Triatoma chiarii sp. nov. (Hemiptera, Reduviidae, Triatominae): a new species in the Triatoma brasiliensis complex from Rio Grande do Norte state, Brazil
Andressa Noronha Barbosa-Silva, Nathan Ravi Medeiros Honorato, Rita de Cássia Moreira de Souza, Carolina Dale, João Luís Reis Cunha, Carlos Eduardo Almeida, Samuel Alexandre Pimenta Carvalho, Ana Carolina Passos, Flávio Campos Ferreira, Paulo Marcos da Matta Guedes

TL;DR
A new species of triatomine bug, Triatoma chiarii, was discovered in Brazil and is related to species known to transmit Chagas disease.
Contribution
The discovery and taxonomic description of Triatoma chiarii sp. nov., a new species within the Triatoma brasiliensis complex.
Findings
Triatoma chiarii sp. nov. has intermediate morphological traits between T. brasiliensis and T. petrocchiae but is distinct enough to be a new species.
Geometric morphometric and phylogenetic analyses confirmed T. chiarii sp. nov. as a new taxonomic entity within the T. brasiliensis complex.
The new species was found in peridomestic environments, suggesting a potential role in Chagas disease transmission.
Abstract
A new triatomine species was discovered in the semiarid Caatinga region of Rio Grande do Norte, Brazil, where it coexists with Triatoma brasiliensis in both natural and artificial habitats. Triatomine specimens were captured in peridomestic and sylvatic environments in Rio Grande do Norte. Their identification was based on a combination of analyses, including morphology using dichotomous keys, head and hemelytron morphometry, and phylogeny using cytochrome b and internal transcribed spacer markers. The new species exhibits morphological traits that are intermediate between those of Triatoma brasiliensis and Triatoma petrocchiae, but has distinct characteristics, leading to its designation as Triatoma chiarii sp. nov. Geometric morphometric analysis of its wings and head clearly distinguished T. chiarii sp. nov. from T. brasiliensis and T. petrocchiae, while phylogenetic reconstruction…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
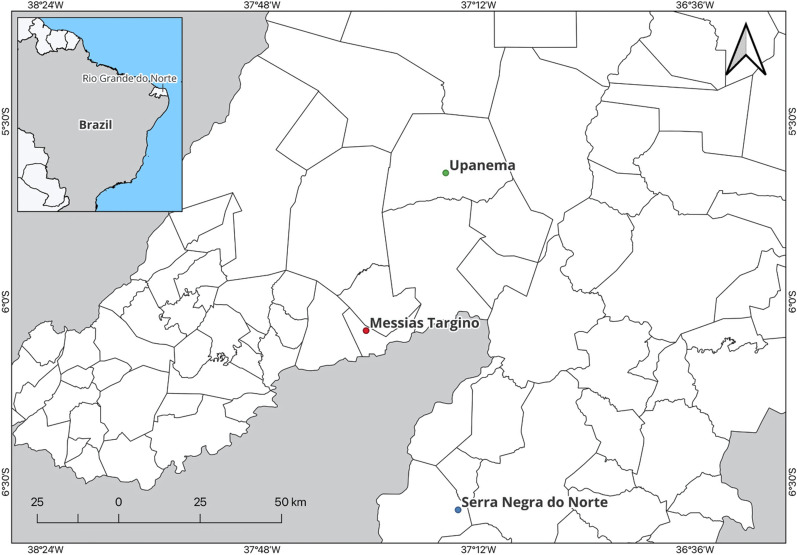 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3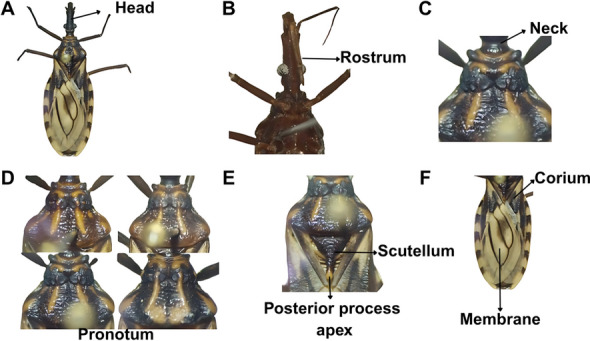 Figure 4
Figure 4 Figure 5
Figure 5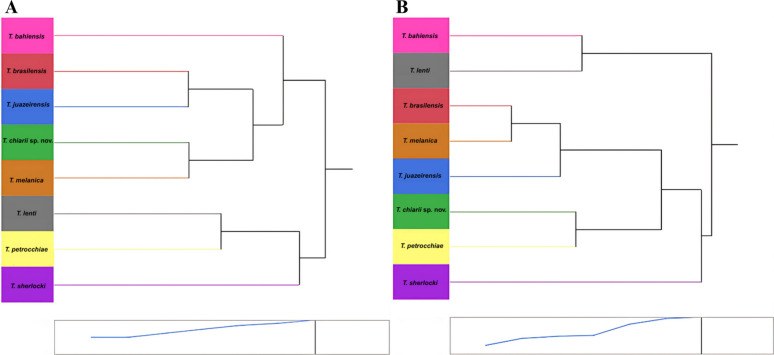 Figure 6
Figure 6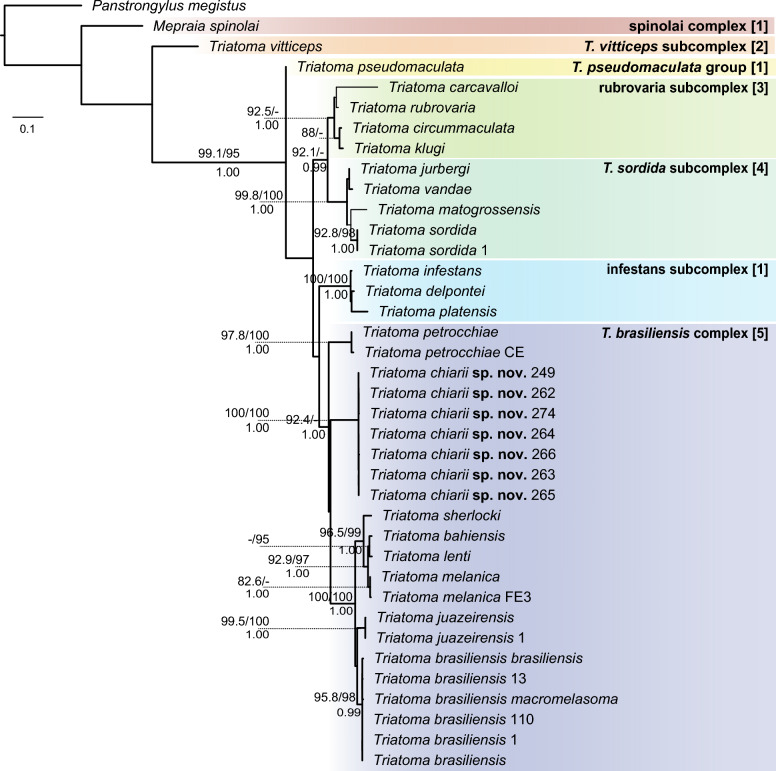 Figure 7
Figure 7- —National Council for Scientific and Technological Development – CNPq
- —CNPq/MCTI/FNDCT N◦ 22/2024 from the National Council for Scientifc and Technological Development – CNPq
