In vitro antibiofilm activity of tyrosol against single and dual-species biofilms of Candida tropicalis and Streptococcus mutans
Zarifeh ADAMPOUR, Betül YILMAZ ÖZTÜRK, Bükay YENİCE GÜRSU, İlknur DAĞ

TL;DR
Tyrosol, a natural compound, reduces biofilm formation by Candida tropicalis and Streptococcus mutans, showing potential as a treatment for oral infections.
Contribution
The study demonstrates tyrosol's antibiofilm activity against dual-species biofilms and its selective cytotoxicity.
Findings
Tyrosol inhibited biofilm formation and microbial viability in dual-species biofilms.
Tyrosol caused structural changes in biofilms as observed via microscopy.
Tyrosol showed selective cytotoxicity to fibroblast cells at higher concentrations.
Abstract
The cross-kingdom biofilm structure formed by Candida tropicalis and Streptococcus mutans may increase caries formation. The aim of this study was to evaluate the in vitro effect of the exogenous tyrosol on single- and dual-species biofilms as well as planktonic cultures formed by C. tropicalis and S. mutans. The antimicrobial efficacy of tyrosol was evaluated through broth microdilution, colony-forming unit (CFU) enumeration, and XTT reduction tests to assess cell viability and metabolic activity. Transmission electron microscopy (TEM) was used to examine ultrastructural changes in planktonic cells. Biofilm dynamics were visualized via scanning electron microscopy (SEM) and confocal laser scanning microscopy (CLSM). The in vitro cytotoxicity of tyrosol was evaluated using NIH/3T3 fibroblast cells. XTT results showed that the biofilm-reducing effect of amphotericin B (AMB) on single…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
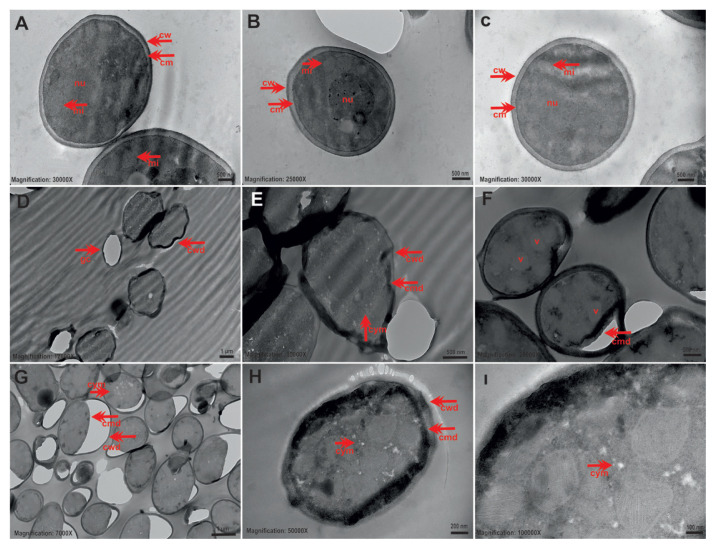 Figure 1
Figure 1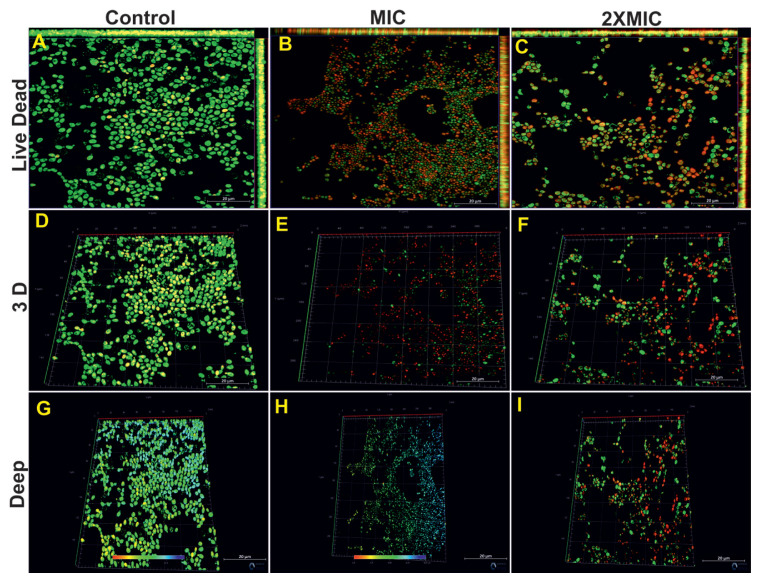 Figure 2
Figure 2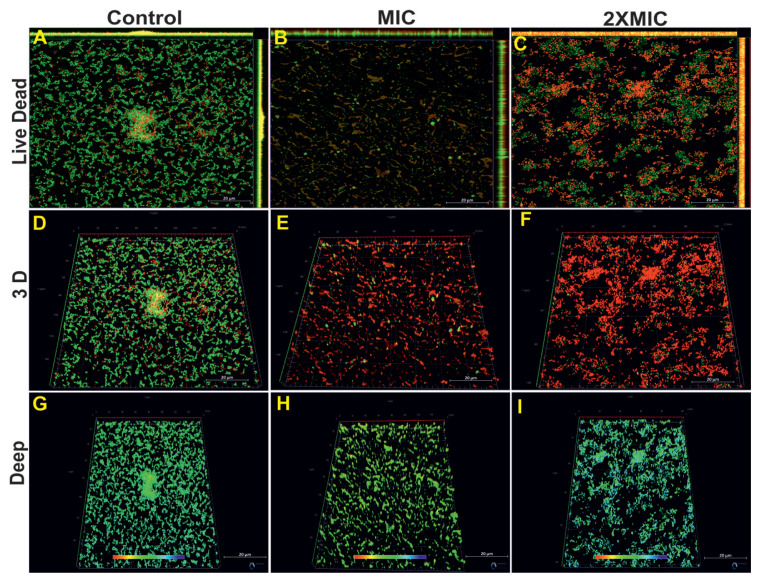 Figure 3
Figure 3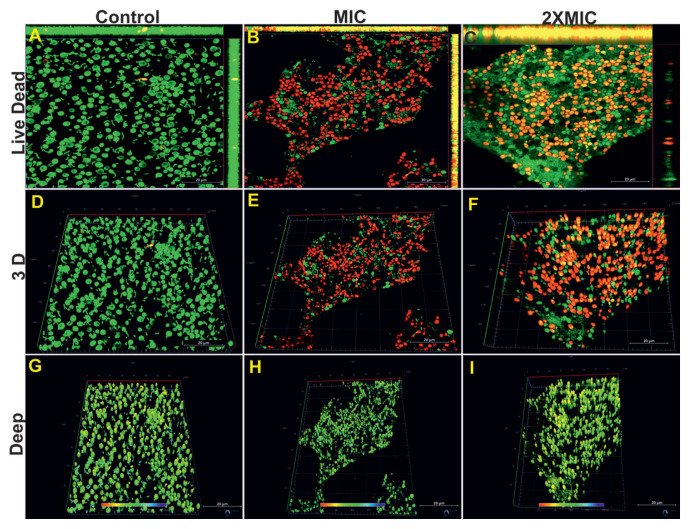 Figure 4
Figure 4 Figure 5
Figure 5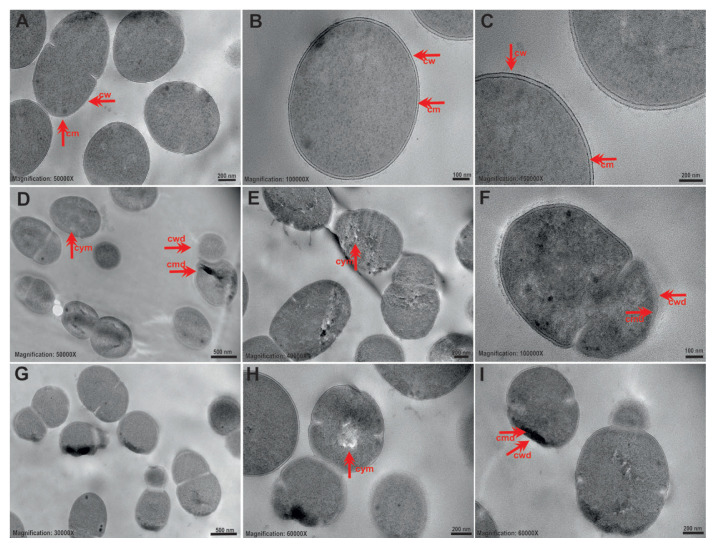 Figure 6
Figure 6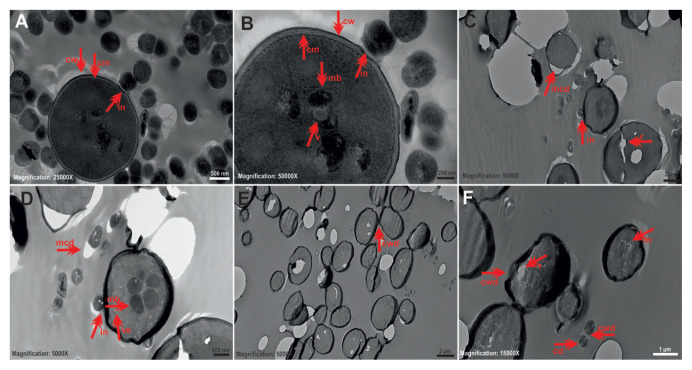 Figure 7
Figure 7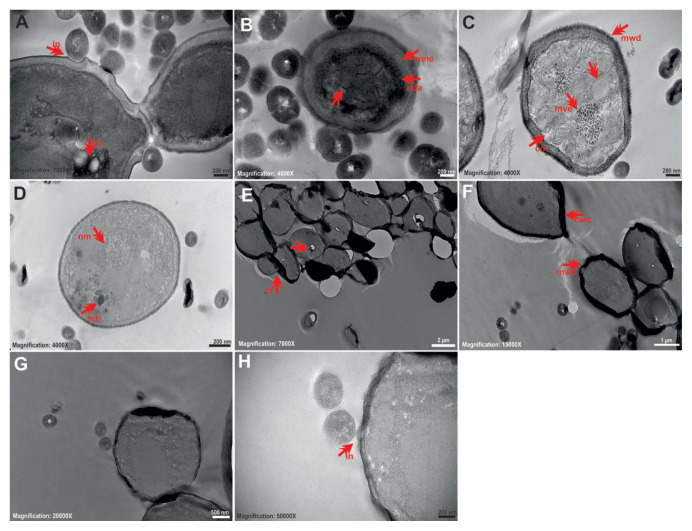 Figure 8
Figure 8 Figure 9
