate at the site of infection, and limit the emergence of resistance by applying a second, independent mechanism of bacterial killing. Thus, rather than replacing active antibiotics, phages provide an additional layer of activity that enhances and prolongs antibiotic efficacy, particularly in the context of biofilm-associated VRE infections where treatment options remain limited.
In parallel, the genomic characterization of the two isolates revealed distinct virulence gene repertoires, with E. faecium carrying a limited set of adhesins and immune evasion genes, while E. faecalis harbored a broader array of adhesins, pili, biofilm-associated loci, and cytolysin genes. This more extensive virulence profile in E. faecalis is consistent with its stronger biofilm tolerance and higher pathogenic potential. Importantly, these findings aligned with the MIC data: both isolates displayed high-level vancomycin resistance in agreement with their van operons, while E. faecalis additionally showed high-level gentamicin resistance corresponding to aminoglycoside-modifying enzyme genes detected by WGS. Such genotype-phenotype concordance underscores the value of WGS as a predictive tool while reinforcing the importance of phenotypic validation in antimicrobial testing.
For phage characterization, we prioritized genomic analysis to assess therapeutic safety. Genomic screening provides important insights into phage biology, including genome size, open reading frames, and the absence of genes associated with virulence or antimicrobial resistance, which are crucial for clinical safety. Moreover, conserved and divergent genomic regions identified by phylogenetic analyses help classify phages and guide their therapeutic selection. Equally important is the exclusion of harmful genes while recognizing beneficial features, such as anti-CRISPR loci, which may enhance phage efficacy by overcoming bacterial defense systems. Together, these genomic insights support the rationale for developing safe and effective phages against multidrug-resistant Enterococcus spp. (10).
Our analyses confirmed both isolated phages as suitable candidates. CUB-FM (92.65% identity to Enterococcus phage Athos, Salasmaviridae) showed a strictly lytic profile without lysogeny, virulence, or resistance genes, and TEM confirmed a podovirus morphology with an icosahedral head and a short non-contractile tail (24, 25). CUB-FS showed high sequence identity (98.56%) to phage phiEF24C, within the Herelleviridae family. Like CUB-FM, CUB-FS lacked lysogeny- or resistance-associated genes and displayed classic Herelleviridae morphology with an icosahedral head and a contractile tail. TEM confirmed the presence of a baseplate structure.
Interestingly, phages propagated on E. faecium and E. faecalis consistently reached higher titers (~4 × 10¹³ PFU/mL). However, in the efficacy assays, different titers were identified for different bacterial strains, and relatively high phage titers were employed to ensure reproducible activityPEER REVIEW HISTORY against biofilm-embedded E. faecium. This discrepancy likely reflects differences in host-phage interactions, including replication dynamics, burst size, and adsorption efficiency (26, 27). Such host-dependent variability in phage yield has been described previously and highlights the importance of host selection in achieving high-titer preparations for therapeutic applications (28). In the efficacy assays, relatively high phage titers were employed to ensure reproducible activity against biofilm-embedded bacteria. This approach reflects the well-recognized challenge that biofilms pose as a physical and physiological barrier, often necessitating higher phage inputs compared to planktonic cultures. While such concentrations may exceed those typically achievable with systemic administration, they are relevant for therapeutic strategies based on local delivery. Intra-articular instillation, intraoperative irrigation, or the use of phage-loaded biomaterials can yield locally elevated titers without systemic toxicity, as already reported in preclinical and compassionate-use settings (8). Thus, the high titers used in vitro should be viewed as proof-of-concept conditions to demonstrate antibiofilm efficacy. Future studies are warranted to establish the minimal effective dose under clinically achievable conditions and to optimize phage-antibiotic combinations for translation into therapeutic applications.
In microcalorimetry assays, CUB-FM exhibited optimal biofilm suppression at 10¹² PFU/mL. Interestingly, higher titers (10¹³ PFU/mL) paradoxically decreased efficacy, potentially due to lysis inhibition or quorum sensing interference at high phage densities (29–31). CUB-FM, thus, emerges as a promising therapeutic candidate, demonstrating genomic safety and optimal performance at mid-to-high phage concentrations.