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTrypanosoma species research and implications · Phytoplasmas and Hemiptera pathogens · Insect symbiosis and bacterial influences
Background
Systematic efforts in Brazil to control domiciliated Chagas disease vectors were conducted between 1975 and 1999. During this period, Triatoma infestans (Klug, 1834) was the primary vector of Chagas disease in most Brazilian states. Systematic chemical control and housing improvements led to Brazil being certified as free of T. infestans vectorial transmission of Chagas disease by 2006 [1, 2]. However, Chagas disease control remains a public health concern in various regions, as autochthonous triatomine species continue to invade and colonize domestic environments, exposing both humans and domestic animals to the risk of Trypanosoma cruzi infection [3]. Furthermore, Chagas disease transmission control faces additional challenges, particularly due to oral transmission, which primarily results from consuming açai berries, juices, and other food products contaminated with T. cruzi [4–6].
Triatoma brasiliensis Neiva, 1911 is the primary vector of Chagas disease in the Caatinga biome, which is located in the semiarid region of northeastern Brazil [7, 8]. This species is the nominal member of the Triatoma* brasiliensis* species complex [9, 10], a monophyletic group that includes the subspecies Triatoma brasiliensis* brasiliensis* and Triatoma brasiliensis* macromelasoma* Galvão, 1956, as well as the species Triatoma bahiensis Sherlock and Serafim, 1967; Triatoma juazeirensis Costa and Felix, 2007; Triatoma lenti Sherlock and Serafim, 1967; Triatoma melanica Costa, Argolo and Felix, 2006; Triatoma petrocchiae Pinto and Barreto, 1925; and Triatoma sherlocki Papa et al., 2002. These taxa exhibit variations in color patterns and morphology [11–13], as well as differences in epidemiological significance, natural history, ecological requirements, and dispersal abilities [8, 14–20].
Specimens of the genus Triatoma Laporte, 1832, which did not conform to the typical morphological characteristics outlined in established dichotomous keys [10, 11], were collected from wild and peridomestic environments in 2016 and 2017 in the state of Rio Grande do Norte, Brazil. These specimens exhibited morphological traits that were intermediate between those of T. brasiliensis and T. petrocchiae. Consequently, we describe Triatoma chiarii sp. nov., a new species within the genus Triatoma. We additionally conducted morphometric and phylogenetic analyses to assess the morphological and evolutionary relationships of T. chiarii sp. nov. in comparison to those of representatives of other species complexes.
