Multidrug-resistant Klebsiella pneumoniae and Klebsiella oxytoca isolated from backyard broiler chickens and their contacts with antimicrobial resistance genes of Klebsiella pneumoniae
Shimaa El Baz, Abdelfattah H. Eladl, Reham A. El-Shafei, Mohamed A. Elmorsy

TL;DR
This study found high rates of multidrug-resistant Klebsiella in backyard chickens and their human contacts, highlighting a public health concern.
Contribution
The study identifies specific antimicrobial resistance genes in Klebsiella isolates from chickens and humans in a backyard setting.
Findings
Klebsiella pneumoniae and K. oxytoca showed high resistance to erythromycin and clindamycin in both chickens and humans.
The most common serotype of K. pneumoniae was K1, and resistance genes like ermB, blaTEM, and blaCTX-M1 were frequently detected.
Multidrug-resistant Klebsiella isolates were found in 11.3% of chicken and 15% of human stool samples.
Abstract
The objective of this study was to investigate the prevalence and phenotypic identification of antimicrobial resistance of K. pneumoniae and K. oxytoca recovered from backyard broiler chickens and their human contacts. The serotypes and genotypes of antibiotic resistance genes of K. pneumoniae isolates were investigated. A total of 80 samples were collected from backyard broiler chickens that showed signs of illness, sneezing, coughing, and diarrhea. Twenty stool samples were collected from individuals who had been in contact simultaneously. In total, 19 Klebsiella species were isolated, 12 of which were from broiler chicken samples and seven from human stool samples. Recovery rates of K. pneumoniae were 11.3% (n = 9/80) and 15% (n = 3/20) of broiler and human stool samples, respectively. K. oxytoca was detected in 3 of 80 (3.75%) broiler chicken and 4 of 20 (20%) human stool samples.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
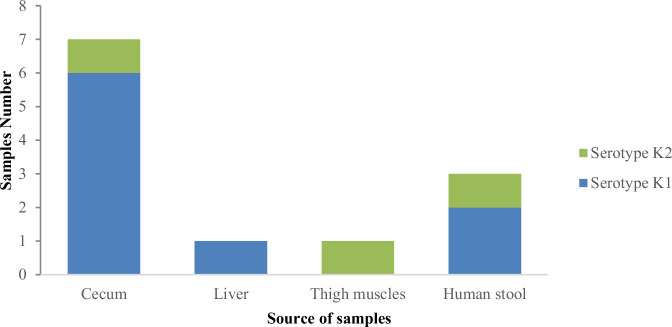 Figure 1
Figure 1 Figure 2
Figure 2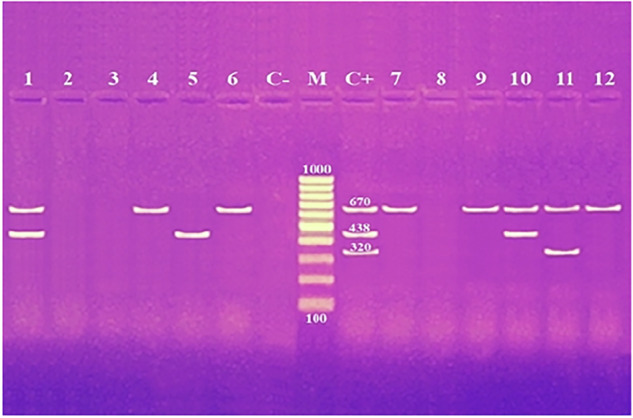 Figure 3
Figure 3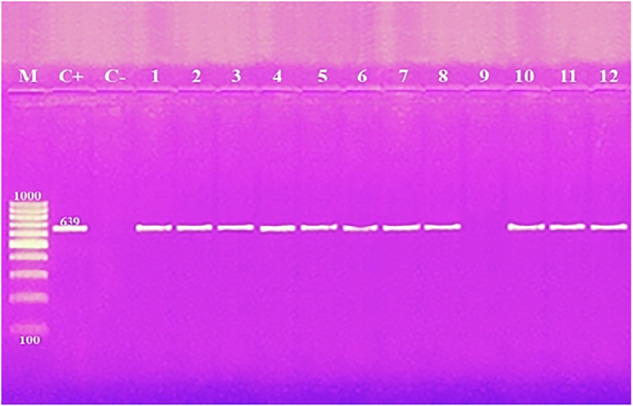 Figure 4
Figure 4 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial biofilms and quorum sensing · Pharmaceutical and Antibiotic Environmental Impacts
Introduction
Klebsiella is a member of the Enterobacteriaceae family, one of the largest groups of Gram-negative bacteria [1]. The species of greatest clinical importance within this genus are K. pneumoniae and K. oxytoca, which are opportunistic pathogens responsible for serious diseases in both humans and animals, raising public health concerns [2]. The most important reason why this pathogen is of great concern is the problem of antimicrobial resistance that hampers efforts to control it. K. pneumoniae is primarily found in the oropharynx and is associated with blood, urinary tract, and respiratory tract infections in humans [3]. It is a major cause of hospital-acquired infections and actively produces enzymes that confer resistance to β-lactam antibiotics [4].
