Impact of synonymous mutations in the blaTEM-3 gene on gene expression and Escherichia coli fitness
Rinku Dhungana, Heba Kaadan, Aparna Paudel, Peter Oelschlaeger

TL;DR
Synonymous mutations in a gene for antibiotic resistance affect bacterial fitness and resistance, possibly through epigenetic or recombination mechanisms.
Contribution
Demonstrates that synonymous mutations in a β-lactamase gene influence Escherichia coli fitness and resistance, beyond transcriptional effects.
Findings
Synonymous mutations in blaTEM-3 alter bacterial fitness and resistance in antibiotic presence.
Cells with synonymous mutations outcompete others under antibiotic stress but are outcompeted in its absence.
Transcriptional and mRNA structural differences are not responsible for the observed effects.
Abstract
Synonymous mutations (SMs) have a significant effect on the expression of the β-lactamase TEM-3 and the resistance phenotype conferred. Cell-free transcription did not reproduce this effect. The amount of mRNA obtained was identical for blaTEM-3 transcripts with or without SMs, and the transcripts were indistinguishable in terms of secondary structure and thermal de- and renaturation, as shown by circular dichroism experiments. In asymmetric competition assays, cells expressing the gene with SMs replaced cells expressing the gene without SMs after five days of passaging in a low concentration of ceftazidime. However, without antibiotic present, cells expressing the gene without SMs replaced cells expressing the gene with SMs. These experiments indicate that either post-transcriptional or in cellulo processes are responsible for the differences in expression levels and fitness. Possible…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
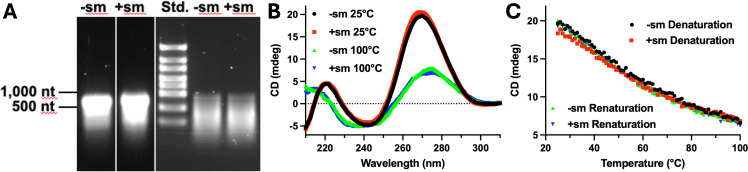 Fig 1
Fig 1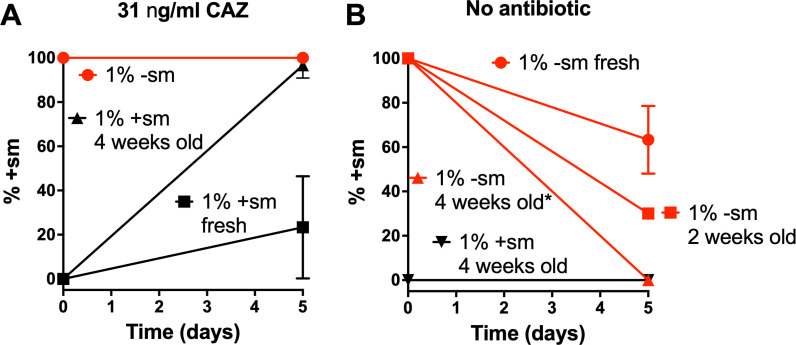 Fig 2
Fig 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial biofilms and quorum sensing · Escherichia coli research studies
OBSERVATION
In a previous report about synonymous mutations (SMs) in three genes encoding TEM β-lactamase variants, SMs had the greatest effect in blaTEM-3 (1). TEM-3 expression confers the 2be phenotype, which is characterized by resistance to extended-spectrum β-lactams, such as third-generation cephalosporins and aztreonam (2, 3). Western blots indicated TEM-3 expression in Escherichia coli DH10B cells from its published gene (4), which includes four SMs relative to the blaTEM-1a gene (5) (Fig. S1 in the Supplemental Material), was more than fourfold higher than when it was expressed from an artificial gene containing only nonsynonymous mutations (NMs). The former is designated as the blaTEM-3(+sm) gene and the latter as the blaTEM-3(-sm) gene. In disc diffusion assays with E. coli cells expressing these genes, blaTEM-3(+sm) resulted in smaller diameters than the blaTEM-3(-sm) gene. The minimum inhibitory concentration (MIC) of ceftazidime was 64 times higher with the blaTEM-3(+sm) gene, while no significant difference was observed with the other antibiotics tested.
