Dual‐Pathway Defense: Ultramicro‐Pulverised Powder of Polygonum chinense L. and Atractylodes rhizome (PAUP) Rescues Broilers From E. coli‐Triggered Liver Injury Through Modulation of Oxidative Stress and Inflammation
Jia‐Ci Cai, Yan‐Na Guo, Shao‐Shan Liang, Yan Liu, Fu‐Qiang Huang, Qi‐Peng Lv, Lan‐Yi Zhang, Yi Qin, Xiao‐Jing Chen, Yu‐Xin Liang, Yong‐Ming He, Lu‐Ping Tang

TL;DR
This study shows that a natural powder (PAUP) protects chickens from E. coli-induced liver damage by reducing inflammation and oxidative stress, offering a safer alternative to antibiotics.
Contribution
PAUP is shown to mitigate E. coli-induced liver injury through dual pathways of antioxidant activation and inflammation suppression in broilers.
Findings
PAUP reduced bacterial load, liver damage, and inflammation in E. coli-infected broilers.
PAUP restored redox balance via Nrf2-HO-1 pathway activation and suppressed inflammation through TLR4/NF-κB inhibition.
PAUP improved weight gain, feed efficiency, and liver function, offering a natural alternative to antibiotics.
Abstract
This study characterizes the composition and blood components of ultramicro‐pulverized Polygonum chinense L. and Atractylodes rhizome powder (PAUP) and investigates its protective mechanisms against Escherichia coli ( E. coli )‐induced liver injury in broilers. PAUP constituents and serum metabolites were profiled by ultra‐high‐performance liquid chromatography‐mass spectrometry (UPLC‐MS). Fourteen‐day‐old broilers were randomly divided into 6 groups: control, E. coli model, gentamicin, and PAUP high/medium/low‐dose groups. Broilers were infected with 1010 CFU/mL E. coli O157:H7 via intraperitoneal injection. PAUP was administered 4 h post‐ E. coli infection for 7 days by gavage. Then, serum and liver samples from half of the chicks in each group were collected for further analysis. Recovery groups were analyzed after 7 additional days. UPLC‐MS identified 718 PAUP components,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
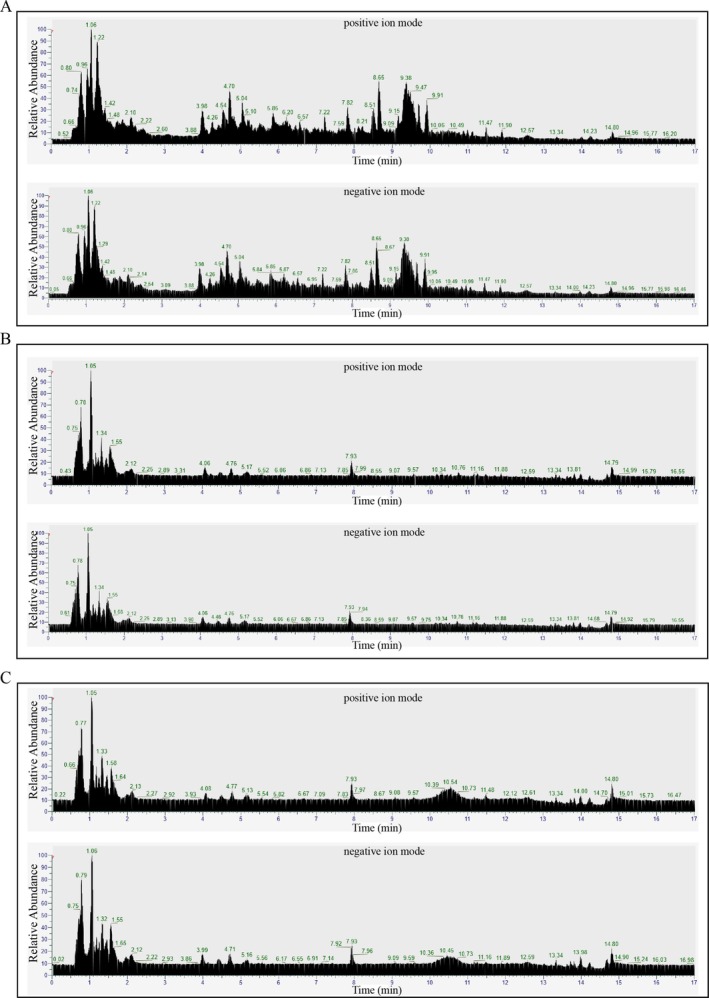 FIGURE 1
FIGURE 1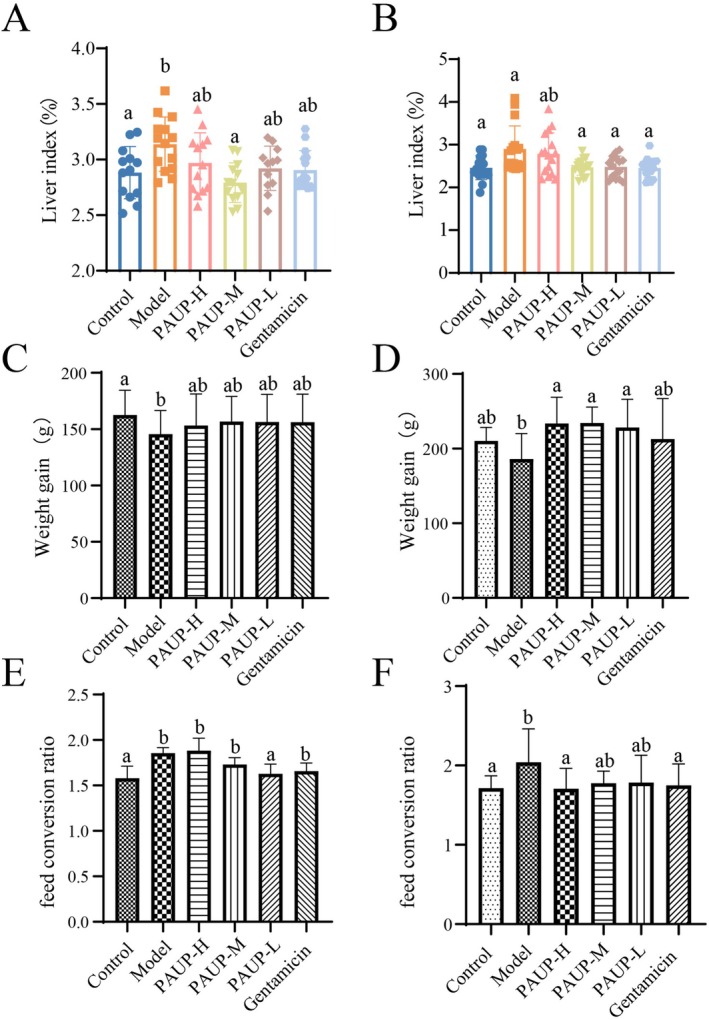 FIGURE 2
FIGURE 2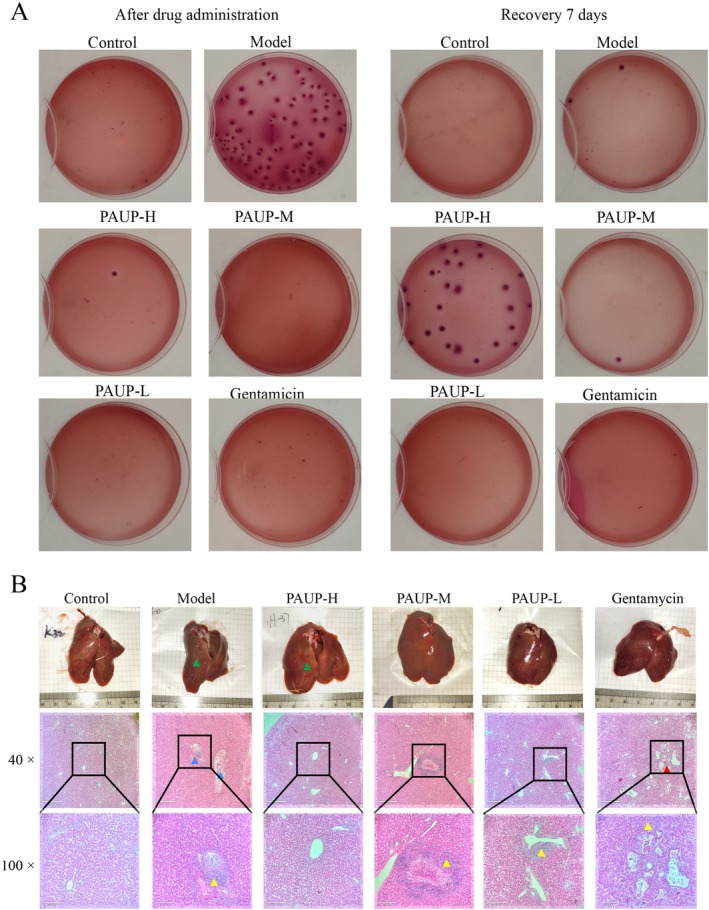 FIGURE 3
FIGURE 3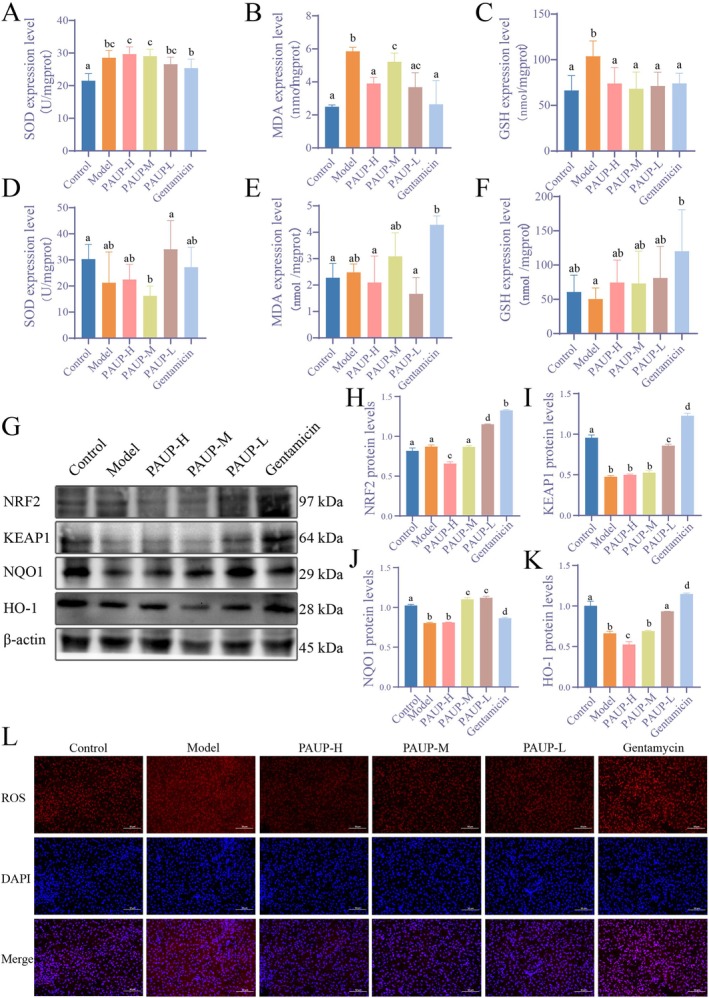 FIGURE 4
FIGURE 4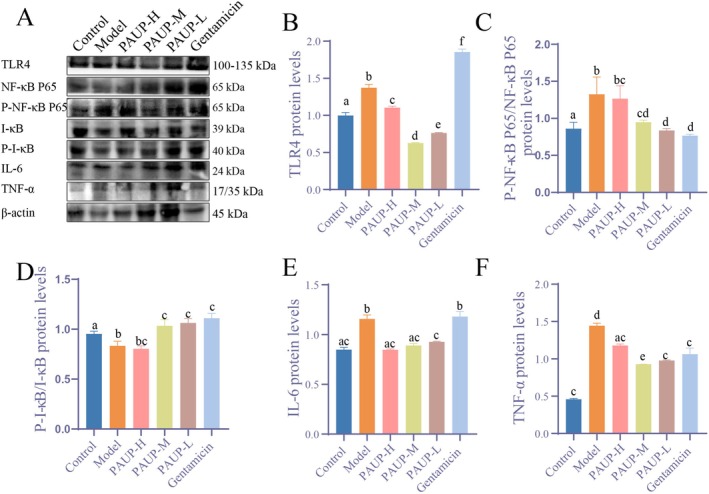 FIGURE 5
FIGURE 5| Order | Metabolite | Formula | Retention time (min) | Proportion (%) |
|---|---|---|---|---|
| 1 | Hesperidin | C28H34O15 | 6.224 | 8.840 |
| 2 | Citrate | C6H8O7 | 1.220 | 7.007 |
| 3 | 3‐O‐Caffeoylquinic acid | C16H18O9 | 4.285 | 6.385 |
| 4 | 2‐(9‐Decenyl) glutaconic acid | C15H24O4 | 9.692 | 5.281 |
| 5 | Narirutin | C27H32O14 | 6.001 | 5.023 |
| 6 | Ginnalin B | C13H16O9 | 2.127 | 4.442 |
| 7 | 3,5‐Dicaffeoylquinic acid | C25H24O12 | 6.165 | 4.328 |
| 8 | Olivetol | C11H16O2 | 7.222 | 3.309 |
| 9 | Phenylalanin | C9H11NO2 | 2.127 | 2.941 |
| 10 | Chebulagic acid | C41H30O27 | 5.441 | 2.467 |
| 11 | Didymin | C28H34O14 | 7.015 | 2.205 |
| 12 | Cumanin | C15H22O4 | 8.866 | 1.814 |
| 13 | Isomaltotriose | C18H32O16 | 0.819 | 1.667 |
| 14 | Geraniin | C41H28O27 | 5.049 | 1.574 |
| 15 | Chebulanin | C27H24O19 | 4.988 | 1.427 |
| 16 | Epicatechin gallate | C22H18O10 | 5.883 | 1.268 |
| 17 | (L)‐Dehydroascorbic acid | C6H6O6 | 1.293 | 1.261 |
| 18 | Beta‐Glucogallin | C13H16O10 | 1.424 | 1.232 |
| 19 | Neoschaftoside | C26H28O14 | 5.260 | 1.218 |
| 20 | Oleuropeinic acid | C25H30O15 | 5.776 | 1.018 |
| 21 | Orientin | C21H20O11 | 5.343 | 0.998 |
| 22 | Di‐O‐methylbergenin | C16H20O9 | 5.224 | 0.854 |