One of the important pathogens in poultry farming that has emerged is K. pneumoniae, which mainly causes respiratory symptoms in broiler chickens, which can lead to high mortality rates and significant economic losses. The isolation rate of this bacterium has been reported to range from 9% to 35% [5]. Some factors of Klebsiella pathogenicity have been proposed, namely the smooth lipopolysaccharide (LPS) antigen (O antigen) and the capsular polysaccharide antigen (K antigen, the most important virulence factor in K. pneumoniae) [6]. Additionally, fimbrial adhesion proteins facilitate the adherence of bacterial cells to host tissues [2].
In poultry farming, antibiotics are often used not only as medicines but also as prophylactic agents and growth promoters [7]. As a result, Klebsiella - a notable zoonotic pathogen- frequently exhibits multidrug-resistant (MDR). This leads to increased growth of super-resistant bacterial strains and the risk of foodborne diseases that are transmitted to humans through the food chain [8]. The emergence of MDR bacteria poses significant therapeutic challenges worldwide, particularly in the treatment of infections caused by Klebsiella species, especially K. pneumoniae, due to its ability to acquire and disseminate resistance genes [9]. K. pneumoniae commonly harbors multidrug resistance (MDR) determinants, including plasmid-mediated quinolone resistance (PMQR) and extended-spectrum β-lactamase (ESBL) genes, most notably blaOXA, blaCTX, and blaTEM which confer substantial resistance to β-lactam antibiotics [10, 11]. The first line of treatment for serious infections caused by K. pneumoniae that produces ESBLs is carbapenems. As the most common bacteria associated with K. pneumoniae carbapenemase (KPC) resistance determinants, carbapenem-resistant K. pneumoniae strains have emerged [12].
The zoonotic potential of K. pneumoniae has been emphasized, for example, with research highlighting its presence in food animals and retail meat, and linking it to human infection through direct contact and consumption. This trend indicates that these bacteria could be transmitted from animals to humans through animal contact and food consumption, posing a major public health risk [13]. Therefore, it is important to conduct regular monitoring and surveillance of resistant bacteria in foods to implement appropriate control strategies.
The two main types of poultry production – intensive (commercial) and extensive (backyard) – are used all over the world, including Egypt. Klebsiella has emerged as a growing problem for poultry and humans worldwide. Despite the increasing prevalence of Klebsiella species in the food chain, research on K. pneumoniae and K. oxytoca in backyard broiler chickens and the risk of human infection through animal contact or food consumption remains limited worldwide, including in Egypt. Therefore, this study aimed to evaluate the prevalence and pattern of antibiotic resistance of these bacteria in backyard broiler chickens and humans who have contact with those chickens.
Materials and methods
Ethical approval
This study was approved by the Ethical Committee of the Faculty of Veterinary Medicine, Mansoura University, Egypt, and was conducted in accordance with the guidelines of the “Guide for the Care and Use of Laboratory Animals” with approval number: MU-ACUC (VM.R.25.02.213).
Samples collection
Birds were collected from different owners in urban areas at different localities in Dakahlia, Gharbia and Damietta Governorates in Egypt. Samples were collected over an 8-month period, from February to September 2024. The birds were Fayoumi and White Baladi breeds raised in backyards. A total of 80 broiler chicken samples from cecum, liver, heart, and thigh muscles (20 each) were collected from backyard broiler carcasses, aged 35–50 days. The chickens showed signs of illness, including loss of appetite, sneezing, coughing, and diarrhea. At the same time, 20 stool samples were collected from individuals who had been in contact with these chickens. All collected samples were sent to the laboratory for bacteriological examination in iceboxes at 4 °C.
Samples preparation
Samples were prepared as described previously [14]. Briefly, samples (25 g) were then placed in a sterile mortar, with 225 mL of buffered peptone water (Oxoid Ltd., Basingstoke, UK; BPW; CM0509B) and homogenized for 2 min. The homogenates were then transferred to sterile tubes and incubated at 37 °C overnight.
Isolation and identification of Klebsiella species
A loopful of enriched BPW was placed on MacConkey agar plates (M0007B, Oxoid Ltd., Basingstoke, UK) and incubated aerobically overnight at 37 °C [15]. Mucoid and pink colonies were then picked and subcultured on MacConkey agar (Oxoid Ltd., Basingstoke, UK). The suspected pure Klebsiella species were identified by the following biochemical tests: Indole, methyl red, Voges-Proskauer and citrate utilization (IMViC) tests, hydrogen sulfide production test, urease test, gelatin hydrolysis test, nitrate reduction test, ornithine decarboxylase (ODC) detection, L-lysine decarboxylase (LDC) detection, arginine decarboxylase (ADH) detection, β-galactosidase (ONPG) detection, and fermentation of sugars including glucose, sucrose, lactose, maltose and mannitol [16].
String test
In order to better understand the virulence mechanisms of the hypermucoviscous (HMV) and as part of the culture characterization process, phenotype string tests of K. pneumoniae were also performed to assess microviscosity [17]. In the hypermucoviscous K. pneumonia (hvKp) string test, a viscous strings longer than 5 mm when the colony contacted by a loop and then stretched vertically from the surface of the agar plate identified as a positive mucoid phenotype [18].