Methods
Sampling
Eleven adult specimens of T. chiarii sp. nov. (four females and six males) were collected from rock outcrops in the sylvatic environment at the Seridó Ecological Station (06°34′37"S, 37°15′24"W), a federal conservation unit managed by the Chico Mendes Institute for Biodiversity Conservation, located in the municipality of Serra Negra do Norte, Rio Grande do Norte, Brazil. Additionally, one adult male specimen was collected from a stack of tiles located in a peridomestic environment, in the municipality of Messias Targino (06°04′47″S, 37°30′42″W), and another in Upanema (05°38′32″S, 37°17′27″W), where they co-occurred with T. brasiliensis (Fig. 1). The insects were manually captured in 2016 and 2017 using tweezers and flashlights, without the use of dislodging substances. The insects were individually stored in sealed plastic containers and maintained at – 20 °C until further analysis.Fig. 1. Map showing the exact locations (points) where the triatomine specimens analyzed in this study were collected, in the municipalities of Messias Targino, Upanema, and Serra Negra do Norte in the state of Rio Grande do Norte, Brazil
We used dichotomous keys [10, 11, 21] to examine morphological characters, and compared the collected specimens with those of the T. brasiliensis species complex deposited in the Trypanosomatid Vector Collection of the Oswaldo Cruz Foundation (FIOCRUZ-COLVET) at the René Rachou Institute, FIOCRUZ Minas Gerais, and with T. petrocchiae specimens from the Entomological Collection of the Oswaldo Cruz Institute (FIOCRUZ-CEIOC), Rio de Janeiro.
Hemelytra and head morphometry
Image acquisition and geometric morphometric analysis
The hemelytra (left wing) and the heads (dorsal view) of specimens belonging to species in the T. brasiliensis complex were photographed: T. b. brasiliensis (10 specimens, five males and five females); T. b. macromelasoma (10 specimens, five males and five females; T. melanica (eight specimens, seven males and three females); T. juazeirensis (10 specimens, five males and five females; T. lenti (10 specimens, five males and five females); T. petrocchiae (10 specimens, five males and five females); T. sherlocki (10 specimens, five males and five females); and T. chiarii sp. nov. (16 specimens, nine males and seven females). Images were obtained using a Leica Automontage Magnifier (DMC 2900). Pinned specimens were stabilized with modeling clay, and their wings were kept intact.
Next, 10 type I landmarks were selected on both the hemelytra and heads (Fig. 2A, B) of each specimen [22] by utilizing previously acquired images and TPSdig software version 2.31 [23]. All landmarks were defined at points where distinct structural features converge [24].Fig. 2. Landmarks on the A hemelytron and B head used for the morphometric analysis. (Photographs by CD)
Data transformation and multivariate analysis
Data were converted into numerical coordinates and organized into a weighted matrix in NTS format using TPSrelw software version 1.75 [23]. After generating the matrix, the centroid size and the uniform* x* and y components were calculated for each specimen, following the methodology described by Coutinho [25] and Passos et al. [26]. The landmark coordinates were then subjected to Procrustes superimposition [24], followed by thin plate spline transformation and discriminant analysis. Multivariate analyses and factorial maps were subsequently generated using JMP software version 17 [27].
Principal component analysis (PCA) and canonical correlation analysis were performed using morphometric data from the hemelytra and heads to determine the differences between the evaluated species. Procrustes coordinate analysis and multivariate ANOVA were subsequently conducted to assess the shape variability. These data were used to generate factorial maps and dendrograms, employing Mahalanobis distances for clustering analysis. Finally, statistical tests (Wilks’ lambda, Pillai’s trace, Hotelling-Lawley trace, and Roy’s max root) were performed for both hemelytra and head analyses using JMP software version 17 [27].
Molecular analysis
DNA extraction and sequencing
DNA was extracted from two legs of each of the 14 triatomine specimens, including seven Triatoma chiarii sp. nov., three Triatoma brasiliensis, one Triatoma juazeirensis, one Triatoma melanica, one Triatoma petrocchiae, and one Triatoma sordida, using the Wizard® Genomic DNA Purification Kit (Promega, Madison, WI), following the manufacturer’s instructions. DNA quantification was performed using a NanoDrop™ 2000 spectrophotometer (Thermo Scientific®, Waltham, MA). The cytochrome b (cytb) and internal transcribed spacer-1 (ITS-1) regions were amplified using specific primers (cytb [28] and [29]; ITS-1 [30]), following the amplification conditions recommended by the respective authors. PCR products were purified and sequenced using an ABI 3730XL automated sequencer (Applied Biosystems) at the René Rachou Institute–FIOCRUZ Minas Gerais. Sequences from 14 individuals were newly generated in this study, manually checked for quality, and deposited in GenBank (Additional file 1: Table S1).