| 23 | 1,6‐anhydro‐b‐D‐Glucose | C6H10O5 | 0.731 | 0.822 |
| 24 | L‐Asparagine | C4H8N2O3 | 0.761 | 0.742 |
| 25 | 1,3‐Dicaffeoylquinic acid | C25H24O12 | 5.120 | 0.724 |
| 26 | Vincosamide | C26H30N2O8 | 0.839 | 0.724 |
| 27 | Cis‐1,2‐Dihydroxycyclohexane | C6H12O2 | 1.744 | 0.687 |
| 28 | Azelaic acid | C9H16O4 | 6.552 | 0.686 |
| 29 | Linoleic acid | C18H32O2 | 13.995 | 0.643 |
| 30 | 3‐hydroxy‐4‐E‐Hexenoic acid | C6H10O3 | 4.554 | 0.587 |
| Order | Metabolite | Formula | Retention time(min) | Proportion (%) |
|---|---|---|---|---|
| 1 | Atractylenolide III | C15H20O3 | 9.847 | 29.989 |
| 2 | L‐Proline | C5H9NO2 | 0.803 | 11.324 |
| 3 | Parthenolide | C15H20O3 | 9.300 | 4.451 |
| 4 | Neochlorogenic acid | C16H18O9 | 4.653 | 4.264 |
| 5 | Tangeretin | C20H20O7 | 9.869 | 4.211 |
| 6 | Lathosterol | C27H46O | 12.225 | 2.210 |
| 7 | Byzantionoside B | C19H32O7 | 6.160 | 2.044 |
| 8 | Vicenin 3 | C26H28O14 | 5.189 | 1.622 |
| 9 | Isoorientin | C21H20O11 | 5.271 | 1.549 |
| 10 | 1‐Linoleoyl‐sn‐glycero‐3‐phosphorylcholine | C26H50NO7P | 10.767 | 1.405 |
| 11 | Oxymatrine | C15H24N2O2 | 2.264 | 1.314 |
| 12 | Curcumadione | C15H22O2 | 9.177 | 1.106 |
| 13 | Aurantiamide acetate | C27H28N2O4 | 10.029 | 1.098 |
| 14 | Naringenin | C15H12O5 | 5.919 | 0.940 |
| 15 | Atractylenolide I | C15H18O2 | 11.659 | 0.911 |
| 16 | Flavinantine | C19H21NO4 | 8.166 | 0.884 |
| 17 | 6‐Demethoxytangeretin | C19H18O6 | 8.776 | 0.876 |
| 18 | Isosinensetin | C20H20O7 | 8.285 | 0.863 |
| 19 | Mamanine | C15H22N2O2 | 2.124 | 0.783 |
| 20 | 1,3‐Di‐O‐caffeoylquinic aicd | C25H24O12 | 6.093 | 0.779 |
| 21 | Sinensetin | C20H20O7 | 8.812 | 0.759 |
| 22 | Isosakuranetin | C16H14O5 | 6.948 | 0.696 |
| 23 | Eriodictiol‐7‐glucoside | C22H24O11 | 6.138 | 0.649 |
| 24 | Kurarinone | C26H30O6 | 9.276 | 0.629 |
| 25 | Salviolone | C18H20O2 | 10.587 | 0.600 |
| 26 | Specnuezhenide | C31H42O17 | 5.835 | 0.591 |
| 27 | Isovitexin | C21H20O10 | 5.650 | 0.578 |
| 28 | Neoisoliquiritin | C21H22O9 | 5.919 | 0.545 |
| 29 | Icariside B1 | C19H30O8 | 6.354 | 0.522 |
| 30 | Excavatin M | C19H20O7 | 9.363 | 0.505 |
| Order | Metabolite | Formula | Order | Metabolite | Formula |
|---|---|---|---|---|---|
| 1 | Icosanedioic acid | C20H38O4 | 66 | Obscuraminol F | C16H33NO |
| 2 | Linoleic acid | C18H32O2 | 67 | Marumoside A | C14H19NO6 |
| 3 | N‐Acetyl‐L‐ornithine | C7H14N2O3 | 68 | (+)‐Junenol | C15H26O |
| 4 | Cyclo(Ile‐Leu) | C12H22N2O2 | 69 | (2S,3R,4E)‐2‐Amino‐4‐heptadecene‐1,3‐diol | C17H35NO2 |
| 5 | cis‐1,2‐Dihydroxycyclohexane | C6H12O2 | 70 | n‐Docosanol | C22H46O |
| 6 | L‐Tyrosine | C9H11NO3 | 71 | Stearic acid amide | C18H37NO |
| 7 | Isomaltotriose | C18H32O16 | 72 | Obscuraminol B | C16H31NO |
| 8 | Ganoderal A | C30H44O2 | 73 | Steviol‐19‐O‐Glucoside | C12H19N3O5 |
| 9 | Methyl eugenol | C11H14O2 | 74 | Triptobenzene H | C21H28O4 |
| 10 | Abscisic Acid | C15H20O4 | 75 | 13,16,19‐Docosatrienoic acid | C22H38O2 |
| 11 | Pelletierine | C8H15NO | 76 | D‐proline | C5H9NO2 |
| 12 | L‐Asparagine | C4H8N2O3 | 77 | Salirepin | C13H18O8 |
| 13 | Bufalin | C24H34O4 | 78 | Atractylodin | C13H10O |
| 14 | (R)‐a‐Terpinyl b‐D‐glucoside | C16H28O6 | 79 | Betonicine | C7H13NO3 |
| 15 | Furfuryl acetate | C7H8O3 | 80 | 9,10‐Methylenehexadecanoic acid | C17H32O2 |
| 16 | Undecylenic acid | C11H20O2 | 81 | Lycernuic ketone B | C31H48O6 |
| 17 | 1,11‐Undecanedicarboxylic acid | C13H24O4 | 82 | Isoorientin | C21H20O11 |
| 18 | Olivetol | C11H16O2 | 83 | Tangeretin | C20H20O7 |