Serological identification of capsular antigen
Serological testing for the two antigenic types (K1 and K2) of K. pneumoniae was performed using the Quellung test “Neufeld reaction” as previously described [19]. The Quellung test was performed according to the manufacturer’s protocol, which included microscopic observation of the interaction between antigens and antibodies. A bacterial suspension (equivalent to 1.5 × 10^8^ CFU/mL) was prepared and mixed well with antiserum targeting the K1 and K2 capsular antigens of K. pneumoniae (Staten Serum Institute, Copenhagen, Denmark) on a glass slide. Microscopically, a positive Quellung test reaction is characterized by a clearly defined halo capsule surrounding the dark blue-stained bacterial cell.
Antimicrobial susceptibility testing
Phenotypic and genetic identification of Klebsiella species was performed at the Food Analysis Center, Faculty of Veterinary Medicine, Benha University, Egypt, according to the guidelines described by the Clinical and Laboratory Standards Institute [20]. The antimicrobial susceptibility test was conducted by the agar disk diffusion method against 16 agents of seven categories of clinically relevant and commonly used antibiotics. The study included 19 Klebsiella species isolates, comprising 12 isolates (K. pneumonia = 9 and K. oxytoca = 4) from broiler chickens and seven (K. pneumonia = 4 and K. oxytoca = 3) from human stool samples. The following antibiotic disks were used: erythromycin (E; 15 μg), clindamycin (CL; 10 μg), penicillin (P; 10 IU), ampicillin (AM; 10 μg), cefotaxime (CF; 30 μg), sulfamethoxazole (SXT; 25 μg), tetracycline (T; 30 μg), gentamicin (G; 10 μg), ciprofloxacin (CP; 5 μg), amikacin (AK; 30 μg), chloramphenicol (C; 30 μg), nalidixic acid (NA; 30 μg), kanamycin (K; 30 μg), meropenem (M; 10 μg), ofloxacin (O; 5 μg), and colistin (CO; 25 μg). A single colony of the tested bacterial strain was picked and spread evenly on the surface of Mueller–Hinton agar (Basingstoke, UK, Oxoid Ltd; CM0337). Antibiotic discs were placed on the inoculated plates, and then incubated at 37 °C for 16–18 h. After incubation, the diameters of the inhibition zone were measured using a micrometer and classified into sensitive, intermediate, and resistant. The susceptibility of Klebsiella species isolates to the tested antimicrobial panels was evaluated according to the guidelines described by the Clinical and Laboratory Standards Institute [20]. The following formula was used to determine the multiple antibiotic resistance (MAR) index for each strain: MAR index = number of resistances/total number of antibiotics tested. If a strain shows resistance to one of three or more antimicrobial categories, it is considered resistant. Escherichia coli (ATCC 25922) and K. pneumoniae (ATCC 700603) strains were used as quality control strains throughout the study for both culture and antimicrobial susceptibility testing.
Screening of K. pneumoniae for antimicrobial-resistance (AMR) genes
DNA extraction
A single colony of the bacterial strains was incubated overnight in 100 µL of sterile distilled water. Cellular remnants were removed by centrifugation at 12,000 × g for 5 min after cell lysis by heating to 96 °C for 15 min. The resulting supernatant was used as a DNA template for PCR amplification and stored at −20 °C until further use [21]. Only K. pneumonia strains were screened for antimicrobial resistance genes.
Detection of antimicrobial resistance genes by polymerase chain reaction (PCR)
The amplification process was performed using a thermal cycler (Master Cycler, Eppendorf, Hamburg, Germany). The sequences, product sizes, and annealing temperatures for all forward and reverse primers used are detailed in Table 1 [14, 22–27]. All primers were obtained from Promega (Promega Corporation, Madison, USA) and were used at a concentration of 10 pmol/μl for both forward and reverse primers.Table 1. Primer sequences for genotypic identification of antibiotic resistance genes of K. pneumoniaAntibiotic classTarget geneOligonucleotide sequence (5′ → 3′)Product size (bp)Annealing temperatureReferencestetAF: 5′ GTAATTCTGAGCACTGTCGC ′396554 °C for 1 min[22]TetracyclineR: 5′ CTGCCTGGACAACATTGCTT ′3QuinolonesgyrAF: 5′ CGACCTTGCGAGAGAAAT ′362654 °C for 1 min[23]R: 5′ GTTCCATCAGCCCTTCAA ′3bla_TEMF: 5′ ATCAGCAATAAACCAGC ′351654 °C for 1 min[14]β-lactamR: 5′ CCCCGAAGAACGTTTTC ′3blaCTX-M1F: 5′ AAGACTGGGTGTGGCATTGA ′367061 °C for 35 s[24]R: 5′ AGGCTGGGTGAAGTAAGTGA ′3Sulfonamidessul1F: 5′ TCACCGAGGACTCCTTCTTC ′333154 °C for 1 min[25]R: 5′ CAGTCCGCCTCAGCAATATC ′3CarbapenemsblaOXA-48_F: 5′ GCGTGGTTAAGGATGAACAC ′343861 °C for 35 s[26]R: 5′ CATCAAGTTCAACCCAACCG ′3Peptidesmcr-1F: 5′ AGTCCGTTTGTTCTTGTGGC ′332058 °C for 90 s[27]R: 5′ AGATCCTTGGTCTCGGCTTG ′3Macrolides**ermBF**: 5′** GAAAAGGTACTCAACCAAATA ′363953 °C for 30 s[14]R: 5′ GTAACGGTACTTAAATTGTTTAC ′3
To detect resistance genes, three PCR reactions were performed, consisting of two multiplex reactions and one uniplex PCR reaction
The first multiplex was used to amplify the genes tetA, gyrA, blaTEM, and sul1. Each 50 µL PCR reaction contains the following components: 25 μL of RedMasterMix (2×) Taq PCR (GENAXXON Bioscience, Ulm, Germany), 3 μL of DNA template, 2 μL of forward primers, and 2 μL of reverse primers. A total volume of 50 μl was obtained by adding nuclease-free water. Multiplex PCR conditions were performed using the procedures previously outlined [14]. The amplification process started with a pre-denaturation step at 94 °C for 2 min. The following steps were included in each of the 30 cycles: 30 s denaturation at 94 °C, 1 min of annealing at 54 °C, and 4 min of extension at 72 °C. A final extension was performed for 10 min at 72 °C.