Phylogenetic analysis
Cytb and ITS-1 sequences generated herein were aligned with other sequences from GenBank (Additional Table S1). Alignments were conducted using the Geneious algorithm in Geneious Prime for cytb [428 base pairs (bp)], and the E-INS algorithm in MAFFT [31] for ITS-1 (1019 bp). Phylogenetic inference was conducted using a concatenated dataset of the two markers under two approaches: maximum likelihood (ML) analysis was performed in IQ-TREE 2.4.0 [32] and Bayesian inference (BI) was conducted in MrBayes 3.2 [33], using two independent runs of four Markov chain Monte Carlo chains for 10 million generations, sampling every 10,000 trees. The model selection and partitioning strategy for the ML analysis were determined by initially partitioning the data by gene and codon positions of cytb, and by gene only for the BI analysis, with the restriction on the number of models in ModelFinder [34] in IQ-TREE. Models and partitions selected by BIC were TIMe + G4 for cytb_pos1, F81 + F for cytb_pos2, TIM3 + F + R2 for cytb_pos3, and HKY + F + G4 for ITS-1 in the ML analysis; and GTR + F + I + G4 for cytb, and HKY + F + G4 for ITS-1 in the BI analysis. Convergence of independent BI runs was assessed by examining the SD of split frequencies, aiming for a value below 0.05 at the final generation. Additionally, a visual inspection of the sampled parameter distributions was performed using Tracer version 1.7.1 [35]. An effective sample size > 200 was considered the threshold to ensure proper mixing and reliable parameter estimates.
Clade support was assessed by Bayesian posterior probabilities (BPP) and 1000 replicates of the Shimodaira-Hasegawa approximate likelihood ratio test (SH-aLRT; [36]) and 1000 pseudoreplicates of Ultrafast Bootstrap (UFBoot; [37]) in ML.
In addition, we calculated Kimura two-parameter (K2P) distances in MEGA5 [38] to complement the phylogenetic analysis and facilitate comparisons with previous studies on triatomine speciation. The cytb gene was selected as a standard marker based on prior literature [29, 39].
Results
Taxonomy
Order Hemiptera Linnaeus, 1758
Family Reduviidae Latreille, 1807
Subfamily Triatominae Jeannel, 1919
Genus* Triatoma* Laporte, 1832
Triatoma chiarii sp. nov.
Type material
Holotype female, BRAZIL, Rio Grande do Norte, Serra Negra do Norte, Seridó Ecological Station, coordinates 06°34′37"S, 37°15′24"W. 2016. Wild environment. Number 13341, deposited in FIOCRUZ-COLVET of the regional unit of FIOCRUZ Minas Gerais. Four paratype females, same data as holotype except collection codes, 13339, 13342, 13345, 13348. Six paratype males, same data as holotype except for collection codes 13338, 13340, 13343, 13344, 13346, 13347. One paratype male, Rio Grande do Norte, Upanema, coordinates 05°38′32″ S, 37°17′27″W. 2016. Peridomestic environment 13211, deposited in FIOCRUZ-COLVET. One paratype male, Rio Grande do Norte, Messias Targino, coordinates 06°04′47″S, 37°30′42″W. Peridomestic environment. Fifth November 2017, 10021, deposited in FIOCRUZ-COLVET.
Type locality
Seridó Ecological Station, Serra Negra do Norte, Rio Grande do Norte State, coordinates 06°34′37"S, 37°15′24"W; Upanema, Rio Grande do Norte, coordinates 05°38′32″S, 37°17′27″W; and Messias Targino, Rio Grande do Norte, 06°04′47″S, 37°30′42″W (Fig. 1).
ZooBank registration
To comply with the regulations set out in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature, details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) of the article is urn:lsid:zoobank.org:pub:56EFEA36-AC0D-42B5-A83B-A73FDFA06781. The LSID for the new name Triatoma chiarii sp. nov. is urn:lsid:zoobank.org:act:49976E40-C8A6-4683-AF61-5D5F3A0AB8ED.
Etymology
The specific epithet “chiarii” was chosen to honor the distinguished career of Prof. Dr. Egler Chiari (1934–2020), who was internationally recognized for his contributions to the understanding of T. cruzi.
Bionomics
Most of the specimens were found in rock outcrops within the semiarid Caatinga biome. Some adults were found in peridomestic environments without signs of domiciliation.
Biological observations
Most specimens were found in rock outcrops, but some adults were collected in peridomestic environments, with no evidence of domiciliation. Two specimens collected prior to the formal description were tested for T. cruzi infection using optical microscopy of fecal samples and were found to be negative. However, the type series was not screened, as the intention was to preserve specimen integrity.