| 19 | D‐Glucosamine | C6H13NO5 | 84 | 11‐Hydroxydrim‐7‐en‐6‐one | C15H24O2 |
| 20 | 4‐Hydroxyisoleucine | C6H13NO3 | 85 | 2,3,24‐Trihydroxy‐12‐ursen‐28‐oic acid | C30H48O5 |
| 21 | Orientin‐2″‐O‐p‐trans‐coumarate | C30H26O13 | 86 | Longicaulenone | C12H18O4 |
| 22 | Chrysin‐7‐O‐glucuronide | C21H18O10 | 87 | Calycosin | C16H12O5 |
| 23 | Quinic acid | C7H12O6 | 88 | Cholinesulfuric acid. | C5H13NO4S |
| 24 | 2‐(9‐Decenyl)glutaconic acid | C15H24O4 | 89 | 6,9,10‐Trihydroxy‐7‐megastigmen‐3‐one | C13H22O4 |
| 25 | Neoagarobiose | C12H20O10 | 90 | 6‐O‐Ethyltetradymodiol | C17H26O3 |
| 26 | Caffeic acid | C9H8O4 | 91 | Coniferaldehyde | C10H10O3 |
| 27 | Shikimic acid | C7H10O5 | 92 | Curcumol | C15H24O2 |
| 28 | 2‐O‐alpha‐D‐Glucopyranosyl‐L‐ascorbic acid | C12H18O11 | 93 | Geranylacetone | C13H22O |
| 29 | 4‐Hydroxy‐1,10‐secocadin‐5‐ene‐1,10‐dione | C15H24O3 | 94 | Kaempferol | C15H10O6 |
| 30 | Gamma‐Aminobutyric acid | C4H9NO2 | 95 | 6‐Shogaol | C17H24O3 |
| 31 | Gentisic acid | C7H6O4 | 96 | Cyclo(alanylleucyl) | C9H16N2O2 |
| 32 | Isomangiferin | C19H18O11 | 97 | Panowamycin B | C17H26O3 |
| 33 | 3‐hydroxy‐4‐E‐Hexenoic acid | C6H10O3 | 98 | 7,8‐Dihydroxyflavone | C15H10O4 |
| 34 | Trans‐2,3‐Dihydro‐3‐ethoxyeuparin | C15H18O4 | 99 | 5‐Methoxyindoleacetate | C11H10NO3 |
| 35 | Methyl Kakuol | C11H12O4 | 100 | LINAMARIN | C10H17NO6 |
| 36 | Vanillic acid | C8H8O4 | 101 | Cucumegastigmane I | C13H20O4 |
| 37 | 6‐Hydroxy‐7‐methoxydihydroligustilide | C13H18O4 | 102 | 15‐Dihydroepioxylubimin | C15H26O3 |
| 38 | N‐Acetyltryptamine | C12H14N2O | 103 | GeranylTiglate | C15H24O2 |
| 39 | Aquilegiolide | C8H8O3 | 104 | linolenic acid | C18H30O2 |
| 40 | Umbelliferone | C9H6O3 | 105 | Rhodiocyanoside A | C11H17NO6 |
| 41 | Araliadiol | C15H20O2 | 106 | Perillene | C10H14O |
| 42 | Baicalin | C21H18O11 | 107 | Lysofungin | C27H49O12P |
| 43 | Sanggenol P | C30H36O6 | 108 | (+)‐Perillaldehyde | C10H14O |
| 44 | Opuntiol | C7H8O4 | 109 | Isoboonein | C9H14O3 |
| 45 | 3,4‐O‐Isopropylidene shikimic acid | C10H14O5 | 110 | Homocitrulline | C7H15N3O3 |
| 46 | Hymecromone | C10H8O3 | 111 | Isoferulic acid | C10H10O4 |
| 47 | 3‐(hydroxymethyl)cyclopentanone | C6H10O2 | 112 | 6‐Demethoxytangeretin | C19H18O6 |
| 48 | 4‐Hydroxycinnamamide | C9H9NO2 | 113 | 1,3‐Di‐O‐caffeoylquinic aicd | C25H24O12 |
| 49 | (−)‐Gallocatechin | C15H14O7 | 114 | Trolline | C12H13NO3 |
| 50 | Beta‐Glucogallin | C13H16O10 | 115 | Betaine | C5H11NO2 |
| 51 | Avicularin | C20H18O11 | 116 | Senkyunolide I | C12H16O4 |
| 52 | Vanillin | C8H8O3 | 117 | Atractylenolide I | C15H18O2 |
| 53 | Ethyl gallate | C9H10O5 | 118 | Excavatin M | C19H20O7 |
| 54 | Griffonilide | C8H8O4 | 119 | Prunasin acid | C14H18O8 |
| 55 | Myriocin | C21H39NO6 | 120 | 4‐Methylumbelliferone | C10H8O3 |
| 56 | Gallic acid | C7H6O5 | 121 | Protocatechualdehyde | C7H6O3 |
| 57 | Ethyl 2,4,6‐trihydroxybenzoate | C9H10O5 | 122 | Shizukanolide C | C15H18O3 |
| 58 | 3‐O‐Caffeoylquinic acid | C16H18O9 | 123 | Songoramine | C22H29NO3 |
| 59 | Shizukanolide H | C17H20O5 | 124 | Benzyl glucopyranoside | C13H18O6 |
| 60 | Esculetin | C9H6O4 | 125 | Neridienone B | C21H28O4 |
| 61 | Loganetin | C11H16O5 | 126 | Norpterosin B | C13H16O2 |
| 62 | 15‐Nor‐14‐oxolabda‐8(17),12‐dien‐18‐oic acid | C19H28O3 | 127 | Alpha‐Cembrenediol | C20H34O2 |
| 63 | Brevifolincarboxylic acid | C13H8O8 | 128 | Cis‐3,4‐Dihydroxy‐beta‐ionone | C13H20O3 |
| 64 | Diosmetin | C16H12O6 | 129 | 2,2,5,7,8‐Pentamethyl‐6‐Chromanol | C14H20O2 |
| 65 | DL‐Lysine | C6H14N2O2 | 130 | 3‐Butylidenephthalide | C12H12O2 |