The second multiplex was used to amplify the blaCTX-M1, blaOXA-48, and mcr-1genes. Each 50 µL reaction was used using 5 µL of DNA solution, 25 µL of RedMasterMix (2×) Taq PCR (GENAXXON, Bioscience), and 2 µL of forward and reverse primers. A total volume of 50 μl was obtained by adding nuclease-free water. Multiplex PCR was performed according to protocols developed previously [28]. The cycle parameters were 10 min at 94 °C, followed by 30 cycles of 30 s at 94 °C, 35 s at 61 °C, and 1 min at 72 °C. Finally, the final extension step lasted 9 min at 72 °C.
PCR reaction for amplification of the uniplex ermB gene consisted of 12.5 μL of My Taq™ HS Red Mix (Bioline, Meridian Life Science Inc, Ohio, United States), 1 μL of DNA extract, 1 μL of forward primer, 1 μL of reverse primer, and 9.5 μL of nuclease-free water. The PCR cycling conditions were performed according to the guidelines previously provided [29]. Thirty cycles of denaturation at 95 °C for 3 min, denaturation at 95 °C for 30 s, and annealing at 60 °C for 30 s were all part of the PCR cycling conditions. All PCR products were visualized by electrophoresis using 1% agarose gel (Puregene™, India). The gel was run at a constant voltage of 120 V for 1 h and then stained in 1 μg/mL ethidium bromide for 10 min. The gel was imaged under a gel document with ultraviolet illumination (acculab, Montreal, Quebec, Canada).
Results
Prevalence of K. pneumoniae and K. oxytoca in broiler chickens and human samples
Klebsiella species was isolated from 19 out of 100 (19%) of the total samples examined. Among these samples, K. pneumoniae was isolated from 12 out of 100 samples (12%) of the total samples, while K. oxytoca was positive in 7 out of 100 samples (7%; Table 2). Among the 12 positive K. pneumoniae isolates, 9 (11.3%) strains were recovered from broiler samples, of which 7/20 (35%), 1/20 (5%), and 1/20 (5%) were recovered from cecal, liver, and thigh muscle samples, respectively. Meanwhile, no K. pneumoniae was recovered from the heart samples. Regarding human stool samples, 3/20 (15%) of the samples contained K. pneumoniae. On the other hand, the total recovery from K. oxytoca was 3/80 (3.75%) of broiler samples, while 4/20 (20%) of human stool samples tested positive for K. oxytoca, as shown in Table 2. In chicken samples, K. oxytoca was recovered from 2/20 (10%) and 1/20 (5%) of bird liver and cecum, respectively (Table 2).Table 2. Prevalence of Klebsiella species in broiler samples and human stool regarding to biochemical identificationSamples sourceSamples numberKlebsiella sppK. pneumoniaeK. pneumoniae String testK. oxytocahvKpcKpNo.%No.%No.No.No.%Broiler chickensCecum2094573552210Liver20210150115Heart2000000000Thigh muscles2015150100Total801215911.35433.75HumanStool2073531521420Total100191912127577HvKp Hypervirulent K. pneumoniae, cKp Classical K. pneumoniae, No. Number
K. pneumoniae isolates of string test
The results of the string test showed that out of 9 K. pneumoniae isolates obtained from broiler chickens, 5 were identified as hvKp, all of which were isolated from cecal samples. In contrast, 4 isolates were classified as classical K. pneumoniae (cKp): 2 from cecal samples, 1 from liver, and 1 from thigh muscle. Regarding human K. pneumoniae isolates, two were classified as hvKp, while one was identified as cKp as shown in Table 2.