Figure 9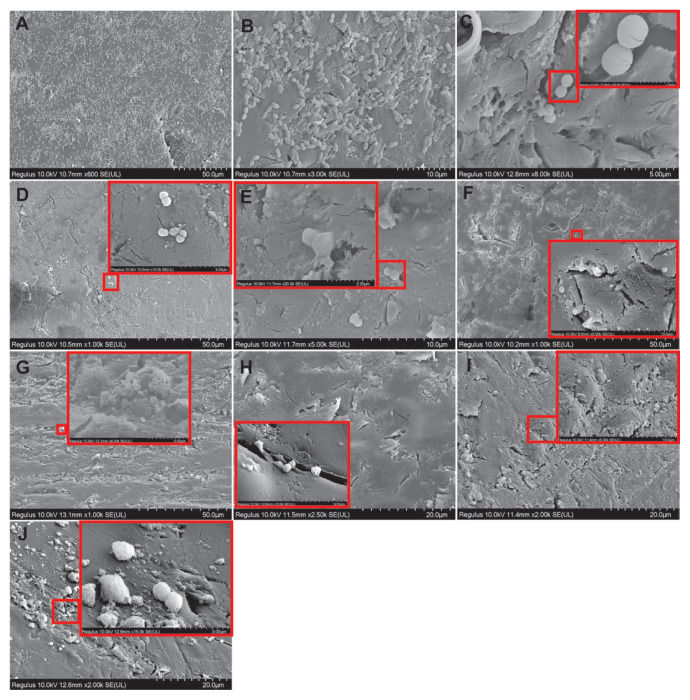 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12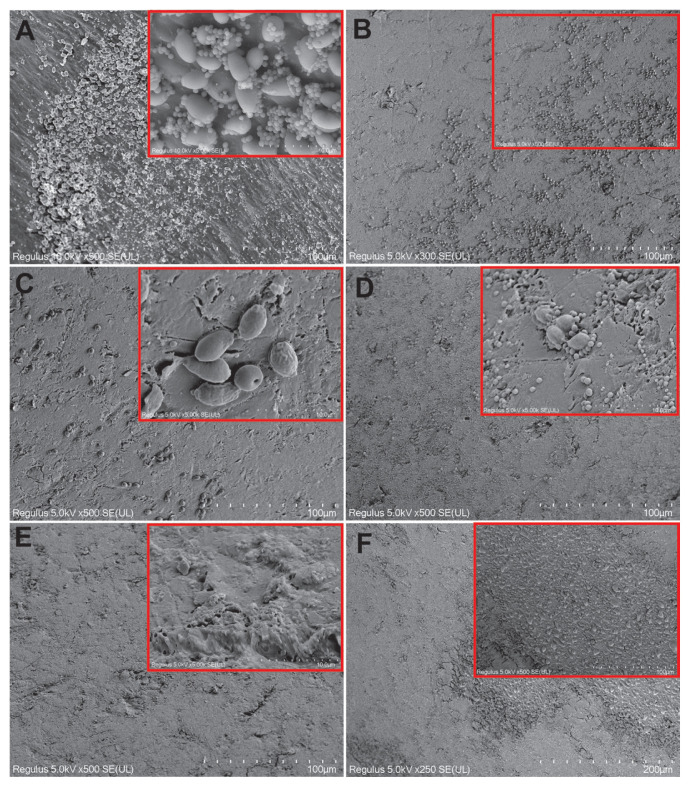 Figure 13
Figure 13- —Eskişehir Osmangazi University Scientific Research Projects Coordination Unit
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Oral microbiology and periodontitis research · Essential Oils and Antimicrobial Activity
Introduction
Colonization of teeth by cariogenic bacteria is the most significant risk factor for the development of dental diseases. Streptococcus mutans is the primary extracellular polymeric substances (EPS) producer associated with dental caries development (Kreth et al., 2004). This species, a gram-positive facultative anaerobic bacteria, has the ability to produce acid and form amyloid, and is well adapted to biofilm formation. S. mutans can form biofilms through interkingdom polymicrobial interactions with fungal Candida species during the development of dental caries. This interaction increases acid production and exopolysaccharide synthesis, thereby increasing cariogenic potential (Ribeiro et al., 2012). Candida spp. are members of the normal microbiome and act as opportunistic pathogens, causing significant mucosal and systemic infections such as oral candidiasis in immunocompromised individuals. They are particularly found in coatings, dentine, and cement surfaces. Candida albicans is the most common etiological agent, followed by C. tropicalis and C. glabrata (Fumes et al., 2018). C. tropicalis is found in mixed cultures with some Candida species and bacteria in oral infections and can form polymicrobial biofilms. This species has true hyphal formation, increasing its virulence (Fuad et al., 2023).
The most complex pathologies related to oral diseases are caused by microbial biofilms formed by a mixed group of microorganisms and protected by a complex polymer network (Salehi et al., 2020). Studies on combating S. mutans–Candida spp. dual-species biofilms in the mouth have focused intensively on C. albicans. The GtfB protein of S. mutans binds to the cell wall mannoproteins in C. albicans, resulting in increased matrix synthesis