Similarly, antibiofilm assays with CUB-FS revealed a plateau effect beyond 10⁸ PFU/mL, suggesting phage saturation or limited penetration into dense biofilm matrices. This aligns with previous observations indicating diminishing returns at higher phage titers in mature biofilms (9, 32). CUB-FS, therefore, shows strong therapeutic potential at clinically relevant titers, supported by a favorable safety profile and consistent morphological features. However, due to the observed saturation effect, its efficacy may be better enhanced through combination with antibiotics or biofilm-disrupting agents rather than by increasing phage dose alone.
Furthermore, we evaluated the synergy between antibiotics and phages, which can lower required drug doses, reducing toxicity and delaying resistance by forcing bacteria to overcome multiple mechanisms simultaneously (33, 34). This is particularly relevant in biofilm-associated and multidrug-resistant infections, where phages enhance antibiotic penetration and disrupt biofilm tolerance (35). In our study, all phage-antibiotic combinations produced additive or synergistic effects, supported by calorimetric data showing significant reductions in HF_Max_ and extensions of t_Max_ compared to monotherapies. The combination of dalbavancin and phages yielded the most potent biofilm suppression. The strongest effect was observed for dalbavancin with phage CUB-FS in E. faecalis, where synergy at 1× MIC and complete biofilm clearance at higher concentrations were achieved, consistent with recent clinical reports (14). For E. faecium, the same combination showed additive effects, still outperforming dalbavancin alone. Although direct in vitro evidence on dalbavancin-phage combinations is limited, our results indicate that this strategy effectively inhibited biofilm formation. The underlying mechanisms likely include a reduction in the effective antibiotic concentration, enhanced drug penetration into the biofilm matrix via phage-encoded depolymerases, and suppression of resistance development, mechanisms that are probably similar to those described for vancomycin (36, 37). Daptomycin-phage combinations yielded additive activity against E. faecium and synergistic activity at higher concentrations against E. faecalis, in line with previous studies reporting enhanced phage access through daptomycin-induced membrane disruption (15, 38). Fosfomycin-phage treatment overcame the limited efficacy of fosfomycin monotherapy, showing synergy at 1× MIC and complete eradication at higher concentrations likely due to phage-mediated disruption of the biofilm matrix, enhancing fosfomycin penetration into bacterial cells, as supported by prior work (39). Similarly, further research demonstrated that combining phages with fosfomycin enhanced bactericidal activity, reduced the emergence of resistant bacterial clones, and decreased the MIC values compared to fosfomycin monotherapy (40, 41). Together, these findings highlight that phage-antibiotic synergy in enterococci may result from reduced resistance emergence, lowered antibiotic MICs, and improved biofilm penetration. This variability further suggests that tailoring phage selection to individual strains may maximize therapeutic success (36).
This study has some limitations. First, only two strains (a clinical E. faecium VRE isolate and E. faecalis ATCC 51299) were available for phage characterization, as lytic phages could not be recovered for the other screened isolates. Although this restricts representativeness, the findings provide proof of concept for phage-antibiotic synergy in biofilm-associated infections. Future work should include a broader strain panel to capture the genetic and phenotypic diversity of VRE. Second, antibiofilm activity was assessed only by isothermal microcalorimetry. This method provides sensitive, real-time, and non-invasive monitoring of metabolic activity, but it does not directly quantify bacterial numbers and requires cautious interpretation. Still, previous studies demonstrate that parameters, such as time to detection and peak heat flow, correlate well with CFU counts, supporting its validity (42).
In conclusion, this study demonstrates the potential of phage-antibiotic combinations, particularly involving dalbavancin and fosfomycin, as a powerful strategy against Enterococcus spp. biofilms. The novel lytic phages CUB-FM and CUB-FS showed strain-specific efficacy and genomic safety profiles, supporting their therapeutic use. Importantly, the synergistic interactions observed in our in vitro models may offer a pathway to improved clinical outcomes in complex biofilm-associated infections, including those caused by vancomycin-resistant enterococci. Further in vivo studies and clinical trials are warranted to explore optimal dosing regimens and to validate these promising in vitro findings.