Description
Length of male 19.0–21.5 mm, of female 19–24 mm; pronotum width of male and female 4.0–5.0 mm. Overall color dark brown, with orange markings on pronotum, and yellowish on scutellum, hemelytra, and connexivum (Fig. 3A, B). Integument appearing glabrous, only with very sparse, short, inapparent setae. Head uniformly dark (Fig. 4A), slightly rugose and granulose, somewhat twice as long as wide across eyes (1:0.40–0.50) and longer than pronotum (1:0.75–0.80). Anteocular region 6 times as long as postocular (1:0.16–0.17), postocular with sides almost straight, distinctly converging toward behind. Genae pointed apically, distinctly surpassing the apex of the clypeus. Jugae angular apically. Eyes small, in lateral view approaching but not attaining the level of the lower and remote from the level of the upper surface of the head. Ratio width of eye to synthlipsis 1:1.8–2.0, 1:2.5. Antenniferous tubercles situated at or slightly before the middle of the anteocular region. First antennal segment appearing unusually short, extending only slightly beyond half the distance from its base to the apex of the head, viz., falling considerably short of the apex level of the clypeus. Second segment subcylindrical, with adpressed setae shorter than its diameter. Ratio of antennal segments 1:3.3–4.2:2.3–2.8:2.1–2.5. Rostrum slender, practically glabrous, except for a group of long hairs at the apex of the third segment (Fig. 4B). Ratio of rostral segments 1:2.2–2.8:1.1–1.2. First segment not attaining the level of the apex of antenniferous tubercles, second approaching the hind margin level of eyes. Neck dark, with a pair of light-colored spots laterally (Fig. 4C).Fig. 3A, BDetail of the female holotype of Triatoma chiarii sp. nov. A Dorsal view, B ventral view. (Photographs by CD)Fig. 4A–FDetail of the male of Triatoma chiarii sp. nov. A Head, B rostrum, C neck, D pronotum, E scutellum, F hemelytra. (Photographs by CD)
Pronotum very sparsely granulose, dark brown, presenting orange spots on the collar (Fig. 4D), discal callosities, on the lateral margins of the anterior lobe, submedian carinae, and on the humeral area. Anterior lobe with orange discal tubercles reduced to suboval transverse callosities, lateral tubercles absent. Posterior lobe shallowly rugose. Submedian carinae extending close to the hind margin of the sclerite. Humeri narrowly rounded, almost angular. Figure 4D shows changes in the spots and color of the pronotum. Scutellum rugose, dark brown, with a central depression. Posterior process almost entirely dark brown except in the rounded apex (Fig. 4E). Hemelytra not attaining the apex of the seventh urotergite. Corium light yellow with irregularly dark areas, the apex of the costal margin narrowly yellow. Clavus yellow with dark areas. Membrane yellowish white. Veins of membrane are narrowly dark brown; cells of membrane have a conspicuous dark spot of variable extension, centered on the vein separating cells (Fig. 4F).
Legs are uniformly dark and slender. The second pair bears two inconspicuous denticles on the femora (Fig. 5A, B). Spongy fossulae absent in both sexes.Fig. 5. Ventral views of female and male specimens of Triatoma chiarii sp. nov., showing A, B denticles on the legs, and C, D position and coloration of the abdominal spiracles, as well as the coloration the connexivum. (Photographs by CD)
Venter delicately striated transversally, sparsely setose. Venter dark brown (Fig. 5C, D), with spiracles enclosed in small yellow areas confluent with the corresponding yellow spots of the connexivum adjoining the connexival suture, which is almost imperceptible in females (Fig. 5C). Connexivum dark brown, segments on the disc with large yellowish spot occupying two-thirds of the surface of each segment, approaching but not touching posteriorly, and distant from the anterior intersegmental suture. Both dark and light areas span the entire width of the connexival segments, with the light areas shortly extending on the respective urosternite (Fig. 5C, D). Abdominal spiracles are almost imperceptible. Table 1 summarizes the described characteristics of T. chiarii sp. nov. and compares them with those of T. brasiliensis and T. petrocchiae.Table 1. Comparison of morphometric characteristics of Triatoma chiarii sp. nov., Triatoma brasiliensis and Triatoma petrocchiae**T. chiarii sp. novT. brasiliensis**T. petrocchiaeTotal length (mm)Male 19.0–21.5;Female 19.0–24.0Male 22.0–25.0;Female 23.0–25.5Male 17.0–21.5;Female 18.0–23.0Overall colorDark brown with orange markings on the pronotum and yellow markings on the scutellum, hemelytra, and connexivumDark brown to black with yellow markings on the neck, pronotum, legs, hemelytra, and connexivumDark brown with yellow markings on the pronotum, scutellum, hemelytra, and connexivumHeadUniformly dark, slightly rough and granulated, slightly more than twice as long as the width of the eyes (1:0.40–0.50), and longer than the pronotum (1:0.75–0.80)Dark brown or black, rough dorsally and laterally, distinctly longer than wide at the level of the eyes (1:0.5–0.55), and clearly longer than the pronotum (1:0.85–0.95)Black, delicately rough and granulated, about twice as long as wide at the level of the eyes (1:0.40–0.45), and much longer than the pronotum (1:0.70–0.75)Anteocular regionSix times longer than the postocular region (1:0.16–0.17)Four times longer than the postocular region (1:0.25)Four times longer than the postocular region (1:0.25)EyesIn lateral view, they approach but do not reach the lower level and are distant from