and biofilm formation. The symbiotic relationship between these two species leads to the upregulation of genes related to acid production and acid tolerance. Despite Candida species other than C. albicans, such as C. tropicalis, are frequently found in oral infections and caries lesions, their specific role in cariogenesis progression as a result of their interaction with S. mutans remain unclear (Barbosa et al., 2016). Since the cell wall composition and adhesion properties may differ in other Candida species, this could influence interactions in interspecies biofilms. Thus, comprehensive studies on this topic are needed.
A biofilm matrix provides a protective niche, increasing microbial resistance to antimicrobials and facilitating chronic infections. Therefore, reliable antibiofilm strategies and effective diagnostic tools are urgently needed (Mi et al., 2018). Advanced microscopic techniques such as scanning electron microscopy (SEM), confocal laser scanning microscopy (CLSM), and atomic force microscopy offer insights into biofilm structure and treatment effects (Hung et al., 2013; Azeredo et al., 2017). In particular, CLSM allows detailed visualization of biofilm architecture using fluorescent labeling, while SEM enables surface morphology assessment, including EPS detection.
Quorum sensing (QS) has emerged as a promising strategy to interfere with biofilm development. Signaling molecules modulate gene expression in response to cell density, thereby regulating microbial physiology, including virulence and biofilm formation. Tyrosol, a QS molecule secreted by C. albicans, induces morphogenesis in a concentration-dependent manner (Cordeiro et al., 2015). Exogenous application of tyrosol can inhibit planktonic and biofilm Candida cells, especially when combined with antifungals (Arias et al., 2016; Uzundağ et al., 2020). However, limited information exists regarding its impact on dual-species biofilms.
This study aimed to investigate the antibiofilm effects of tyrosol on single- and dual-species models of C. tropicalis and S. mutans using microbiological, ultrastructural, confocal, and cytotoxic assays.