| Control | Model | PAUP‐H | PAUP‐M | PAUP‐L | Gentamicin | Recovery control | Recovery model | Recovery PAUP‐H | Recovery PAUP‐M | Recovery PAUP‐L | Recovery gentaminic | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ALB | 13.59 ± 1.38abc | 13.99 ± 0.94bc | 13.29 ± 1.09abc | 13.01 ± 1.09ab | 12.84 ± 1.40a | 14.17 ± 1.69c | 13.16 ± 1.18a | 14.92 ± 1.71b | 13.31 ± 1.57a | 13.37 ± 1.48a | 13.38 ± 1.37a | 13.04 ± 1.77a |
| ALP | 2352.75 ± 775.57a | 3351.52 ± 1590.18ab | 3439.22 ± 2023.33ab | 3455.68 ± 2001.53ab | 3983.40 ± 1752.95b | 4286.78 ± 1913.78b | 3127.04 ± 2021.36a | 2419.95 ± 1437.65a | 3420.24 ± 1718.76ab | 4440.20 ± 1413.56b | 3125.94 ± 1709.21a | 3062.57 ± 2203.74a |
| ALT | 2.17 ± 0.78ac | 2.91 ± 0.61bc | 2.40 ± 0.73ab | 2.67 ± 0.56abc | 2.59 ± 0.70abc | 3.16 ± 0.77c | 2.68 ± 0.76a | 2.90 ± 0.78a | 2.84 ± 0.74a | 2.56 ± 0.96a | 3.22 ± 0.70a | 2.94 ± 0.59a |
| AST | 222.12 ± 23.32a | 290.10 ± 97.14b | 213.95 ± 18.77a | 225.73 ± 29.66a | 222.18 ± 26.83a | 215.16 ± 22.90a | 202.49 ± 20.84acd | 206.32 ± 20.56acd | 187.18 ± 21.00b | 208.05 ± 14.76c | 193.51 ± 17.13d | 185.45 ± 17.94b |
| TBA | 3.37 ± 1.17a | 5.24 ± 2.40b | 3.25 ± 0.79a | 3.15 ± 0.71a | 3.03 ± 0.67a | 3.73 ± 1.83a | 4.97 ± 1.87acd | 3.51 ± 1.28bc | 5.00 ± 2.60acd | 4.19 ± 1.34c | 5.73 ± 2.75d | 4.10 ± 1.47ab |
| T‐BIL | 18.59 ± 2.94a | 21.06 ± 2.89a | 18.87 ± 3.34a | 19.38 ± 3.64a | 19.96 ± 2.83a | 23.71 ± 3.32b | 5.52 ± 1.58ab | 3.29 ± 1.15b | 4.82 ± 2.90ab | 5.52 ± 2.23a | 5.91 ± 1.88a | 5.35 ± 1.98ab |
| TC | 3.56 ± 0.39a | 3.84 ± 0.41a | 3.60 ± 0.25a | 3.51 ± 0.34a | 3.59 ± 0.35a | 4.01 ± 0.37b | 3.49 ± 0.42a | 3.31 ± 0.34a | 3.43 ± 0.44a | 3.53 ± 0.37a | 3.47 ± 0.51a | 3.53 ± 0.35a |
| TG | 0.31 ± 0.06a | 0.30 ± 0.03a | 0.34 ± 0.05ab | 0.32 ± 0.06a | 0.34 ± 0.04ab | 0.31 ± 0.05b | 0.31 ± 0.07a | 0.30 ± 0.09a | 0.31 ± 0.08a | 0.29 ± 0.06a | 0.31 ± 0.07a | 0.32 ± 0.07a |
- —National Natural Science Foundation of China10.13039/501100001809
- —Guangdong Provincial Department of Science and Technology10.13039/501100007162
- —Department of Education of Guangdong Province10.13039/501100010226
- —Science and Technology Projects of Xizang Autonomous Region, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytochemistry and biological activity of medicinal plants · Phytochemical Studies and Bioactivities · Food Science and Nutritional Studies
Introduction
1
Escherichia coli ( E. coli ) infection in broilers represents a significant threat to poultry health and food safety. As a major etiological agent of colibacillosis, septicemia, and hepatic or intestinal pathology, E. coli infection compromises flock performance, leading to reduced growth rates, increased mortality, and impaired productivity (Narasinakuppe Krishnegowda et al. 2022; Islam et al. 2023). These outcomes not only cause substantial economic losses but also heighten concerns related to the microbiological safety of poultry products entering the food chain. Pathogenic E. coli strains in poultry may contribute to carcass contamination, increasing the risk of foodborne illness in humans and posing a public health challenge. Furthermore, the persistence of infection and associated antimicrobial treatments can influence antimicrobial resistance dissemination, ultimately affecting human health and quality of life. Therefore, effective control and therapeutic strategies for E. coli in broilers are essential to safeguard animal welfare, ensure food safety, and protect public health.