Two avenues were pursued to decipher the underlying mechanisms responsible for the different expression and resistance levels in E. coli. Firstly, cell-free transcription and analysis of the transcript by circular dichroism (CD) spectropolarimetry were carried out to discern if transcription efficiency or stability of the transcript was affected. Cell-free transcription (MEGAscript Transcription Kit, ThermoFisher) did not result in different amounts of transcript (Fig. 1A), and the transcripts appeared to degrade at equal rates as assessed by agarose gel electrophoresis of transcript after six months of storage at −20°C and several freeze-thaw cycles (Fig. 1A). CD scans of the two transcripts at 25 and 100°C, respectively, are superimposable, indicating no difference in secondary structure (Fig. 1B), and thermal denaturation measured by CD at 270 nm was identical and completely reversible (Fig. 1C). Cell-free translation from these transcripts and coupled transcription/translation was attempted but did not yield detectable amounts of TEM-3 possibly due to insufficient formation of a disulfide bond present in TEMs (6) and improper protein folding.
(A) Agarose gel electrophoresis of transcripts obtained after cell-free transcription of the -sm and +sm genes (left side), RNA standard in the middle, and transcripts after storage at −20°C for six months and several freeze-thaw cycles (right side). (B) Circular dichroism (CD) scans of the -sm and +sm transcripts at 25°C and 100°C. (C) Thermal denaturation (from 25°C to 100°C) and renaturation (from 100°C to 25°C) of the two transcripts measured by CD signal at 270 nm.
*Results of asymmetric competition assays with ceftazidime (A) and without antibiotic (B). At day 0, the assays were started with either 1% -sm and 99% + sm cells or vice versa. At day 5, cells were harvested and plated on agar plates. Ten colonies were randomly isolated and identified based on their blaTEM-3 sequence. All experiments were carried out three times except the experiment with 2-week-old cells in panel B, which was done once. Labels of the lines in the graphs indicate the initial conditions. The results for 1% -sm in panel A and 1% + sm in panel B (horizontal lines) represent 6 (3 with fresh and 3 with 4-week-old cells) and 7 (3 with fresh, 1 with 2-week-old, and 3 with 4-week-old cells) experiments, respectively. Means ± standard deviations are shown. In some of these cells, the -sm gene was deleted as explained in the text.
Another avenue is to study the cellular background in more detail. Harboring vectors with the different genes could affect growth independent of antibiotics. Growth curves of E. coli DH10B cells harboring either pBC SK(+)-blaTEM-3(-sm) or pBC SK(+)-blaTEM-3(+sm), an empty pBC SK(+) vector, or no vector, in Müller-Hinton broth (MHB) showed no significant difference in growth (data not shown). To assess the fitness conferred by either expression of blaTEM-3(-sm) or blaTEM-3(+sm) in the presence of a small concentration (31 ng/mL or 10^−9^ times MIC) of ceftazidime, an asymmetric competition (AC) assay (7) was adapted. In short, starting with mixtures of cells expressing either the -sm or +sm genes in ratios of 1:100 or 100:1, cultures were grown in MHB in the presence of 31 ng/mL ceftazidime and passaged once per day for five days (1:50 dilution). At the end of the assay, cells were diluted and plated on agar plates (200 colonies target, actual ~50–500 colonies). Ten colonies from each plate were randomly isolated, their plasmids were extracted, and the blaTEM-3 genes were sequenced. Under these conditions, cells expressing blaTEM-3(-sm) started at 1% never replaced cells expressing blaTEM-3(+sm). In contrast, cells expressing blaTEM-3(+sm) starting at 1% constituted ~23% of colonies isolated at the end of the experiment (Fig. 2A). Interestingly, when agar plates with colonies were kept at 4°C for four weeks and then used in the same assay, cells expressing blaTEM-3(+sm) constituted almost all (~97%) of the colonies isolated. To test if the greater fitness of cells expressing blaTEM-3(+sm) was due to ceftazidime resistance, the same experiment was repeated without antibiotic present (Fig. 2B). This resulted in the reverse outcome. Cells expressing the +sm gene at an initial 1% never replaced cells expressing the -sm gene. With freshly transformed cells, about 37% of colonies picked at the end of the experiment that started with 1% -sm cells contained the plasmid encoding the -sm gene. With cells from agar plates kept at 4°C for two and four weeks, but otherwise the same conditions, this percentage increased to 70% and 100%, respectively. In the absence of antibiotics, the increased expression level of TEM-3 from the blaTEM-3(+sm) seems to lower fitness. Some of the cells also contained genes truncated after 210 nucleotides with an a109c NM, encoding a fragment with Q39—as in TEM-1—rather than K39, which is found in TEM-3. Upon closer examination, these truncated genes seem to be relics of the partially removed blaTEM-1a gene that occurred when pBC SK(+) was generated from pBluescript II (Agilent Technologies) and the blaTEM-3(-sm) gene was deleted, likely through homologous recombination.