Materials and methods
2.1. Strains, cultural conditions, and antimicrobial susceptibility tests
C. tropicalis 1678 isolate was obtained from Eskişehir Osmangazi University Health, Practice and Research Hospital, Department of Microbiology. S. mutans ATCC 25175 strain was obtained commercially. The yeast strain was activated in yeast extract peptone dextrose medium at 37 °C and then incubated in Roswell Park Memorial Institute (RPMI) 1640 broth at 37 °C for 24 h. The cultures were suspended in 0.85% physiological saline and adjusted to 0.5 McFarland standard (1–5 × 10^6^ cells/mL). The suspensions were diluted 1:50 with physiological saline and 1:20 with RPMI to reach 1–5 × 10^3^ cells/mL, in accordance with the Clinical and Laboratory Standards Institute (CLSI M27-A3, 2008) guidelines.
S. mutans ATCC 25175 was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured on brain heart infusion agar (BHI) at 37 °C in 5% CO_2_ (Zhou et al., 2025). Colonies were transferred to BHI broth and incubated statically for 18 h at 37 °C under 5% CO_2_. Cells were centrifuged at 6500 rpm for 5 min, washed with phosphate-buffered saline (PBS), and adjusted to the McFarland 0.5 standard, which corresponds to approximately 1.5 × 10^8^ CFU/mL, in accordance with the Clinical and Laboratory Standards Institute (CLSI M07-A8, 2009) guidelines, and as described by Arias et al. (2016).
For C. tropicalis, tyrosol (Sigma-Aldrich Chemical Co., St. Louis, MO, USA) was tested in concentrations from 600 to 1.17 μg/mL, prepared in sterile distilled water. A 100 μL aliquot of medium and 100 μL of serially diluted tyrosol were added to 96-well microplates. Fungal suspensions were added except in column 11 (sterility control). Amphotericin B (Sigma, USA) was tested in 16–0.0313 μg/mL concentrations. Minimum inhibitory concentration (MIC) values were determined as the lowest concentrations showing no visible growth after 24 h at 37 °C. Each test was performed in triplicate according to the CLSI M27-A3 guidelines.
For S. mutans ATCC 25175, tyrosol was tested in the 20–0.039 mg/mL range. Procedures were the same as the yeast protocol, and ampicillin (AMP) was used as a control antibiotic (330–0.31 μg/mL), following the CLSI M07-A8 guidelines. For dual cultures, isolates were coincubated in 50:50 RPMI and BHI broth. In each well, 50 μL of each cell suspension and 100 μL of tyrosol (10–0.019 mg/mL) were added. Amphotericin B (AMB) and AMP were tested again in relevant concentration ranges. All assays were performed in triplicate (Öztürk et al., 2022).
2.2. Transmission electron microscopy analysis of planktonic cells
Morphological effects of tyrosol on planktonic cells were examined using sterile 6-well plates. After inoculation and tyrosol treatment (MIC and 1/2 MIC), cultures were incubated at 37 °C for 24 h. Cells were fixed in 2.5% glutaraldehyde, postfixed with osmium tetroxide (OsO_4_), embedded in agar, and processed with standard TEM protocols, including uranyl acetate and lead citrate staining. Sections (60 nm) were analyzed with a Hitachi (Tokyo, Japan) HT7800 TEM. Dual cultures followed the same protocol using 50:50 RPMI and BHI medium (Öztürk et al., 2020).
2.3. Scanning electron microscopy analysis of biofilms
Biofilms were formed on UV-sterilized acrylic resin disks (8 × 4 mm). Acrylic resin was preferred because it is frequently used in the literature to investigate biofilm binding and structure, and because it is reproducible and easier to measure (Takamiya et al., 2021). Tyrosol was applied at MIC and 2× MIC before (prebiofilm) and after (postbiofilm) biofilm formation. Controls included no treatment, AMB, AMP, and chlorhexidine gluconate. Disks were fixed in glutaraldehyde, postfixed with OsO_4_, dehydrated in ethanol gradients, coated with gold–palladium, and examined with Hitachi Regulus 8230 SEM (Yapıcı et al., 2021).
2.4. XTT assay for biofilm metabolic activity
Biofilms were formed in 96-well plates. After treatment with tyrosol and control agents at MIC and 2× MIC, wells were incubated for 24 h. XTT and menadione solutions were prepared and added, and plates were incubated for 2 h at 37 °C. Absorbance was measured at 490–630 nm using a Chromate microplate reader (Molecular Devices, San Jose, CA, USA) (Ramage et al., 2002).
2.5. CFU quantification of biofilms
Biofilms were washed with PBS and sonicated at 30 W for 30 s. Suspensions were serially diluted and plated on Sabouraud dextrose agar and BHI for monocultures, and BHI + CHROM agar with AMB for dual cultures. CFUs were counted after 24 h at 37 °C and expressed as Log_10_ CFU/cm^2^ (Fernandes et al., 2016).
2.6. Hydrophobicity assay
Cell surface hydrophobicity was assessed using the sessile drop method. Polylysine-coated slides were inoculated and biofilms were treated with tyrosol or control drugs at 2× MIC. After 24 h incubation, contact angles were measured with an Attension Theta instrument (Biolin Scientific, Frölunda, Sweden) for 10 s after applying 10 μL deionized water (Arias et al., 2016).
2.7. Confocal laser scanning microscopy analysis
Biofilms grown in 24-well plates were treated with tyrosol at MIC and 2× MIC. LIVE/DEAD BacLight dye (SYTO 9 and PI) was applied, and samples were imaged with ZEISS (Oberkochen, Germany) LSM 800 CLSM at 40× magnification (Fernandes et al., 2018; Guo et al., 2023).