Serotyping of K. pneumoniae
Serotyping analysis revealed the K1 capsular types in 6/7, 1/1, and 2/3 of K. pneumoniae strains collected from broiler samples (cecum and liver) and stool samples, respectively. Meanwhile, the K2 capsular types were detected in 1/7 and 1/3 of K. pneumoniae strains obtained from broiler chickens’ cecum and human stool samples, respectively, as shown in Table 2 and Fig. 1.Fig. 1. Serotype identification of K. pneumoniae strains recovered from broiler chicken samples and human stool. There was no isolates recovered from heart
Phenotypic antimicrobial resistance of K. pneumoniae and K. oxytoca and MDR profiles
Nineteen samples tested positive for Klebsiella species against 16 antimicrobial agents from seven categories as shown in Table 3. The resistance rate to erythromycin and clindamycin was highest (100% each), followed by penicillin (94.7%) and ampicillin (84.2%). In contrast, the lowest resistance rate was observed for colistin and ofloxacin (10.5% each). Antimicrobial susceptibility profiling of Klebsiella species (n = 19) against 16 antimicrobial agents from seven different antibiotic classes revealed different patterns of antimicrobial resistance. It is worth noting that 100% (19/19) of Klebsiella isolates were resistant to at least two antimicrobials as shown in Table 4. Isolates obtained from broiler chickens showed a higher rate of MDR compared to isolates obtained from humans, as indicated in Table 4.Table 3. Percentage of phenotypic identification of antibiotic resistance in Klebsiella species bacteria (n = 19) obtained from broiler chicken samples and human stoolAntibiotic classesAntimicrobial agentSusceptibleIntermediateResistanceNo.%No.%No.%MacrolidesErythromycin (E)000019100LincosamideClindamycin (CL)000019100β-LactamPenicillin (P)15.3001894.7Ampicillin (AM)15.3210.51684.2CephalosporinCefotaxime (CF)315.815.31578.9SulfonamideSulphamethoxazol (SXT)421.1210.51368.4TetracyclinesTetracycline (T)631.6315.81052.6AminoglycosidesGentamicin (G)736.8315.8947.4Amikacin (AK)1052.6315.8631.6Kanamycin (K)1473.700526.3Amphenicol-class antibacterialChloramphenicol (C)1263.215.3631.6QuinoloneNalidixic acid (NA)1263.2210.5526.3CarbapenemMeropenem (M)1578.915.3315.8FluorquinolonesCiprofloxacin (CP)1157.915.3736.8Ofloxacin (O)1684.215.3210.5PeptidesColistin (CO)1789.500210.5Table 4Multiple antibiotic resistance index of Klebsiella species isolates (n = 19) recovered from broiler chicken samples and human stoolNo.Source of isolateStrainsAntimicrobial resistance profileMAR index1CecumK. pneumoniaeE, CL, P, AM, CF, SXT, T, G, CP, AK, C, NA, K, M, O, CO12CecumK. pneumoniaeE, CL, P, AM, CF, SXT, T, G, CP, AK, C, NA, K, M0.8753CecumK. pneumoniaeE, CL, P, AM, CF, SXT, T, G, CP, AK, C, NA, K0.8124CecumK. pneumoniaeE, CL, P, AM, CF, SXT, T, G, CP, AK, C0.6875CecumK. pneumoniaeE, CL, P, AM, CF, SXT, T, G0.56CecumK. pneumoniaeE, CL, P, AM, CF, SXT, T0.4377CecumK. pneumoniaeE, CL, P, AM, CF, SXT0.3758LiverK. pneumoniaeE, CL, P, AM, CF0.3129thigh musclesK. pneumoniaeE, CL, P, AM, CF0.31210StoolK. pneumoniaeE, CL, P, AM0.2511StoolK. pneumoniaeE, CL, P0.18712StoolK. pneumoniaeE, CL, P0.18713CecumK. oxytocaE, CL, P, AM, CF, SXT, T, G, CP, AK, C, NA, K, M, O, CO114CecumK. oxytocaE, CL, P, AM, CF, SXT, T, G, CP, AK, C, NA, K0.81215LiverK. oxytocaE, CL, P, AM, CF, SXT, T, G, CP0.56216StoolK. oxytocaE, CL, P, AM, CF, SXT, T, G0.517StoolK. oxytocaE, CL, P, AM, CF, SXT0.37518StoolK. oxytocaE, CL, P, AM, CF, SXT0.37519StoolK. oxytocaE, CL0.125MAR Multiple Antibiotic Resistance, No. Number
Antibiotic resistance profile and multiple antibiotic resistance (MAR) index
The antibiotic resistance profile of K. pneumoniae isolates (n = 12) tested against 16 antibiotics showed that these isolates were resistant to at least three antibiotics. In cecal isolates, K. pneumoniae showed resistance to all antibiotics tested, resulting in a MAR index equal to one. In contrast, stool isolates showed low resistance, with a MAR index equal to 0.187 for K. oxytoca (n = 7). High resistance was also observed in cecal isolates, which had a MAR index equal to one, while resistance in stool isolates was lower, with a MAR index equal to 0.125, as shown in Table 4.
Prevalence of antibiotic resistance gene profiles
In this study, K. pneumoniae strains showed resistance against seven types of resistance genes, including tetracycline gene tetA, quinolone gene gyrA, β-lactamase genes blaTEM, blaCTX-M1, sulfonamides gene sul1, carbapenems gene blaOXA-48, peptides gene mcr-1, and macrolides gene ermB as shown in Table 5. The results showed that the highest positive resistance rate was for the macrolide gene ermB 11/12 (91.7%), followed by the β-lactamase genes blaTEM 9/12 (75.0%) and blaCTX-M1 8/12 (66.7%), the sulfonamides gene sul1 6/12 (50%) and the quinolone gene gyrA 5/12 (41.7%). On the other hand, the lowest positive rate was for the peptide gene mcr-11/12 (8.3%), followed by the tetracycline gene tetA 2/12 (16.7%) and the carbapenemase gene blaOXA-48 3/12 (25.0%). In chicken isolates, resistance was highest for the ermB gene (8/11), followed by blaTEM (7/9), blaCTX-M1 (5/8), sul1 (5/6), and gyrA (3/5). Resistance to tetA was observed in 2/2, while no isolates carried the mcr-1 gene.Table 5. Prevalence of genotypic detection of antibiotics resistance genes (tetA, gyrA, blaTEM, blaCTX-M1, sul1, blaOXA-48, mcr-1, and ermB) among K. pneumonia strains isolated from broiler chicken samples and human stoolIsolates sourcePositive isolates for each resistance gene (%)TetracyclineQuinoloneβ-lactamaseSulfonamidesCarbapenemPeptidesMacrolidetetAgyrAblaTEMblaCTX-M1sul1**blaOXA-48mcr-1**ermBBroiler chickens (n = 9)23755208Human (n = 3)02231113Total (n = 12)2 (16.7)5 (41.7)9 (75.0)8 (66.7)6 (50.0)3 (25.0)1 (8.3)11 (91.7)
In human isolates, 3/8 carried the blaCTX-M1 gene, 2/9 carried blaTEM, 3/11 were positive for ermB, and 1/6 carried sul1. In addition, 2/3 were positive for gyrA, and 1/2 for mcr-1, with no isolates carrying tetA.