The liver is a primary target organ affected by E. coli . Following entry into the bloodstream, E. coli can disseminate systemically, reaching organs such as the liver and resulting in hepatic inflammation, hepatomegaly, and abscess formation (Shen et al. 2023). However, research on E. coli ‐induced liver damage in broilers remains scare, and its underlying mechanisms are incompletely understood. Therefore, thoroughly elucidating the biological mechanism of liver damage in poultry during E. coli infection, along with identifying potential targets for prevention and treatment, provides critical guidance for developing drugs against avian colibacillosis. Currently, antibacterial drugs and vaccines are the primary effective treatments for E. coli infections. However, antibacterial drugs carry significant drawbacks, including variable side effects, resistance development, and drug residues, which pose serious public health risks (Barbarossa et al. 2022). Vaccines, meanwhile, lack broad efficacy due to the high genetic diversity and rapid mutation rate of E. coli strains (Pokharel et al. 2023). These limitations underscore the urgent need for alternative strategies to manage E. coli infections in poultry. Traditional Chinese medicine (TCM), with its lower residues and reduced side effects (Li et al. 2022), has gained attention in recent years as a potential alternative to conventional antibacterial agents.
Ultrafine powder technology is a cutting‐edge process that can make Chinese medicine into a powder with a particle size below 10 μm. It enhances the bioavailability of active ingredients, promotes the release of otherwise difficult‐to‐extract compounds, and improves the homogeneity and dissolution rate of powdered TCM products. Ultrafine powders can improve the stability and consistency of herbal formulations, ensuring more precise dosing and enhanced therapeutic outcomes (Zhi et al. 2019). Research on the application of TCM superfine powders in livestock and poultry is advancing. Research found that dietary rosemary ultrafine powder supplementation can enhance the health and productivity of aged hens (Li et al. 2024). Stems and leaves in ultrafined powder of Astragalus can boost the immune response to prevent Newcastle disease infections in poultry farms (Xi et al. 2014). Various studies will investigate their use as feed additives for purposes such as growth promotion, immune modulation, and enhancement of gut health, so as to explore their promising potential as natural alternatives or supplements (Guo et al. 2025).
Atractylodes macrocephala koidz (Baizhu) has been incorporated into traditional diets and medicinal cuisines for centuries, often used in porridges, soups, and tonics to strengthen the spleen and improve digestion. Studies show that its rhizomes and polysaccharides enhance metabolic status, strengthen immune responses, and reduce diarrhea and infection risks in animals (Lu et al. 2025; Rashidah et al. 2023). Its long history in both food therapy and herbal formulas highlights its role as a bridge between nourishment and medicinal health support. Polygoni chinensis L. is the dry ground part of Polygonaceae plant, and has the functions of soothing liver and improving eyes, clearing heat and dampness, cooling blood and detoxifying. It is widely used in the treatment of conditions such as dysentery, diarrhea and hepatitis (Hossen et al. 2015; Zeng et al. 2022). Modern pharmacological studies have demonstrated that Polygoni chinensis exhibits antibacterial and anti‐inflammatory effects, particularly in relation to liver health (OuYang et al. 2012). Our precious study has found that Polygoni chinensis L. upregulates IFN production to alleviate liver damage in mice caused by Salmonella typhimurium infection (Shen et al. 2022). Polygoni chinensis L. Capsules have a significant beneficial effect on liver fibrosis induced by carbon tetrachloride in rats (Dong et al. 2021). The two herbs can both be used to treat liver damage and colibacillosis. However, it is not clear whether the Ultramicro‐pulverized powder of Polygoni chinensis L. and Atractylodes macrocephala koidz can alleviate colibacillosis and its liver damage. Ultrafine powder of Polygoni chinensis L. and Atractylodes macrocephala Koidz. were mixed in a ratio of 1:4, which was named as PAUP. This study aims to investigate the effects of PAUP on liver damage in broilers with colibacillosis.
Materials and Methods
2
Bacteria
2.1
The E. coli O157:H7 strain was originally obtained from Guangdong Microbial Culture Collection Center (GDMCC NO:1.1869, Guangdong, China). The strain was inoculated onto lysogeny broth (LB) agar plate using an inoculation loop and cultured upside down in a constant temperature incubator at 37°C for 24 h. A single colony was then transferred to 15 mL of LB broth using an inoculation loop and incubated in a shaking incubator at 37°C and 220 rpm/min for 12 h to activate the strain. After activation, the culture was streaked on an agar plate and incubated upside down at 37°C for 12 h. Single colonies were selected and transferred to 15 mL of LB broth, incubated in a shaking incubator at 37°C and 220 rpm/min for 12 h. The culture was centrifuged at 5000 rpm/min for 10 min, and the bacteria were collected. The bacteria were washed with sterile PBS buffer and then re‐suspended it in sterile PBS buffer to obtain 10^10^ CFU/mL of E. coli O157:H7 bacterial suspension for further use.
Plant Material
2.2
Ultrafine powder technology was used to obtain the ultrafine powder of Polygoni chinensis L. and Atractylodes macrocephala Koidz. The two powders were then mixed in a ratio of 1:4 (PAUP), and distilled water was added to prepare a suspension. The final suspension concentrations were 150, 75, and 37.5 mg/mL, respectively.