2.8. In vitro cytotoxicity assay
The NIH/3T3 mouse fibroblast cell line, derived from Swiss albino mouse embryo tissue, was used to evaluate the in vitro cytotoxicity of tyrosol. Mouse 3T3 fibroblasts were cultured in Dulbecco’s modified Eagle medium (DMEM)-high glucose with 10% fetal bovine serum and maintained at 37 °C in a humidified atmosphere with 5% CO_2_ (Kusena et al., 2021). Cells were seeded in 96-well plates and exposed to tyrosol (1–20 mg/mL) for 22 h. WST-1 solution was added (10% of well volume), incubated for 2 h, and absorbance was read at 450/630 nm (Park and Xian, 2015). Viability was calculated as:
2.9. Statistical Analysis
Statistical evaluation of the XTT assay results was conducted using data presented as mean ± standard deviation (SD) from three independent experiments, each performed in triplicate. Prior to analysis, the assumptions of normal distribution and equal variances were assessed with the Shapiro–Wilk and Levene’s tests, respectively. Once these assumptions were met, differences among control and experimental groups were analyzed using a 1-way analysis of variance (ANOVA) test, followed by Tukey’s honestly significant difference (HSD) post hoc test for pairwise comparisons. Statistical significance was defined as p < 0.05. All analyses were performed using SPSS version 26 software.
Results
3.1. Antimicrobial susceptibility tests
The MIC values of tyrosol against S. mutans ATCC 25175, C. tropicalis 1678, and dual culture are presented in Table 1. AMB is much more effective than tyrosol on the C. tropicalis 1678 isolate. The single C. tropicalis 1678 culture was more sensitive to tyrosol than S. mutans ATCC 25175. For the dual culture, the MIC value of tyrosol was 2.5 mg/mL, and while this was greater than the MIC for C. tropicalis 1678 alone, it was equal to the MIC for S. mutans ATCC 25175 alone. The MIC value of AMB in the dual culture was 0.125 μg/mL—8 times lower than the effect in C. tropicalis 1678 alone. The MIC for AMP in the dual culture was 37.5 μg/mL—30 times higher than the effect on the S. mutans ATCC 25175 isolate alone.
3.2. TEM results for single cultures
TEM findings show ultrastructural changes between control and tyrosol application groups. Control planktonic C. tropicalis cells without tyrosol had typical and healthy Candida morphology. The nucleus was clearly and centrally located, the cytoplasm was regular, and the cell wall and cytoplasmic membrane structure were observed as a whole (Figures 1A–1C). In cells exposed to tyrosol at 1/2 MIC, the cell wall and membrane were folded and disordered, and there was cytoplasmic damage, increased electron density in the cytoplasm, ghost cells, cytoplasmic melts, vacuole formations, and membrane and cytoplasm separation (Figures 1D–1F). In cells exposed to the MIC of tyrosol, increased cytoplasmic membrane separations, ghost cells, cytoplasmic melts, and mitochondria damage were observed (Figures 1G–1I).
The control group planktonic S. mutans ATCC 25175 cells had a typical and healthy bacterial morphology (Figures 2A–2C). The cytoplasm was homogeneously distributed and regular, and the cell wall and cytoplasmic membrane structure were intact. In cells exposed to tyrosol at 1/2 MIC, cell wall and membrane melting was observed in some places in addition to microbody (peroxisome) formation, but no deformations were observed in the cell and the cytoplasm remained relatively homogeneous (Figures 2D–2F). Membrane and wall damage, membrane folding, melting of the stopping center in a small number of cells, and microbodies were detected in cells exposed to the MIC of tyrosol (Figures 2G–2I).
3.3. TEM results for dual cultures
In the TEM micrographs of the control group without tyrosol, bacterial and yeast cells had integrity with round or oval-shaped cell wall and membrane structures. However, some cytoplasmic damage was observed in the central parts of the bacterial cells in addition to a few, partially microbody-like vacuoles in the central parts of the yeast cells. The morphological findings in the control group were much more regular and complete than those in the treatment groups. Yeast cells appeared to interact with some bacterial cells through their wall portions, and there was a slight change in bacteria–yeast cell wall contact points, indicative of binding (Figures 3A and 3B). When exposed to tyrosol at MIC, cytoplasmic ruptures, prominent and numerous microbodies (peroxisomes), cell membrane–cytoplasm separation, damage to the walls in the form of collapse and many small vesicles were observed in yeast cells. Cell membrane–cytoplasm separation was evident in S. mutans ATCC 25175 cells. Bacteria–yeast interactions were noticeable in some areas (Figures 3C and 3D). When exposed to tyrosol at 1/2 MIC, damage to the walls in the form of collapse, cytoplasmic ruptures, and melting were observed in yeast cells, and there was cell wall cytoplasm damage in a small number of bacterial cells (Figures 3E and 3F).
Figures 4A–4D show representative TEM images of ultrastructural changes in planktonic dual cultures exposed to AMB at MIC and 1/2 MIC. Microtubule-like filamentous structures were striking in the cytoplasm of yeast cells as a result of exposure to AMB at MIC. Additionally, cell wall membrane expansion, electron-dense appearance in the cytoplasm, yeast–bacteria interaction zones, and damage to the cytoplasm were observed (Figures 4A and 4B). When exposed to tyrosol at 1/2 MIC, there was membrane–wall damage, numerous microvesicular structures, vacuole formations, nuclear melting, cytoplasmic melting, microbodies (peroxisomes), and vacuole formations in yeast cells (Figures 4C and 4D). Figure 4E–4H show TEM images of ultrastructural changes of planktonic dual cultures exposed to AMP at MIC and 1/2 MIC. As a result of exposure to AMP at MIC, there were fewer bacterial cells, and cytoplasmic ruptures and cell wall damage were observed. In this group of cells, yeast were also fused and wall membrane damage was noticeable (Figures 4E and 4F). When exposed to tyrosol at 1/2 MIC, bacterial cells had a relatively healthier morphology, and yeast–bacteria interaction points were evident (Figures 4G and 4H).