Using primers for antibiotic resistance genes tetA, gyrA, blaTEM and sul1, amplicons of length 965 bp, 626 bp, 516 bp, and 331 bp were amplified from two (16.7%), five (41.7%), eight (75.0%), and six (50.0%) of the 12 K. pneumoniae isolates examined from broiler and human samples, respectively (Fig. 2 and Table 5). Amplification of the antibiotic resistance genes blaCTX-M1, bla OXA-48 and mcr-1 resulted in the detection of amplicons at 670 bp, 438 bp, and 320 bp from eight (66.7%), three (25.0%), and one (8.3%) of the K. pneumoniae isolates examined from broiler and human samples, respectively (Fig. 3 and Table 5). When investigating the ermB gene, a 639 base pair product was identified for 11 (91.7%) of the K. pneumoniae isolates tested (Fig. 4 and Table 5).Fig. 2. Agarose gel electrophoresis of multiplex PCR of tetA (965 bp), gyrA (626 bp), blaTEM (516 bp) and sul1 (331 bp) antibiotic resistance genes for characterization of K. pneumoniae. Lane M: 100 bp ladder as molecular size DNA marker. Lane C+: Control positive for tetA, gyrA, blaTEM and sul1 genes. Lane C−: Control negative. Lanes 3 & 9: Positive K. pneumoniae strains for tetA gene. Lanes 1, 3, 6, 11 & 12: Positive strains for gyrA gene. Lanes 1, 2, 4, 5, 6, 7, 8, 10 and 11: Positive strains for blaTEM gene. Lanes 1, 3, 5, 6, 7 & 10: Positive strains for sul1gene. Lane from 1 to 9 for K. pneumoniae strains from broiler and Lane 10–12 for K. pneumoniae strains from humanFig. 3Agarose gel electrophoresis of multiplex PCR of blaCTX-M1 (670 bp), bla OXA-48 (438 bp) and mcr-1 (320 bp) antibiotic resistance genes for characterization of K. pneumoniae. Lane M: 100 bp ladder as molecular size DNA marker. Lane C+: Control positive for bla CTX-M1, blaOXA-48 and mcr-1 genes. Lane C−: Control negative. Lanes 1, 4, 6, 7, 9, 10, 11 and 12: Positive strains for blaCTX-M1 gene. Lanes 1, 5 & 10: Positive strains for blaOXA-48 gene. Lane 11: Positive strain for mcr-1 gene. Lane from 1 to 9 for K. pneumoniae strains from broiler and Lane 10–12 for K. pneumoniae strains from humanFig. 4Amplification of ermB (639 bp) antibiotic resistance gene for characterization of K. pneumoniae. Lane M: 100 bp ladder as molecular size DNA marker. Lane C+: Control positive for ermB gene. Lane C−: Control negative.Lanes from 1 to 8, 10, 11 & 12: Positive strains for ermB gene. Lane 9: Negative strain for ermB gene. Lane from 1 to 9 for K. pneumoniae strains from broiler and Lane 10–12 for K. pneumoniae strains from human
Discussion
Klebsiella and other bacteria are serious pathogens that affect both humans and animals [2, 30]. The current study highlights that K. pneumoniae and K. oxytoca bacteria recovered from backyard broiler chickens and their human contacts exhibit variable MDR. Furthermore, the MDR of K. pneumoniae carrying multiple antimicrobial resistance genes was detected in samples from backyard broiler organs (cecum, liver, and thigh muscles) and stool samples from humans who had contact with them.
In the present study, K. pneumoniae was isolated from broiler chickens at a rate of 11.3% (9 out of 80 samples), which is higher than the isolation rate (7.08%) found in a poultry farm in India [31]. In turn, it was lower than the 25.8% recorded in Norway [32]. A previous report showed the presence of K. pneumoniae in 8% of lung and liver samples taken from apparently healthy chickens in Nigeria [33]. In a study conducted in Egypt [34], Klebsiella species were isolated from different organs of diseased chickens, with the highest recovery rate from the lungs (46.67%), followed by the liver and spleen, at 20%, and then the heart at 13.33%. It is generally known that K. pneumoniae is the primary pathogen of respiratory infections in poultry or can be associated with intestinal and systemic infections. In some cases, they may act as opportunistic secondary invaders, exacerbating existing clinical conditions [34, 35]. Our study was done to investigate the possible causes of clinical illness and to maximize the likelihood of isolating pathogenic strains associated with disease. While including healthy birds, as previously described [33], would provide important information on asymptomatic carriers, our intention in this preliminary work was to concentrate on clinically affected birds.