Animals and Experimental Design
2.3
A total of 216 one‐day‐old Nanhai Mahuang chickens were raised to 14 days of age. Healthy broilers with comparable body weights were randomly divided into 6 groups: control, model, gentamicin (20 mg/kg), high‐dose PAUP (PAUP‐H, 1500 mg/kg), medium‐dose PAUP (PAUP‐M, 750 mg/kg), and low‐dose PAUP (PAUP‐L, 375 mg/kg). Each group consisted of 6 replicates with 6 birds per replicate. Broilers were infected with 1 mL of E. coli O157:H7 (10^10^ CFUs/chick) by intraperitoneal injection. Then, PAUP and gentamicin were subsequently administered by oral gavage for 7 continuous days, beginning 4 h post‐infection. Eighteen chicks from each group were anesthetized for blood collection via the jugular vein for serological assays and analysis of drug constituents. The remaining broilers were allowed to recover for an additional 7 days after treatment, after which liver samples were collected for organ index determination, H&E staining, and other analyses. Throughout the experiment, feed intake, body weight, clinical symptoms, and feces characteristics were recorded. The feed conversion ratio (FCR) and organ index of broilers were calculated using the formulas: FCR = Feed intake/weight gain. Organ index = (organ weight/broiler weight) × 100%. All broilers were raised with ordinary feeding; the composition and nutrient levels of the experimental diets were referred to previous article (Guo et al. 2024).
Serological Test
2.4
Serum samples were collected, and the concentrations of alkaline phosphatase (ALP, 140323004), albumin (ALB, 148322010), alanine aminotransferase (ALT, 140123007), aspartate aminotransferase (AST, 140223007), total bilirubin (T‐BilirV, 140,623,005), triglyceride (TG, 141723003), total cholesterol (TC, 2285099), total protein (TP, 140821003), and total bile acid (TBA, 143223006) were quantified using commercial assay kits following the manufacturer's instructions. All kits and the biochemical analyzer (BS‐240VET) were supplied by Shenzhen Mindray Bio‐Medical Electronics Co. Ltd. (Mindray, Shenzhen, China).
UPLC‐MS
2.5
50 mg of PAUP was transferred into a centrifuge tube, and 1 mL of extraction solution (water/acetonitrile/isopropyl alcohol, 1:1:1, v/v/v) was added to prepare the PAUP solution for UPLC‐MS analysis. Plasmam samples (0.5 mL) from broilers in the normal group and the PAUP‐treated group (1.5 g/kg) were collected after 7 days of drug administration. Extraction solvent (methanol/acetonitrile, 1: 1, v/v) was added to obtain plasmam PAUP extracts. All samples were sonicated at low temperature for 30 min and centrifuged at 12000 rpm for 10 min at 4°C. The resulting supernatants were stored at −20°C for 1 h to precipitate proteins, and then centrifuged again at 12000 rpm for 10 min at 4°C. The supernatant was vacuum‐dried and redissolved in 200 μL of 50% acetonitrile solution. After swirling, the samples were centrifuged at 14000 rpm for 15 min at 4°C, and the final supernatant was collected for analysis using a UPLC‐Q Exactive HFX system (Thermo, USA).
Bacterial Load
2.6
Hepatic bacterial load was quantified as follows: approximately 0.05 g of liver tissue was homogenized in 1 mL of sterile normal saline. Then, 100 μL of the homogenate was mixed with MacConkey agar plates and incubated at 37°C for 12 h to assess bacterial growth.
Tissue Histopathological Changes Examined by H&E Staining
2.7
The liver tissues were fixed in 4% paraformaldehyde for 24 h, and then subjected to hematoxylin and eosin (H&E) staining. Histopathological changes were examined using a light microscope (RVL‐100‐G, ECHO, America).
The Contents of GSH, MDA and SOD
2.8
A 10% liver tissue homogenate was prepared by adding 0.1 g of liver tissue to 0.9 mL of saline and homogenizing thoroughly. The homogenate was centrifuged at 2000 rpm for 10 min, and the supernatant was collected for biochemical analysis. Levels of malondialdehyde (MDA, 20220622), superoxide dismutase (SOD, 20230313), and glutathione (GSH, 20221028) were measured using commercial assay kits from Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
Western Blot Analysis
2.9
Total protein was extracted from chicken liver using RIPA lysis buffer (1:100) supplemented with phenylmethylsulfonyl fluoroprotease inhibitor (PMSF). The proteins were separated by SDS–PAGE, transferred onto 0.22 μm PVDF membranes at low temperature, and blocked with 5% skim milk for 1 h at room temperature. Membranes were incubated for 3 h at room temperature with primary antibodies against TLR4 (#14358), NF‐κB (#8242), P‐NF‐κB (#3033), I‐κB (#4814), P‐I‐κB (#2859), IL‐6 (#16912), TNF‐α (#3707), Nrf2 (#12721), KEAP1 (#4678), NQO1 (#62262), HO‐1 (#86806), and β‐actin (#4970). After two 15 min washes with TBST, membranes were incubated with HRP‐conjugated sheep anti‐rabbit secondary antibody (SA00001‐2) for 1 h at room temperature, followed by two washes with TBST. Protein signals were visualized using ECLchemiluminescence, and band intensities were quantified using Image J software. All antibodies were obtained from Cell Signaling Technology (Massachusetts, American).
ROS Measurement
2.10
The frozen liver sections were gently air‐dried before adding ROS dye solution, followed by incubation at 37°C for 30 min in the dark. The sections were washed 3 times with PBS by shaking for 5 min each time. After slight drying, DAPI staining solution was applied and incubated at room temperature for 10 min. The sections were then dried again, mounted with anti‐fade medium, and examined using a fluorescence microscope (OLYMPUS IX53, Japan).
Statistical Analysis
2.11
Quantitative data are expressed as mean ± standard deviation. Statistical analysis and charting were performed using GraphPad Prism 8 (San Diego, CA, USA). Group differences were evaluated by one‐way nonparametric analysis of variance (ANOVA), and values of p < 0.05 were considered statistically significant.
Results
3
Analysis of Chemical Components of PAUP and Its Absorption Into Blood by UPLC‐MS
3.1
As shown in Figure 1, a total of 718 components were detected in PAUP, including lipids, flavonoids, acids, oxides, and related metabolites. The top 30 compounds identified in the negative (NEG) and positive (POS) modes are listed in Tables 1 and 2, respectively. These major constituents included atractylenolide III, hesperidin, alpha‐lactose, D‐proline, citrate, and 3‐O‐caffeoylquinic acid, among others. Furthermore, comparative analysis of plasma samples from the control and PAUP‐H groups revealed that, compared to the control, 130 metabolites were significantly elevated in the PAUP‐H group (Table 3). These elevated metabolites included atractylenolide I, linoleic acid, araliadiol, baicalin, gallocatechin, and atractylodin.
Chemical constituents of PAUP and blood analyzed by UPLC‐MS. (A) Chemical composition of PAUP. (B) Serum components in broilers of the control group. (C) Serum components in broilers of the PAUP‐H group.
PAUP Increased Growth Performance in Broilers Infected With
E. coli
3.2
As shown in Figure 2C–F
E. coli infection significantly reduced body weight gain and increased the feed conversion ratio (FCR), indicating impaired growth performance. FCR was lower in the PAUP‐L group compared with the model group (p < 0.05), whereas gentamicin did not produce a notable improvement. After a 7‐day recovery period, broilers in the PAUP‐treated groups exhibited significantly increased weight gain and decreased FCR compared to the model group. These results demonstrate that PAUP improved the growth performance in E. coli ‐infected broilers, with its growth‐promoting effects persisting for at least 1 week following treatment withdrawal.