To investigate this phenomenon further, Epsilometer tests (E-tests) (8) were performed. The results with old cells (Fig. S2A) resemble previous findings from disc diffusion assays (1), and the results for the -sm cells were comparable to the negative controls harboring the same plasmid without blaTEM-3 genes. Cells from the agar plates used in these tests were isolated, their plasmids extracted, and the blaTEM-3 genes sequenced. All the -sm plasmids had the -sm gene removed, as described above for the AC assay, while the +sm plasmids were intact. The experiment was repeated with freshly transformed cells selected for chloramphenicol resistance encoded on pBC SK(+) plus carbenicillin to ensure full-length blaTEM-3 genes. In these E-tests, the -sm cells were comparable to the +sm cells (Fig. S2B). Sequencing of the plasmids harbored by these cells confirmed that the full-length blaTEM-3 genes were still present. Further studies on why blaTEM-3(-sm) is apparently more prone to deletion than blaTEM-3(+sm) or blaTEM-1a are underway.
These results confirm that SMs in the blaTEM-3 gene increase antibiotic resistance (1). Cell-free transcription demonstrates that neither transcription efficiency nor transcript stability is responsible for this effect. The facts that the transcripts are indistinguishable in terms of secondary structure and thermal stability, as assessed by CD, and that no rare codons are used (1) suggest that translation is also unlikely to be a limiting factor. The -sm gene in the pBC SK(+) plasmids seems to be less stable than the +sm gene when there is no β-lactam selection. This could be due to epigenetic effects (9, 10). In this context, it is interesting to note that both blaTEM-1a and blaTEM-3(+sm) contain seven 5′-gatc-3′ sites for methylation by DNA adenine methyltransferase (Dam), while blaTEM-3(-sm) only contains six. Future studies will investigate the molecular mechanisms underlying the differential deletion of the -sm versus +sm blaTEM-3 genes, as well as the effects of SMs in cells and/or vectors that eliminate possible recombination.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Faheem M, Zhang CJ, Morris MN, Pleiss J, Oelschlaeger P. 2021. Role of synonymous mutations in the evolution of TEM β-lactamase genes. Antimicrob Agents Chemother 65:e 00018-21. doi:10.1128/AAC.00018-2133820762 PMC 8315918 · doi ↗ · pubmed ↗
- 2Sirot D, Sirot J, Labia R, Morand A, Courvalin P, Darfeuille-Michaud A, Perroux R, Cluzel R. 1987. Transferable resistance to third-generation cephalosporins in clinical isolates of Klebsiella pneumoniae: identification of CTX-1, a novel β-lactamase. J Antimicrob Chemother 20:323–334. doi:10.1093/jac/20.3.3233316146 · doi ↗ · pubmed ↗
- 3Bush K, Jacoby GA, Medeiros AA. 1995. A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob Agents Chemother 39:1211–1233. doi:10.1128/AAC.39.6.12117574506 PMC 162717 · doi ↗ · pubmed ↗
- 4Mabilat C, Lourençao-Vital J, Goussard S, Courvalin P. 1992. A new example of physical linkage between Tn 1 and Tn 21: the antibiotic multiple-resistance region of plasmid p CFF 04 encoding extended-spectrum β-lactamase TEM-3. Molec Gen Genet 235:113–121. doi:10.1007/BF 002861881331747 · doi ↗ · pubmed ↗
- 5Sutcliffe JG. 1978. Nucleotide sequence of the ampicillin resistance gene of Escherichia coli plasmid p BR 322. Proc Natl Acad Sci USA 75:3737–3741. doi:10.1073/pnas.75.8.3737358200 PMC 392861 · doi ↗ · pubmed ↗
- 6Villanueva M, Vostal LE, Cohen DN, Biesbrock D, Kuwaye EP, Driver SG, Hart KM. 2024. Differential effects of disulfide bond formation in TEM-1 versus CTX-M-9 β-lactamase. Protein Sci 33:e 4816. doi:10.1002/pro.481637897253 PMC 10731493 · doi ↗ · pubmed ↗
- 7Negri M-C, Lipsitch M, Blázquez J, Levin BR, Baquero F. 2000. Concentration-dependent selection of small phenotypic differences in TEM β-lactamase-mediated antibiotic resistance. Antimicrob Agents Chemother 44:2485–2491. doi:10.1128/AAC.44.9.2485-2491.200010952599 PMC 90089 · doi ↗ · pubmed ↗
- 8Nachnani S, Scuteri A, Newman MG, Avanessian AB, Lomeli SL. 1992. E-test: a new technique for antimicrobial susceptibility testing for periodontal microorganisms. J Periodontol 63:576–583. doi:10.1902/jop.1992.63.7.5761324301 · doi ↗ · pubmed ↗