MATERIALS AND METHODS
Bacterial strains and bacteriophages
Initially, four clinical Enterococcus faecium VRE isolates, one clinical Enterococcus gallinarum VRE isolate, and the reference Enterococcus faecalis VRE ATCC 51299 strain were included for screening. However, lytic phages were recovered only against one clinical E. faecium isolate and E. faecalis ATCC 51299, which were, therefore, selected for further characterization. Clinical isolates of Enterococcus spp*.* obtained from a bone and joint infection site (Labor Berlin—Charité Vivantes GmbH, Berlin, Germany) and the laboratory reference strain E. faecalis ATCC 51299 were stored at −80°C using a cryovial bead preservation system (Microbank, Pro-Lab Diagnostics, Canada). For bacterial strains, DNA was extracted using the DNeasy UltraClean Microbial Kit (Qiagen) and prepared for whole-genome sequencing on an Illumina Miniseq device as previously described (43). An assembly was constructed using Unicycler (Galaxy v0.5.1) (44). ResFinder (v4.7.2; DTU Center for Genomic Epidemiology; minimum DNA identity of 90% and minimum coverage of 60%) was used to determine resistance genes present in the isolates (45). Virulence factors and biofilm-associated genes were identified using the same thresholds from the VFDB database (46). PHASTEST v3.0 (47) was run for prophage detection in the genomes.
Strictly lytic bacteriophages targeting E. faecium (CUB-FM phage) and E. faecalis (CUB-FS phage) were isolated from hospital sewage and subsequently employed in experimental procedures. The phage isolation procedure employed an enrichment method detailed in a previous study (48). To ensure phage purity, four consecutive cycles of single-plaque isolation were conducted on the bacterial host strain. For propagation, the host strain was grown overnight at 37°C in tryptic soy broth (TSB; US Biological, Eching, Germany). Next, 1 mL of phage lysate at a concentration of approximately 10⁸ PFU/mL was incubated at 4°C for 1 h. An aliquot of 0.2 mL overnight culture was inoculated into 20 mL TSB and incubated with shaking at 37°C until OD₆₀₀ reached 0.4, after which 0.1 mL of the phage suspension was added. The culture was incubated until complete clearing (~5 h or overnight if necessary). The lysate was centrifuged at 4,000 × g for 20 min, and the supernatant was sterilized using 0.22 µm filter. Polyethylene glycol (PEG) precipitation was carried out by adding PEG/NaCl stock solution (20% w/v PEG-8000, 2.5 M NaCl; PanReac AppliChem, Darmstadt, Germany) to the lysate at 25% of its volume (e.g., 250 µL per 1 mL lysate), mixing by inversion, and incubating on ice for ≥1 h. Phages were pelleted by centrifugation at 13,000 × g (4°C, 40 min), the supernatant discarded, and residual liquid removed. Pellets were gently resuspended in 100 µL SM buffer (10 mM Tris-HCl, pH 7.5, 100 mM NaCl, 8 mM MgSO₄) and incubated at room temperature for 1 h with occasional pipetting. Final titers were determined by plaque assay on the host strain by serial dilution. Subsequently, the phage was cultured in a liquid medium, with bacterial strains grown in tryptic soy broth (TSB) (US Biological, Eching, Germany) overnight at 37°C. These phage stocks were used for whole-genome sequencing analysis as detailed in Sharifi et al. (49). The phage’s taxonomy was determined using Virus Intergenomic Distance Calculator (VIRIDIC) v1.1 analysis (50). The phage genomes of CUB-FM and CUB-FS were submitted to NCBI GenBank and are available under accession numbers PV646496 and PV646497, respectively.