the upper surface of the headIn lateral view, they do not reach the ventral surface and are distant from the dorsal surfaceIn lateral view, they do not reach the level of the ventral surface of the head and are distant from the level of the dorsal surfaceFirst antennal articleAppears unusually short, extending only slightly beyond half the distance from its base to the apex of the head, significantly below the level of the apex of the clypeusReaching the level of the apex of the clypeusShort and does not reach the level of the apex of the clypeusRostrumSlim, practically glabrous, except for a group of long hairs at the apex of the third segmentThick and as dark as the headNarrow, almost glabrous, except for a group of long bristles at the apex of the third articleProportions between rostral segments1:2.2–2.8:1.1–1.21:1.7–1.9:0.9–1.051:2,2- 2,8:1,1–1,2NeckDark with a pair of light spots on the sidesDark with a pair of light lateral spotsBlack with a pair of light lateral spotsPronotumDark brown, with orange spots on the collar, discal callosities, on the lateral margins of the anterior lobe;Submedian carinae and in the humeral region, extending near the posterior margin of the sclerite;Anterior lobe with reduced orange discal tubercles, forming transverse suboval callosities. Lateral tubercles absentPosterior lobe slightly rugoseDark brown to black with yellow spots on the collar;Lateral processes, discal tubercles of the anterior lobe, entire submedian carinae, and adjacent area to the carinae;Anterior lobe with very small discal tubercles, difficult to notice;Lateral tubercles absentDark brown collar, discal callosities, and in some cases, the lateral margins of the anterior lobe are yellowish dark brown;Anterior lobe with discal tubercles reduced to suboval callosities, and lateral tubercles absentScutellumDark brown with a central depression;Rough;Posterior process almost entirely dark brown, except for the rounded apexDark brown;Apical process as long as the main body of the scutellum, with a yellow tipDark brown;Apical process short, about three-thirds the length of the main body of the scutellumHumeral anglesNarrowly rounded, almost angularRoundedRounded, almost angularHemelytraNot reaching the apex of the seventh urotergite;Pale yellow corium with irregularly dark areas, narrowly yellow at the costal margin apex;Yellow clavus with dark areas;Whitish-yellow membrane;Narrow dark brown veins on the membranePale yellow corium with dark areas of variable extent;Entirely black clavus;Smoky yellow to light brown membrane;Black veins on the membraneReaching or almost reaching the apex of the seventh urotergite;Dark brown corium;Whitish-yellow membrane with a large central darker spotLegsUniformly dark;The second pair has two inconspicuous denticles on the femurs;Sponge fossae absent in both sexesDark with light annulation on the femoraBlack;Sponge fossae absent in both sexes
Geometric morphometrics
Hemelytra
All statistical tests revealed significant differences (P < 0.0001) between species (Table 2). PC1 and PC2 used in the analysis of this structure accounted for a total of 71.29% of the variance (PC1 = 62.4%, PC2 = 8.89%). The species are clearly separate in the factorial maps of both the PCA and canonical correlation analysis, with no overlap. Triatoma lenti is the most distant species in the first, while T. sherlocki is the most distant in the second (Additional file 2: Figure S1; Additional file 3: Figure S2).Table 2. Summary of multivariate statistical tests (multivariate ANOVA) performed in JMP software to assess morphological variation in hemelytra (wings) and heads of Triatoma speciesTestValueApproximate* FNumber of dfDenominator df**P* > FWingsWilks’ lambda0.12641057.620828275.44 < 0.0001Pillai’s trace1.34009635.685928316 < 0.0001Hotelling-Lawley trace3.58766479.587928180.47 < 0.0001Roy’s max root2.434863227.4792779 < 0.0001HeadWilks’ lambda0.007570420.875128203.33 < 0.0001Pillai’s trace2.02067588.604728236 < 0.0001Hotelling-Lawley trace26.10745951.133528130.55 < 0.0001Roy’s max root22.806483192.2261759 < 0.0001All of the tests showed statistically significant differences (P < 0.0001) between species for both structures
According to the clustering analysis, two major groups were observed in the dendrogram. The first comprises two subgroups: one includes T. chiarii sp. nov., closely related to T. melanica, and the other includes T. brasiliensis and T. juazeirensis, with T. bahiensis positioned externally to both. The second group includes T. petrocchiae, which is directly associated with T. lenti, while T. sherlocki appears as the outgroup species (Fig. 6A).Fig. 6A, BCluster analysis of Triatoma species based on hemelytra and head structures. Dendograms generated from cluster analysis based on A hemelytra and B head structures of species from the Triatoma brasiliensis complex and Triatoma chiarii sp. nov
Head
As with the hemelytra analysis, all statistical tests for head shape variation revealed significant differences between the species (P < 0.0001; Table 2). The sum of PC1 and PC2 accounted for 80.4% of the variation (PC1 = 69.9%, PC2 = 10.5%). Once again, no overlap was observed between the species in either of the factorial maps. Two closely related groups were identified, one comprising T. brasiliensis with T. juazeirensis, T. melanica and T. bahiensis; and the other one comprising T. petrocchiae together with T. chiarii sp. nov. Triatoma sherlocki and T. lenti were identified as outgroup species (Additional file 4: Figure S3; Additional file 5: Figure S4).