In this study, cecal samples showed the highest detection rate of 35%, followed by liver and thigh muscles (5% each). This may be due to the fact that the anaerobic environment of the cecum promotes the growth of facultative anaerobic bacteria such as Klebsiella [36]. Contamination of poultry meat can occur during processing, especially during the evisceration stage, when cecal contents can transfer these microorganisms to the surface of the carcass, increasing the risk of foodborne illness [37]. Importantly, the heart samples did not show the presence of K. pneumoniae bacteria. Previous studies reported in Egypt showed a low rate of K. pneumoniae infection in broiler livers of 2.5% [2]. The prevalence of K. pneumoniae in chicken thigh muscles is relatively low, with only 5% of samples showing positive results. This contrasts sharply with a study by researchers [38] who found a rate of 60% (high prevalence) after analyzing chicken meat samples from stores in five countries in Europe. Among these countries, Denmark recorded the highest prevalence of K. pneumoniae in meat samples collected from different countries in Denmark, with 90%, followed by France with 73% [39]. These findings emphasize the potential importance of food in the colonization and infection of K. pneumoniae bacteria in humans [38]. The prevalence of K. pneumoniae in chicken cloacal swabs demonstrates that this bacterium is firmly established in the gastrointestinal tract of chickens, posing a serious risk of carcass contamination and potential human infection [40].
In stool samples from individuals who had contact with birds, the detection rate was 15% (3 of 20), which is much higher than the 0.5% found in outpatients in China [41], and contrasts with the 38% previously documented [42]. Furthermore, K. pneumoniae bacteria have been detected in the feces of farm workers and veterinarians [5].
In the present study, the prevalence of K. oxytoca in broiler chickens was 3/80 (3.75%), while 20% (4/20) were positive in human stool. In Egypt, 7/28 (25%) of K. oxytoca was isolated from chicken organs [43]. Smith et al. isolated K. oxytoca from 14 of 200 (7.0%) human patients in Halifax, Nova Scotia [44].
The string test was performed only for K. pneumoniae because hypermucoviscosity, which the test is designed to detect, is a recognized virulence phenotype primarily associated with this species [17, 18]. Moreover, the main objective of our study was to investigate K. pneumoniae isolates, and therefore, additional phenotypic tests were not applied to K. oxytoca. In the current study, with regard to the string test results of K. pneumoniae isolates, 7 isolates were classified as hvKp, which included 5 isolates from the cecum and 2 isolates from human stool. The remaining five isolates were identified as cKp. Both pathotypes of K. pneumoniae are found worldwide; however, hvKp is more harmful than cKp [45]. Over the past three decades, the prevalence of hvKp infection has gradually increased in the Asia-Pacific region, often affecting healthy individuals in the community [46]. Research highlights that K. pneumoniae infection can lead to serious conditions, such as central nervous system infection and endophthalmitis [47]. In contrast, cKp is the most common infectious agent in Western countries, where it typically acts as an opportunistic pathogen [48]. This pathogen primarily infects immunocompromised individuals, especially in healthcare settings [49].
The capsule is an essential virulence component that protects K. pneumoniae from phagocytosis and lethal agents in serum. K1 and K2 are the two most important serotypes, have been confirmed to be pathogenic in mice, and are commonly involved in community-acquired pneumonia [50]. According to this study, the most common serotype in humans and broilers was capsular K1 (83%, 10/12).
In this study, antimicrobial susceptibility testing showed that all tested Klebsiella isolates (n = 19) were resistant to erythromycin and clindamycin (100%). Resistance rates were 94.7% to penicillin and 84.2% to ampicillin, while only 10.5% showed resistance to colistin and ofloxacin. Abd ElGawad et al. found that all tested isolates (n = 20) showed resistance to ampicillin and oxytetracycline [2]. Furthermore, 90% (18/20) of the isolates exhibited erythromycin resistance. In a study conducted in Iraq, K. oxytoca was isolated from urine samples and found to be sensitive to ciprofloxacin. However, it showed resistance to amoxicillin and cefotaxime [51].
The rising numbers of MDR bacteria and the potential risk of zoonotic bacterial spreading from animals to humans are important issues that we need to address [52]. The lack of effective vaccines and environmentally friendly prevention methods makes it necessary to treat foodborne bacterial infections of zoonotic origin with antibiotics. The overuse of antibiotics in animals and poultry has led to an increase in the number of MDR bacteria, posing a threat to human and animal health worldwide [53].