3.4. SEM results for single yeast species biofilm
In the control group samples, the majority of C. tropicalis cells on the acrylic surface were in filamentous form and a mature biofilm structure was observed (Figures 5A and 5B). This structure consisted of a dense and heterogeneous network of yeast, pseudohyphae, and hyphae. Hyphal budding structures were also observed. The EPS structure was not clearly distinguishable, but this may have been due to the intense dehydration stages during SEM monitoring. Prebiofilm samples treated with tyrosol at the MIC had deformed and reduced biofilm structures. There were shrinkages and fusions in the hyphae structures, and bleb formations in some yeast cells (Figure 5C). In the cells exposed to tyrosol at the MIC postbiofilm formation, the biofilm was markedly reduced. The cells had shrunk due to cytosolic loss, the hyphae and biofilm structure was greatly reduced, cavities had formed in the cells, and fused cells increased (Figure 5D). When cells were exposed to tyrosol at 2× MIC prebiofilm formation, t the hyphae and pseudohyphal structures were severely damaged, fused, and disintegrated in most places. The biofilm structure reduced significantly. However, yeast cells were more structurally protected (Figure 5E). When the cells were exposed to tyrosol at 2× MIC postbiofilm formation, the hyphae structure in the biofilm was greatly reduced, and damages such as structural deteriorations, cavity formations, and cell fusion were detected in the yeast cells (Figure 5F). In the prebiofilm group samples treated with antifungal AMB at MIC, although the biofilm structure was dense, it was observed that the cells fused, deformities occurred, and cavity formations increased (Figure 5G). In the postbiofilm group, samples treated with antifungal AMB at MIC had greatly reduced cells, but the yeast cells were structurally preserved (Figure 5H). In the groups where chlorhexidine gluconate was applied, the biofilm was reduced to a great extent in the prebiofilm and postbiofilm groups, with cell shrinkage in the latter (Figures 5I and 5J, respectively).
3.5. SEM results for single bacterial species biofilm
A dense biofilm structure and cells with a typical chain structure were observed on the acrylic surface in the control samples for S. mutans ATCC 25175. The cells appeared morphologically regular (Figures 6A and 6B). When cells were exposed to tyrosol at the MIC prebiofilm formation, fewer cells were observed compared to the control. The cells that were visible were damaged (Figure 6C). When cells were exposed to tyrosol at MIC postbiofilm formation, cell numbers were significantly reduced compared to the control, and these few cells were damaged (Figure 6D). When cells were exposed to tyrosol at 2× MIC prebiofilm formation, the biofilm structure was greatly reduced and a very small number of cells observed on the surface were fused and damaged (Figure 6E). When the cells were exposed to tyrosol at 2× MIC postbiofilm formation, the biofilm was again significantly reduced and a small number of biofilm fragments were encountered (Figure 6F). The cells were almost completely reduced in prebiofilm and postbiofilm samples after treatment with the antibiotic AMP at the MIC compared to the control (Figures 6G and 6H, respectively). In a very small number of cells, there were irregularities and deformities. In the groups treated with chlorhexidine gluconate, the number of cells decreased significantly in both the prebiofilm and postbiofilm groups compared to the control group and the remaining few cells had deformities (Figures 6I and 6J, respectively).
3.6. SEM results for dual biofilms
In the control samples, the cells on the acrylic surface were healthy and there was a dense biofilm on the surface (Figure 7A). When cells were exposed to tyrosol at the MIC prebiofilm formation, yeast cells were reduced and shrunken, bacteria were reduced in size, and damage was observed on the surface of cells belonging to both microorganism groups compared to the control. However, there was no significant biofilm reduction compared to the control (Figure 7B). Application of tyrosol at the MIC postbiofilm (MIC) was more effective compared to prebiofilm (MIC) application (Figure 7C). When cells were exposed to tyrosol at 2× MIC prebiofilm formation, tyrosol was highly effective and its biofilm-reducing effect was observed (Figure 7D). When the cells were exposed to tyrosol at 2× MIC postbiofilm formation, significant damage was detected in the yeasts. However, the bacteria appeared healthier and a relative decrease in the biofilm was observed in general (Figure 7E). Thus, prebiofilm application was much more effective than postbiofilm for 2× MIC application. In the groups where chlorhexidine gluconate was applied, very effective results were obtained groups compared to the control group (Figure 7F).
Figure 8 shows the effects of AMP on dual biofilms by SEM. A dense biofilm structure was observed on the acrylic surface in the control group samples. The cells were morphologically regular and have a dense biofilm appearance (Figure 8A). When cells were exposed to AMP at the MIC prebiofilm formation, there was a small decrease in biofilm with more of an effect on bacteria (Figure 8B). When cells were exposed to AMP at the MIC postbiofilm formation, the cells were almost completely reduced and AMP was very effective (Figure 8C). Thus, postbiofilm application was more effective than prebiofilm application at the MIC. When cells were treated with AMP at 2× MIC prebiofilm formation, significant biofilm reduction and occasional yeast lysis were detected (Figure 8D). Cells were very effective in samples treated with AMP at 2× MIC postbiofilm formation (Figure 8E). Postbiofilm application was more effective than prebiofilm application at 2× MIC. Chlorhexidine gluconate treatment was more effective than the control group (Figure 8F).
Figure 9 shows the effects of AMB on dual biofilms by SEM. A dense biofilm structure was observed on the acrylic surface in the control group samples. The cells were morphologically healthy and had extensive biofilm formation (Figure 9A). When cells were exposed to AMB at the MIC prebiofilm formation, there was a reduction in biofilm (Figure 9B). When cells were exposed to AMB at the MIC postbiofilm formation, the cells were almost completely reduced and AMB was very effective (Figure 9C). Thus, there was significant biofilm reduction in both groups. Biofilm reduction was also detected when cells were treated with AMB at a 2× MIC prebiofilm formation (Figure 9D). It was found to be very effective in samples treated with antifungal AMB at 2× MIC after cells formed biofilm (postbiofilm) (Figure 9E). Thus, postbiofilm application was more effective than prebiofilm application at 2× MIC. Chlorhexidine gluconate treatment was more effective compared to the control group (Figure 9F).