Antibiotics
Dilution of antibiotics was prepared according to both Clinical and Laboratory Standards Institute (CLSI) (51) and European Committee on Antimicrobial Susceptibility Testing (EUCAST) (52). Fosfomycin was dissolved in sterile distilled water with glucose-6-phosphate (500 µL glucose-6-phosphate per 10 mL of Muller Hinton Broth [MHB]), daptomycin in sterile distilled water with calcium chloride, and dalbavancin in dimethyl sulfoxide DMSO) with polysorbate-80, immediately prior to utilization.
Morphological analysis of the phages by TEM
The morphology of the phages was analyzed by TEM following negative staining, as previously described (53). Briefly, 15 µL of phage suspension was placed onto parafilm and then transferred onto a carbon-coated, glow-discharged (Leica Microsystems, Wetzlar, Germany) Ni-mesh grid (G2430N; Plano GmbH, Wetzlar, Germany). The phage particles were allowed to adsorb for 10–15 min at room temperature. Subsequently, the grids were washed three times with distilled water and negatively stained with 1% aqueous uranyl acetate (SERVA Electrophoresis GmbH, Heidelberg, Germany) for 20 s. After air drying, the grids were examined with a Zeiss EM 906 transmission electron microscope (Carl Zeiss Microscopy Deutschland GmbH, Oberkochen, Germany) at a voltage of 80 kV. Phage dimensions were determined with the image processing software ImageJ.JS (54, 55).
MIC determination of antibiotics and phage efficacy testing
Minimal inhibitory concentrations (MICs) for the antibiotics were obtained by the E-test (Liofilchem, Roseto degli Abruzzi, Italy) on Muller Hinton agar based on CLSI breakpoints and EUCAST ECOFFs (51, 52).
To determine phage efficacy, biofilms of E. faecium as well as E. faecalis were formed on sterile porous glass beads (diameter 4 mm, pore sizes 60 µm, ROBU1, Hattert, Germany) (56), washed three times with 2 mL 0.9% NaCl to remove planktonic bacteria, and subsequently exposed to the phage dilutions ranging from 10^5^ to 10^13^ PFU/mL. The effects of phage treatment were monitored over a 24 h period using isothermal microcalorimetry.
Antibiotics alone and phage-antibiotic combinations against biofilm
After biofilm formation as described above, beads were transferred to 24-well plates containing 1 mL of MHB with different concentrations of the antibiotic (1, 10, and 100× MIC) or phage (at defined PFU/mL) as monotherapy or phage-antibiotic combinations. For all phage-antibiotic combinations, biofilms were initially treated with 500 µL of phage solution in MHB at twice the intended final concentration, ensuring that after the subsequent addition of the antibiotic, the concentration of both agents would be at the desired level. The phage treatment was carried out for 4 h at 37°C. Following this, 500 µL of the corresponding antibiotic solution also prepared at twice the desired final concentration was added, and the biofilms were incubated for an additional 20 h at 37°C. After a total incubation of 24 h, treated biofilm beads were rinsed three times with 0.9% NaCl, placed in sterile glass ampules with 3 mL fresh TSB, and inserted in the calorimeter, where heat produced by viable bacteria present in the bead after 24 h of treatment or no treatment (growth control) was monitored for 36 h.
Microcalorimetry setup and heat measurement assay
Biofilm treatment efficacy was assessed using an isothermal calorimeter TAM III (TA Instruments, New Castle, DE, USA), as previously described (57, 58). This technique allows real-time monitoring of bacterial metabolic activity by measuring heat flow as an indirect indicator of microbial replication. To evaluate the antimicrobial effects of the treatments, microcalorimetric data were collected by recording the heat generated from the regrowth of viable bacteria following 24 h of exposure to antibiotics, phages, or their combinations. The assay measured two primary parameters: (i) maximum heat flow peaks (HF_Max_, µW) to assess metabolic activity; and (ii) time to reach maximum heat (t_Max_, h) to evaluate delays in bacterial regrowth. To facilitate comparison between treatments, results were expressed as Δt_Max_ and ΔHF_Max_ relative to the untreated growth control for each strain and experiment:
Δt_Max_ = t_Max_treatment − t_Max_control
ΔHF_Max_ = HF_Max_treatment − HF_Max_control
Positive Δt_Max_ values indicate delayed bacterial growth relative to the control, while negative ΔHF_Max_ values indicate reduced metabolic activity. Measurements were recorded every 120 s over a 36 h period. A dose-dependent antimicrobial effect was indicated by a reduction in HF_Max_ values and a delay in t_1Max_ compared to the control, in line with previous methodologies (57, 59). The synergy between antibiotics and phages was defined as a combined effect exceeding the sum of individual effects, whereas additive effects represent improved activity over monotherapy but within the expected sum of the individual effects. All experiments were performed in triplicate to ensure reproducibility.