The presence of resistance genes is one of the reasons for bacterial resistance. K. pneumoniae bacteria have various mechanisms for antimicrobial agent resistance, and the prevalence of resistant K. pneumoniae bacteria is increasing, making the treatment of infections caused by these bacteria more difficult. In the current study, K. pneumoniae isolates showed significant resistance to macrolide genes, especially ermB (91.7%). In addition, significant resistance was observed against β-lactamase genes, with blaTEM at 75% and blaCTX-M1 at 66.7%. The sulfonamide gene sul1 showed a remarkable resistance rate of 50%, while the quinolone gene gyrA showed a resistance rate of 41.7%. Furthermore, 25% of the isolates exhibited resistance to the carbapenem gene blaOXA-48. In contrast, resistance was low for the tetracycline gene tetA at 16.7% and the peptide gene mcr-1 at 8.3%. In a recent study, K. pneumoniae bacteria were isolated from broiler chickens on Chinese farms. The researchers reported that 100% of the isolates contained β-lactam resistance genes, with only one isolate showing resistance to carbapenem and peptide genes [53]. In comparison, the ESBL production rate in a healthy Indian meat and poultry farm was 5% (1 in 20) [31]. In Indonesia [14], 20% of cloacal swab samples collected from broiler farms tested positive for K. pneumoniae. All isolates contained the gyrA and bla_TEM_ genes, with 85% also containing the tetA gene and 53% the ermB gene. In a study conducted in Kenya, researchers recovered 87 isolates of K. pneumoniae and three isolates of K. oxytoca from human stool samples. These isolates were tested for antibiotic resistance genes across different classes. The results showed that the blaTEM gene was resistant in 58% of the isolates, 9% for the blaCXT-M1 gene, 18% for the tetA gene, and 31% for the sul1 gene [54]. In our study, the rate of macrolide resistance was relatively high (91.7%), which may be explained by the frequent use of macrolides in poultry farms, either for prophylaxis or treatment, and in some cases as growth promoters in poultry production. Such practices exert selective pressure that facilitates the emergence and persistence of resistant strains.
In our study, we did not conclude that direct transmission from chickens to humans occurred; rather, we reported the isolation of similar organisms from both sources. This raises the possibility of zoonotic transmission. Moreover, we isolated the same serotypes from poultry and human samples, and these isolates harbored resistance genes, further supporting the potential for cross-species dissemination of resistant strains. Actually, the number of human samples was small; however, these represented the only available samples from individuals who were in direct contact with chickens. While this limits the strength of epidemiological conclusions, our intention was not to make broad generalizations but rather to provide preliminary data highlighting the presence of the pathogen in both humans and chickens. In addition, the collection of human fecal samples poses practical and ethical challenges, which further contributed to the limited sample size. The primary goal was to establish baseline data regarding the occurrence and antimicrobial resistance of isolates from humans and chickens. The incorporating genomic analysis would reinforce these findings and intend to pursue this in future research.
Finally, Klebsiella species were detected in 19% of samples, with K. pneumoniae isolated in 12% and K. oxytoca in 7%. K. pneumoniae showed a prevalence of 11.25% in broiler chickens and 15% in human contacts, with serotype K1 dominating both. All isolates were fully resistant to erythromycin and clindamycin but showed low resistance to colistin (10.5%). The ermB gene was present in 91.6%, along with genes for resistance to β-lactams, sulfonamides, quinolones, and carbapenems. The detection of K. pneumoniae in broiler chicken samples represents a significant public health threat, especially for immunocompromised individuals, severely complicating treatment options. Thus, further studies are needed on K. pneumoniae isolates from broiler chickens and humans, including serotyping and innovative solutions to escape antibiotic resistance. It is important to pay attention to improving the level of biosecurity in backyards, which is crucial to preventing human infection with zoonotic poultry diseases resulting from human-animal interactions.
Conclusion
There was a high prevalence of MDR K. pneumoniae and K. oxytoca in backyard broiler chickens and their contacts with K. pneumoniae AMR genes in Egypt. Klebsiella species were found in a significant number of samples, with K. pneumoniae being more prevalent than K. oxytoca. K. pneumoniae has been identified in both broiler chickens and their human contacts, with serotype K1 being the most common in both. All isolates showed complete resistance to erythromycin and clindamycin, while resistance to colistin was relatively low. The ermB gene has been commonly identified, along with several resistance genes to β-lactams, sulfonamides, quinolones, and carbapenems. The spread of K. pneumoniae bacteria in broiler chickens is a major public health concern, especially for immunocompromised humans. This situation underscores the need for further research on K. pneumoniae isolates from poultry and humans, with a focus on developing effective strategies such as vaccination, the use of probiotics, enhancement of biosecurity and farming practices, and the exploration of novel antimicrobial agents like bacteriophages and antimicrobial peptides, to curb antibiotic resistance and prevent zoonotic disease transmission.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agyare C, Boamah VE, Zumbi CN, Osei FB. Antibiotic use in poultry production and its effects on bacterial resistance. Antimicrobial Resistance-A Global Threat. Intech Open Science: 2018. Open access peer-reviewed chapter. 10.5772/intechopen.79371. https://www.intechopen.com/chapters/62553.
- 2Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, et al. Bergey’s manual of systematic bacteriology. Volume 4: The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes. Springer Science, New York, USA, 2010. https://link.springer.com/book/10.1007/978-0-387-68572-4.
- 3Kot B, Witeska M. Review of antimicrobial resistance of Klebsiella pneumoniae isolated from poultry, cattle and pigs. Animal. 2024;18:101345.10.1016/j.animal.2024.10134539490087 · doi ↗ · pubmed ↗
- 4Russo TA, Marr CM. Hypervirulent Klebsiella pneumoniae. Clin Microbiol Rev. 2019;32:e 00001–19.10.1128/CMR.00001-19PMC 658986031092506 · doi ↗ · pubmed ↗