Effects of PAUP on growth performance in broilers infected with E. coli . (A) Liver index of broilers during the drug treatment period. (B) Liver index of broilers to recover for an additional 7 days after drug treatment. (C) weight gain, and (E) feed conversion ratio (FCR) of broilers during the drug treatment period. (D) weight gain and (F) FCR of broilers to recover for an additional 7 days after drug treatment. Bars with the same letter indicate no significant difference (p > 0.05), while bars with different letters indicate a significant difference (p < 0.05).
The liver index was significantly elevated in the model group compared with the control group (p < 0.05, Figure 2A), but this increase was attenuated by PAUP‐M treatment (p < 0.05). After the 7‐day recovery period, liver index in all drug‐treated groups remained unchanged relative to measurements taken at the end of the treatment period.
PAUP Alleviated Alterations in Serological Enzymatic Activity in Broilers Infected With
E. coli
3.3
Compared with the control group, the model group exhibited significantly increased serum aspartate aminotransferase (AST) and total bile acid (TBA) levels (p < 0.05), accompanied by a significant reduction in triglyceride (TG) levels (p < 0.05, Table 4). Administration of PAUP attenuated these alterations across all measured indices relative to the model group.
PAUP Reduced Liver Bacterial Load and Histopathological Damage in Broilers Infected With
E. coli
3.4
As shown in Figure 3A
E. coli infection resulted in a marked increase in hepatic bacterial load. This elevation was markedly reduced in all PAUP‐treated groups as well as in the gentamicin group. By day 7 of the recovery period, bacterial counts were undetectable in both the PAUP‐L and gentamicin groups.
Effects of PAUP on liver bacterial load and histopathological changes in broilers infected with E. coli . (A) Liver bacterial load. (B) Liver histopathological changes examined by H&E staining. Blue triangles indicate liver abscesses, yellow triangles indicate inflammatory cell infiltration, and red triangles indicate hepatic vacuolation.
E. coli infection induced visible white necrotic spots on the liver surface and evident hepatic abscesses under microscopic examination (Figure 3B). Histopathological assessment revealed fibrous tissue proliferation, congestion, inflammatory cell infiltration, and central necrosis with hepatocellular degeneration. In the PAUP‐H group, only occasional white necrotic spots and mild microvascular congestion were observed. The PAUP‐M group showed no gross abnormalities macroscopically, although mild hepatic abscesses were detected microscopically and were notably less severe than those in the model group. No macroscopic lesions were detected in the PAUP‐L group. However, histological examination demonstrated loosely arranged hepatocytes and mild perivascular inflammatory cell infiltration compared with the control group. In the gentamicin‐treated group, liver abscesses were absent, but hepatocytes displayed ballooning degeneration.
PAUP Alleviated Liver Oxidative Damage in Broilers Infected With
E. coli
3.5
Compared with the control group, E. coli infection significantly elevated hepatic levels of SOD, MDA, and GSH (p < 0.05, Figure 4 A‐C). PAUP treatment markedly reduced MDA and GSH concentrations (p < 0.05). After a 7‐day recovery period, hepatic SOD, GSH, and MDA levels in all treated groups returned to values comparable to those of the control group (p > 0.05, Figure 4D–F).
Effects of PAUP on oxidative damage in liver of broilers infected with E. coli . (A–C) Hepatic levels of SOD, MDA, and GSH following drug administration. (D, E) Hepatic SOD, MDA, and GSH levels after 7 days of recovery. (G–K) Protein expression of factors related to the Nrf2‐Keap1 signaling pathway. (L) Hepatic ROS expression detected by immunofluorescence.
Hepatic ROS levels were significantly elevated in the model group relative to the control (Figure 4L). PAUP treatment significantly reduced ROS levels, whereas gentamicin produced no significant effect. Analysis of the NRF2‐KEAP1 signaling pathway revealed that protein expression levels of KEAP1, NAD(P)H quinone dehydrogenase 1 (NQO1), and heme oxygenase‐1 (HO‐1) were significantly decreased in the model group relative to the control group (Figure 4G–K). PAUP treatment significantly upregulated NRF2, KEAP1, NQO1, and HO‐1 protein expression, with the robust activation observed in the PAUP‐L group (p < 0.05). These results indicate that PAUP mitigates hepatic oxidative damage by enhancing the NRF2‐KEAP1 antioxidant signaling pathway.
PAUP Alleviated Liver Inflammatory Inhury in Broilers Infected With
E. coli
3.6
Compared with the control group, the model group exhibited significantly elevated protein expression levels of TLR4, IL‐6, and TNF‐α, along with increased phosphorylation of NF‐κB p65 (reflected by the P‐NF‐κB p65/NF‐κB p65 ratio) and decreased phosphorylation of I‐κB (P‐I‐κB/I‐κB ratio) (p < 0.05, Figure 5). Treatment with PAUP‐M and PAUP‐L significantly attenuated these inflammatory alterations (p < 0.05), demonstrating stronger anti‐inflammatory effects than gentamicin. These results indicate that PAUP mitigates hepatic inflammatory damage in E. coli –infected broilers, through suppression of the TLR4/NF‐κB signaling pathway.
Effects of PAUP on the TLR4/NF‐κB signaling pathway in the liver of broilers infected with E. coli . (A–F) Protein expression levels of TLR4, NF‐κB, p‐NF‐κB, IκB, p‐IκB, IL‐6, and TNF‐α detected by Western Blot.
Discussion
4
In this study, UPLC‐MS was used to analyze the chemical composition of PAUP and to identify its serum metabolites in broilers. A total of 718 constituent compounds were detected, predominantly lipid molecules, flavonoids, phenolic acids, and their oxidized derivatives. Following oral administration, 130 compounds exhibited elevated serum levels compared to controls. Key circulating metabolites included neoagarobiose, araliadiol, baicalin, sanggenol P, (−)‐Gallocatechin, 7,8‐Dihydroxyflavone, 6‐demethoxytangeretin, excavatin M, atractylenolide I, and benzyl derivatives. Previous studies have shown that several of these PAUP constituents possess antioxidant, anti‐inflammatory, and antimicrobial bioactivities. For example, baicalin demonstrates inhibitory effects against E. coli isolated from bovine mastitic milk and reduces antimicrobial resistance (Zhao et al. 2018). It also suppresses co‐infection by Mycoplasma gallisepticum and E. coli in chickens by inhibiting the NF‐κB signaling pathway (Wu et al. 2020). (−)‐Gallocatechin demonstrates antioxidant activity through inhibition of the phosphorylation of ERK and JNK (Park et al. 2021). Our previous work demonstrated that E. coli O157:H7 infection induces hepatic colonization, oxidative damage, and inflammatory responses in broilers with colibacillosis (Guo et al. 2024). In the present study, histopathological assessment similarly revealed multifocal necrotic spots, hepatic abscess formation, vascular congestion, and inflammatory cell infiltration in infected livers. PAUP administration significantly attenuated these pathological alterations. Furthermore, PAUP‐treated broilers exhibited improved growth performance, reduced liver bacterial burden, decreased serum AST and TBA concentrations, and attenuated liver tissue injury. These findings demonstrate the therapeutic potential of PAUP for mitigating E. coli ‐induced hepatic injury in broilers and support its development as a promising alternative or adjunct to conventional antimicrobial therapies.