3.7. XTT reduction test results
Table 2 shows the XTT absorbance values observed for single-species biofilms of C. tropicalis 1678, S. mutans ATCC 25175, and the dual biofilm of the two species. C. tropicalis biofilm cells treated with tyrosol at 2× MIC had a 39% decrease in metabolic activity and a 42% decrease at the MIC compared to the control group. However, this was not statistically significant. AMB application caused a 47% decrease in 2× MIC and 48% decrease in metabolic activity at the MIC compared to the control (p < 0.05). There was a difference between the effect of tyrosol and AMB for C. tropicalis compared to the control, where MB had a stronger biofilm-reducing effect (p < 0.05). However, neither tyrosol nor AMP had a significant biofilm-reducing effect on single S. mutans biofilm cells, and there was no significant difference between the effect of tyrosol and AMP. Although the absorbance value in the dual culture increased by approximately 15% compared to the control single absorbance value of C. tropicalis, this increase was not statistically significant. Compared to the control single absorbance value of S. mutans, the dual culture absorbance value increased by more than 100% and was statistically significant (p < 0.05). When tyrosol was applied at 2× MIC in the dual culture, there was a 33% biofilm-reducing effect compared to the control. When applied at the MIC, a 2% increase in absorbance was observed, which was not statistically significant. In the dual culture, the biofilm-reducing effect of AMB was 11% at 2× MIC and 17% at the MIC compared to the control, and this was significantly lower than the effect of AMB applied alone on C. tropicalis (p < 0.05). While AMP did not have a significant biofilm-reducing effect on single S. mutans, biofilm resistance was observed in the dual culture with an increase in absorbance of over 100% at 2× MIC and 70% increase in absorbance at MIC compared to the control. When AMB and AMP were used together, there was a 24% reduction at both concentrations.
3.8. Determination of cultivable biofilm cells
The total number of cultivable cells on the surfaces used for single and dual biofilms was evaluated by the plate counting method after incubation. The results are presented in Table 3. All dilution rates were evaluated and the dilution rate data of 10^−5^, which was observed to be most suitable for counting, was taken as the baseline. Tyrosol increased the number of cultivable S. mutans ATCC 25175 cells, while it decreased the C. tropicalis 1678 cells at MIC and 2× MIC. In the dual culture, tyrosol slightly increased biofilm at the MIC, and reduced it at 2× MIC. The count results in both tyrosol and antibiotic/antifungal application groups were similar to the XTT data. However, the coefficient differences observed in the increase or decrease rates in the application results may be due to experimental differences.
3.9. Cell surface hydrophobicity test
Table 4 shows the contact angle values obtained from single and dual biofilms as a result of the treatment of active substance at 2× MIC. In S. mutans ATCC 25175, the contact angle was 28.63° in the control group, 10.8° in the tyrosol treatment group, and 36.28° in the AMP treatment group. This showed that tyrosol at 2x MIC reduced the contact angle for S. mutans and made the surface more hydrophilic, and did not show a significant reducing effect in parallel with the XTT results. Following AMP application, the surface changed from hydrophilic to hydrophobic, in line with the XTT results. In C. tropicalis 1678, the contact angles were 13.36° in the control group, 34.62° in the tyrosol treatment group, and 20.24° in the AMB treatment group. Thus, tyrosol increased the contact angle and the surface became more hydrophobic. AMB changed the surface again from hydrophilic to hydrophobic.
For the dual biofilm, the contact angles were 18.43° for the control group, 33.08° in the tyrosol treatment group, 9.37° in the AMP treatment group, 28.34° in the AMB treatment group, and 67.56° in the antifungal + antibiotic treatment group. Tyrosol application changed the surface from hydrophilic to hydrophobic, consistent with XTT results. In the AMP application, the control group changed from hydrophobic to hydrophilic, and was similar to the XTT results. In the dual group, the AMB treatment shifted the surface character from hydrophilic as observed in the dual control group to a more hydrophobic state. Notably, the combined application of AMB and AMP resulted in a strongly hydrophobic surface character compared to the dual control group.
XTT results of the same group also showed parallel findings. Thus, as the surface became more hydrophobic, the active substance was more effective. As the drug reduced the biofilm, the surface became more hydrophobic (Table 4).
3.10. CLSM findings
In our study, a BacLight LIVE/DEAD cell viability test was used to determine whether there was cell death during fungal–bacterial biofilm development. After 48 h of incubation, live cells were determined with green fluorescent staining (Syto 9), while dead cells were determined with red fluorescent propidium iodide (PI) dye. While a dense yeast biofilm formed in the control group C. tropicalis 1678 samples, the majority of the region was dominated by green fluorescence (Figure 10A). Tyrosol application at MIC and 2× MIC significantly reduced the biofilm and dead cells were quite dense in the environment (Figures 10B and 10C). The 3D images of the same groups are shown in Figures 10D–10F. Confocal z-stack image analysis data was also used to obtain information about biofilm depth. In the control group, biofilm depth reached up to 58.40 μm (Figure 10G). Tyrosol application at MIC (39.00 μm) caused a decrease in biofilm depth. Interestingly, at 2× MIC, the biofilm depth was 52.80 μm and showed much less reduction than the control (Figures 10H and 10I).
A widespread bacterial biofilm was formed in the control group S. mutans ATCC 25175 and the majority of the region was green fluorescence (Figure 11A). There was a dense biofilm in the tyrosol application at MIC and 2× MIC, but there were also dead cells in the medium as well as live cells (Figures 11B–11C). The 3D images of the same groups are shown in Figures 11D–11F. Confocal z-stack image analysis data was also used to obtain information about biofilm depth. In the control group, biofilm depth reached up to 36.40 μm (Figure 11G). Tyrosol application did not cause a significant change in the biofilm depth at MIC (36.25 μm) and 2× MIC (32.40 μm) (Figure 11H and 11I).