Statistical analysis
Results were expressed as mean and standard deviation for normally distributed variables (further analyzed by Student t test) or median and minimum/maximum for non-normally distributed variables (analyzed by Mann-Whitney U test). Hypothesis testing was two-tailed, with P < 0.05 considered statistically significant. Software GraphPad Prism v9.3 (GraphPad Software Inc., La Jolla, CA, USA) was used for statistical analysis and for graphics.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Renz N, Trebse R, Akgün D, Perka C, Trampuz A. 2019. Enterococcal periprosthetic joint infection: clinical and microbiological findings from an 8-year retrospective cohort study. BMC Infect Dis 19:1083. doi:10.1186/s 12879-019-4691-y 31881851 PMC 6935141 · doi ↗ · pubmed ↗
- 2Tai DBG, Patel R, Abdel MP, Berbari EF, Tande AJ. 2022. Microbiology of hip and knee periprosthetic joint infections: a database study. Clin Microbiol Infect 28:255–259. doi:10.1016/j.cmi.2021.06.00634129907 PMC 8665939 · doi ↗ · pubmed ↗
- 3Trampuz A, Piper KE, Jacobson MJ, Hanssen AD, Unni KK, Osmon DR, Mandrekar JN, Cockerill FR, Steckelberg JM, Greenleaf JF, Patel R. 2007. Sonication of removed hip and knee prostheses for diagnosis of infection. N Engl J Med 357:654–663. doi:10.1056/NEJ Moa 06158817699815 · doi ↗ · pubmed ↗
- 4Janice J, Wagner TM, Olsen K, Hegstad J, Hegstad K. 2024. Emergence of vancomycin-resistant enterococci from vancomycin-susceptible enterococci in hospitalized patients under antimicrobial therapy. J Glob Antimicrob Resist 36:116–122. doi:10.1016/j.jgar.2023.12.01038128726 · doi ↗ · pubmed ↗
- 5Levitus M, Rewane A, Perera TB. 2025. Stat Pearls Publishing Copyright © 2025, Stat Pearls Publishing LLC., Treasure Island (FL) ineligible companies. Disclosure: Ayesan Rewane declares no relevant financial relationships with ineligible companies. Disclosure: Thomas Perera declares no relevant financial relationships with ineligible companies. Vancomycin-resistant enterococci. Stat Pearls.
- 6Zagaliotis P, Michalik-Provasek J, Gill JJ, Walsh TJ. 2022. Therapeutic bacteriophages for Gram-negative bacterial infections in animals and humans. Pathog Immun 7:1–45. doi:10.20411/pai.v 7i 2.516PMC 959613536320594 · doi ↗ · pubmed ↗
- 7Walter N, Mirzaei M, Deng L, Willy C, Alt V, Rupp M. 2024. The potential of bacteriophage therapy as an alternative treatment approach for antibiotic-resistant infections. Med Princ Pract 33:1–9. doi:10.1159/000534717 PMC 1089661537879316 · doi ↗ · pubmed ↗
- 8Morovic P, Benavente LP, Karbysheva S, Perka C, Trampuz A. 2023. Bacteriophage therapy in orthopaedic and cardiovascular surgery: clinical experience in eight patients with difficult-to-treat infections. Orthopaedic Proceedings 105-B:26–26. doi:10.1302/1358-992X.2023.17.026 · doi ↗