Upon E. coli invasion, disruption of redox homeostasis within the host organism triggers pronounced oxidative stress (Yu et al. 2024). Mitochondria are the primary source of reactive oxygen species (ROS), and the liver contains a high density of mitochondria; it is a principal target organ vulnerable to ROS‐mediated oxidative damage (Zhang et al. 2022). In this study, E. coli infection induced evident hepatic injury accompanied by notably elevated intracellular ROS levels in hepatocytes. GSH, a key endogenous antioxidant, is synthesized in response to increased ROS to mitigate oxidative damage (Niu et al. 2021). The significantly increased GSH levels observed in E. coli ‐infected broilers suggested a compensatory antioxidant response to oxidative challenge. However, persistently high GSH concentrations may also indicate ongoing oxidative burden and compromised antioxidant capacity. PAUP treatment significantly attenuated hepatic ROS accumulation and restored GSH concentrations to baseline, demonstrating its capacity to alleviate oxidative stress and reestablish redox homeostasis.
Nrf2 serves as a master regulator of cellular defense against oxidative stress by orchestrating the transcription of numerous genes involved in maintaining redox homeostasis (He et al. 2020). One of its key downstream effectors is HO‐1, a stress‐inducible enzyme with potent antioxidant and cytoprotective properties (Zhang et al. 2021). Previous studies have demonstrated that 7,8‐dihydroxyflavone exerts antioxidant effects via induction of HO‐1 expression and suppression of caspase‐3/PARP activation (Chen et al. 2024). Atractylenolide‐I attenuates oxidative stress and Parkinson's disease–related pathology by activating the SIRT1/PGC‐1α/Nrf2 signaling axis (Gao et al. 2024). In the present study, PAUP significantly enhanced the Nrf2/HO‐1 signaling pathway, as demonstrated by the upregulated expression of Nrf2 and its downstream antioxidant proteins, including Keap1, NQO1, and HO‐1. This activation plays a critical role in strengthening the cellular defense system against oxidative stress. Moreover, PAUP treatment significantly reduced hepatic ROS levels, further indicating its protective effect against oxidative damage. These findings demonstrate that PAUP not only mitigates the acute oxidative burden induced by E. coli infection but also activates key endogenous defense mechanisms, thereby bolstering cellular resilience to oxidative injury.
Nuclear factor kappa‐B cells (NF‐κB) is essential for mediating immune responses to bacterial infections (Khan et al. 2024). Its activation can proceed through the canonical pathway, characterized by I‐κB phosphorylation and degradation, followed by nuclear translocation of the p65 subunits or through noncanonical mechanisms. Lipopolysaccharide (LPS) engages TLR4 and activates NF‐κB via MyD88, while ROS can also regulate NF‐κB activation (Sun 2017). Atractylenolide I has been shown to alleviate APAP‐induced hepatic oxidative damage through modulation of the TLR4/MAPKs/NF‐κB axis (Du et al. 2022). In the present study, E. coli infection activated NF‐κB predominantly through upregulation of TLR4 and accumulation of ROS, rather than through increased I‐κB phosphorylation, leading to elevated production of IL‐6 and TNF‐α. PAUP significantly attenuated TLR4 expression and ROS levels, thereby suppressing NF‐κB signaling and alleviating liver inflammation in broilers.
Conclusions
5
These results demonstrate that PAUP concurrently modulates both the TLR4/NF‐κB signaling and the Nrf2 antioxidant pathway, thereby establishing a comprehensive protective mechanism that effectively counteracts E. coli ‐induced liver damage in broilers. These findings highlight PAUP's potential to improve poultry health and food safety, reducing bacterial contamination and oxidative damage. This study provides a scientific basis for translating such natural interventions to support human health through safer, more nutritious animal‐derived food products.
Author Contributions
Conceptualization and investigation and funding: Lu‐Ping Tang; Conceptualization: Yong‐Ming He; Investigation and methodology and writing – original draft: Jia‐Ci Cai, Yan‐Na Guo and Shao‐Shan Liang: Formal analysis and supervision and software, Yan Liu, Fu‐Qiang Huang, and Qi‐Peng Lv: Data curation and resources: Lan‐Yi Zhang, Yi Qin, Xiao‐Jing Chen and Yu‐Xin Liang. All authors have read and agreed to the published version of the manuscript.
Funding
National Natural Science Foundation of China [Grant No. 32402930]. Guangdong Provincial Department of Science and Technology [Grant No. 2024A1515030170]. Department of Education of Guangdong Province [Grant No. 2024KTSCX208]. Science and Technology Projects of Xizang Autonomous Region, China [Grant No. XZ202501ZY0122].
Ethics Statement
All animal work is carried out in accordance with the Guidelines for the Care and Use of Laboratory Animals formulated by the Ministry of Science and Technology, PRC (approval No.: 2006–398) and approved by the Laboratory Animal Management Committee of Foshan University. All efforts were made to minimize animal suffering.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barbarossa, A. , A. Rosato , F. Corbo , et al. 2022. “Non‐Antibiotic Drug Repositioning as an Alternative Antimicrobial Approach.” Antibiotics 11, no. 6: 816. 10.3390/antibiotics 11060816.35740222 PMC 9220406 · doi ↗ · pubmed ↗
- 2Chen, T. X. , S. K. Wang , Y. Q. Zhang , et al. 2024. “7,8‐Dihydroxyflavone Displayed Antioxidant Effect Through Activating HO‐1 Expression and Inhibiting Caspase‐3/PARP Activation in RAW 264.7 Cells.” Journal of Biochemical and Molecular Toxicology 38, no. 1: e 23602. 10.1002/jbt.23602.38053484 · doi ↗ · pubmed ↗
- 3Dong, X. J. , L. B. Xu , Y. Wang , and H. Zhu . 2021. “Study on the Effects of Polygonum Chinense L. Capsules on Liver Fibrosis Rats Induced by C Cl 4 .” Research of Zhuang and Yao Ethnic Medicine 1: 96–106+187.
- 4Du, Z. , Z. Ma , S. Lai , et al. 2022. “Atractylenolide I Ameliorates Acetaminophen‐Induced Acute Liver Injury via the TLR 4/MAP Ks/NF‐κB Signaling Pathways.” Frontiers in Pharmacology 13: 797499. 10.3389/fphar.2022.797499.35126160 PMC 8815859 · doi ↗ · pubmed ↗
- 5Gao, Y. , S. Li , S. Zhang , et al. 2024. “Atractylenolide‐I Attenuates MPTP/MPP+‐Mediated Oxidative Stress in Parkinson's Disease Through SIRT 1/PGC‐1α/Nrf 2 Axis.” Neurochemical Research 50, no. 1: 18. 10.1007/s 11064-024-04258-x.39556135 · doi ↗ · pubmed ↗
- 6Guo, Y. N. , R. W. Mou , M. H. Lu , S. S. Liang , Y. M. He , and L. P. Tang . 2024. “Three Different Routes of EHEC O 157:H 7 Infection Were Used to Establish EHEC Broiler Model.” Poultry Science 103, no. 4: 103561. 10.1016/j.psj.2024.103561.PMC 1090784438417337 · doi ↗ · pubmed ↗
- 7Guo, Z. , X. Wang , Y. Li , et al. 2025. “Evaluation of the Therapeutic Effect of Pomegranate Peel Ginger Ultrafine Powder on Chronic Enteritis in Mice by Regulating Intestinal Microbiota.” Frontiers in Immunology 16: 1563225. 10.3389/fimmu.2025.1563225.40165946 PMC 11955692 · doi ↗ · pubmed ↗
- 8He, F. , X. Ru , and T. Wen . 2020. “NRF 2, a Transcription Factor for Stress Response and Beyond.” International Journal of Molecular Sciences 21, no. 13: 4777. 10.3390/ijms 21134777.32640524 PMC 7369905 · doi ↗ · pubmed ↗