The dendrogram generated by the clustering analysis revealed a major group comprising a subgroup in which T. brasiliensis is closely related to T. melanica, both in close proximity to T. juazeirensis. T. chiarii sp. nov. was directly associated with T. petrocchiae, while T. sherlocki appeared as an external species. Separately, T. bahiensis was directly linked to T. lenti (Fig. 6B).
Phylogenetic reconstructions
Phylogenetic analyses (Fig. 7; Additional file 6: Figure S5) based on concatenated datasets recovered the placement of T. chiarii sp. nov. within the T. brasiliensis species complex (SH-aLRT = 92.4, UFBoot < 95, BPP = 1.00), with T. petrocchiae recovered as a sister group to the remaining species of the complex. All T. chiarii sp. nov. samples were recovered in a strongly maximally supported clade. However, its position as sister to the clade comprising T. brasiliensis, T. juazeirensis, T. sherlocki, T. bahiensis, T. lenti, and T. melanica received weak support (SH-aLRT < 80, UFBoot < 95, BPP < 0.95). Interspecific genetic distances (K2P) between T. chiarii sp. nov. and other members of the T. brasiliensis complex were consistently high, with values ranging from 0.142 to 0.146 when compared to T. brasiliensis, and 0.141 to 0.147 when compared to T. petrocchiae. These values are markedly higher than those observed between several recognized species in the complex, such as T. lenti and T. bahiensis (0.026), T. lenti and T. melanica (0.031), or T. brasiliensis and T. juazeirensis (0.077). (Additional file 7: Table S2). Phylogenetic reconstructions based on individual markers are available in Additional file 8 (Figure S6) and Additional file 9 (Figure S7).Fig. 7. Maximum likelihood tree of triatomine species based on the concatenated molecular dataset. The tree was constructed using 428 base pairs (bp) of cytochrome b (cytb) and 1019 bp of the internal transcribed spacer 1 for 38 terminals representing selected groups within the South American Triatomini. Thickened branches were also recovered in Bayesian inference analysis. Clade support values are indicated as Shimodaira-Hasegawa approximate likelihood ratio test ≥ 80% / Ultrafast Bootstrap ≥ 95% above the branches, and Bayesian posterior probabilities ≥ 0.95 below. Colors indicate members of distinct groups, complexes, and subcomplexes following the classifications proposed by the following authors: Monteiro et al. [64] (1), Alevi et al. [65] (2), Schofield and Galvão [66] (3), Belintani et al. [51] (4), and Dale et al. [10] (5)
Discussion
In this study, we describe a new species of triatomine within the genus* Triatoma* based on a combination of diverse analytical approaches. Triatoma chiarii sp. nov. morphologically exhibits a blend of traits shared with both T. petrocchiae and T. brasiliensis. Features such as the structure of the first antennal segment, a glabrous rostrum, and the absence of a spongy fossette in both sexes resemble those of T. petrocchiae. Conversely, characteristics typically associated with T. brasiliensis, including conspicuous yellow marks on the sides of the neck, a dark brown pronotum, and a scutellum with a rounded and raised posterior process, are also present [11]. However, the morphological similarity between T. chiarii sp. nov., T. brasiliensis, and T. petrocchiae, along with their overlapping geographic distribution, may complicate accurate identification and lead to potential misclassification of these taxa.
Morphological analyses play a crucial role in the description and validation of species [21, 40–42]. Coloration patterns in the early studies of the T. brasiliensis complex were particularly significant. The species currently classified by Costa et al. [21] were initially regarded as chromatic variants of T. brasiliensis [8, 11]. The status of T. b. melanica was subsequently elevated to species rank as T. melanica [43], T. juazeirensis was described [44], and T. b. macromelasoma was redescribed [21]. A morphological character analysis in 2016 was instrumental in revalidating T. bahiensis as a distinct species [45].
Although hybridization events are possible within the T. brasiliensis complex [46, 47], experimental crosses between T. brasiliensis and T. petrocchiae have yielded eggs with low or no viability, indicating the presence of reproductive barriers between these species [48, 49]. While reproductive compatibility has been observed between other species and subspecies within the T. brasiliensis complex [46, 47], further studies are needed to determine whether T. chiarii sp. nov. shares this capacity.
Geometric morphometrics of the wing and head in this study revealed that T. chiarii sp. nov. is distinctly grouped separately from T. brasiliensis and T. petrocchiae. However, it showed greater proximity to the latter, as well as to the other members of the T. brasiliensis species complex. Geometric morphometrics has been widely applied in various studies, focusing on both the wing [20, 26, 45, 50–52] and the head [9, 26, 53, 54], using different approaches. Using this methodology, Costa et al. [15] were the first to demonstrate homoploid hybrid speciation within the triatomine group, specifically in T. b. macromelasoma.
Phylogenetic reconstruction incorporating ITS-1 in addition to the widely used cytb marker placed all of the specimens of T. chiarii sp. nov. in this study within the monophyletic T. brasiliensis species complex with strong support. The sister group position of T. petrocchiae in relation to other members of the T. brasiliensis complex (T. brasiliensis, T. juazeirensis, T. sherlocki, T. bahiensis, T. lenti, and T. melanica) was previously documented by Oliveira et al. [9], also with high support, using multiple mitochondrial genes (16S rRNA, COI, cytb, and 12S rRNA). Notably, the phylogenetic relationships of T. chiarii sp. nov. in relation to the other members of the T. brasiliensis species complex, as well as the position of this complex in relation to other Triatomini species complexes, remain unclear, highlighting the need for additional molecular markers and broader taxonomic sampling to clarify their evolutionary history.
Genetic distance analyses based on variations in the cytb gene have been a key tool in delimiting species status within the Triatominae. In this context, Monteiro et al. [29] showed that distances greater than 0.075 K2P are considered sufficient to establish a newly discovered taxon as an independent evolutionary entity. Herein, we observed that all T. chiarii sp. nov. samples exhibited values approximately twice (0.15) the defined cutoff point.
According to Noireau et al. [55], the use of different methodologies is important for describing new species. A species definition cannot solely rely on morphology; it is essential to understand the phenotypic plasticity driven by different factors and environmental changes [56]. Geometric morphometrics has been employed in several studies as an effective tool for defining species, subspecies, and intraspecific variation within the subfamily Triatominae, with the wings and head being the most commonly used phenotypic markers to address these issues [20, 26, 39, 50, 52, 57, 58]. Likewise, molecular methods are equally critical for understanding speciation processes [29]. Thus, combining both approaches is essential for ensuring the robustness of the results [9]. Based on the results described herein, we conclude that T. chiarii sp. nov. is indeed a new species of triatomine and can be classified within the T. brasiliensis species complex.
Conclusions
Triatoma chiarii sp. nov. was captured in sylvatic and peridomestic environments in regions where other triatomine species are present. Triatoma brasiliensis, T. pseudomaculata, T. petrocchiae, Panstrongylus lutzi (Neiva and Pinto, 1923), Panstrongylus megistus (Burmeister, 1835), Rhodnius nasutus Stål, 1859, and Psammolestes tertius Lent and Jurberg, 1965 have already been identified in the investigated areas [59–63]. The description of a new species from these areas highlights the importance of ongoing vector monitoring actions conducted by those involved in Chagas disease surveillance, as, given the high morphological similarity between other triatomine species present in the region, these actions ensure the accurate identification of triatomine species and thereby enable a timely and appropriate epidemiological response when necessary. Moreover, further investigation into other eco-epidemiological aspects related to Triatoma chiarii sp. nov., such as the prevalence of natural Trypanosoma cruzi infection, feeding sources, and its potential for infesting human dwellings, is crucial.
Supplementary Information
Additional file 1.Additional file 2.Additional file 3.Additional file 4.Additional file 5Additional file 6.Additional file 7.Additional file 8.Additional file 9.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO. Chagas disease (American trypanosomiasis). 2020. https://www.who.int/chagas/disease/en/. Accessed 20 Aug 2023.
- 2Coutinho CBD. Taxonomia integrada de espécies de Triatoma Laporte, 1832 (Hemiptera: Reduviidae: Triatominae) do Estado do Rio Grande do Sul, Brasil. https://www.arca.fiocruz.br/handle/icict/30228 (2017). Accessed 25 Nov 2024.
- 3Rambaut A, Drummond AJ. Tracer version 1.7.1. 2009. http://tree.bio.ed.ac.uk/software/tracer/. Accessed 12 Sept 2024.