While a dense yeast–bacteria biofilm formed in the dual culture control group, the majority of the region was dominated by green fluorescence (Figure 12 A). Tyrosol application at MIC and 2× MIC significantly reduced the biofilm and dead cells were found to be quite dense in the environment (Figure 12B and 12C). The 3D images of the same groups are also shown in Figure 12D–12F. When 3D images and deep images of the same group were examined with confocal z-stack image analysis, the biofilm depth reached up to 51.80 μm in the control group (Figure 12G). Tyrosol application caused a significant decrease in the biofilm depth at the MIC (33.60 μm). Interestingly, at 2× MIC, the biofilm depth was relatively high (78.40 μm) compared to the control (Figure 12H and 12I).
3.11. WST-1 cell viability test
In cytotoxicity test studies, tyrosol was studied in the concentration range of 20–1.0 mg/mL. Different effects were observed on 3T3 cell viability depending on the dose as a result of 24-hour tyrosol application. The inhibitory concentration (IC50) value of tyrosol at 24 h was 1.944. Tyrosol showed a dose-dependent toxic effect on cells in the concentration range of 1–4 mg/mL. At higher concentrations, it showed a cell proliferation-inducing effect compared to the control (Table S1). After 48 h of tyrosol application, different effects were observed on 3T3 cell viability, depending on the dose. The IC50 value of tyrosol at the 48th hour was 1.5783. Similar to the 24 h results, there was a significant toxic effect on the cells depending on the dose in concentration range of 1.6–4 mg/mL. At higher concentrations, it showed a cell proliferation-inducing effect compared to the control (Table S2).
Discussion
Microorganisms often exist within polymicrobial biofilms that may involve mutualistic, commensal, or antagonistic interactions. In the human body, the diversity of microbiota plays a critical role in maintaining microbial equilibrium; however, disruptions such as antibiotic exposure may lead to dominance by opportunistic species (Brown et al., 2012; Srinivasan et al., 2021). One such dual interaction is that between Candida spp. and S. mutans, which often form resilient, synergistic biofilms that are difficult to treat using conventional antimicrobial strategies (Ribeiro et al., 2012; Rodrigues et al., 2019). The dual biofilm formed by C. tropicalis and S. mutans is found in natural biological environments, especially in moist, nutrient-rich, and high microbial diversity natural biological environments such as the oral cavity. Other microorganisms cohabiting in this environment can have positive and negative effects on the biofilm. For example, the presence of acid-tolerant Lactobacillus species or C. albicans colocalizing with C. tropicalis may promote biofilm formation and stability, while the presence of antifungal metabolite-producing Pseudomonas aeruginosa or pH-changing Actinomyces species may suppress the growth or biofilm-forming capacity of the dominant species in the biofilm. The presence of carbon sources or QS molecules in the environment may also favor the development of this dual biofilm, while the presence of molecular agents such as hydrogen peroxide or high doses of farnesol may have an inhibitory effect.
This study showed the antimicrobial and antibiofilm efficacy of tyrosol against single and dual biofilms of C. tropicalis and S. mutans. The MIC findings for both C. tropicalis and S. mutans were lower than many previous reports (Monteiro et al., 2015; Tsikopoulos et al., 2019), indicating possible strain-specific sensitivity. Therefore, tyrosol showed also strong inhibitory effects on C. tropicalis and moderate effects in dual biofilm systems, although its efficacy against S. mutans alone was limited. The fact that S. mutans is more resistant to tyrosol than C. tropicalis may be due to differences in cell membrane and wall structure. Tyrosol may have affected ergosterol synthesis and membrane permeability in the C. tropicalis cell membrane, making yeast cells more sensitive. Since S. mutans does not contain ergosterol, it is less affected by compounds with membrane-targeted effects. On the other hand, S. mutans is a bacterium well adapted to acidic environment and stress conditions and has a highly dense and protective EPS structure. This adaptation includes more effective antioxidant defense systems against reactive oxygen species (ROS) and harmful compounds. C. tropicalis may have had a more limited response to oxidative stress from molecules such as tyrosol. Furthermore, tyrosol is a fungal QS molecule and affects processes such as morphogenesis, filamentation, or cell division in yeast cells. The bacterium S. mutans, on the other hand, may not contain a direct target for this molecule and may therefore be more resistant.
Structural analysis using SEM and TEM suggested widespread cellular and architectural damage, particularly at higher tyrosol concentrations. TEM images showed multitargeted cellular damage by tyrosol, including membrane rupture, cytoplasmic melting, and mitochondrial damage (Yapıcı et al., 2021). In dual cultures, yeast–bacterial interaction points were frequently observed, supporting the idea of physical cooperation in biofilm architecture (Pidwill et al., 2018; Krüger et al., 2019). The direct contact between yeast and bacterial cells plays an important role in biofilm organization, increasing the stability and structural integrity of the environment. Contact may facilitate genetic and biochemical signaling, nutrient sharing, and metabolic support. Also, virulence may increase due to changes in the level of antimicrobial resistance in the biofilm. With close contact, QS molecules can affect the gene expression of both bacterial and yeast cells. All of these mechanisms may cause treatment resistance problems. SEM analyses also confirmed concentration-dependent antibiofilm effects of tyrosol, with 2× MIC more effective in the prebiofilm phase, consistent with prior studies showing inhibition of EPS production at early stages (Monteiro et al., 2015). Postbiofilm applications, however, led to more dramatic biofilm disruption, particularly in mixed cultures. This was corroborated by XTT findings, where metabolic activity was significantly reduced in fungal populations (Arias et al., 2016; Fernandes et al., 2016). On the other hand, the dual-species model showed signs of protective bacterial dominance and physical interactions, potentially mediated by EPS or adhesion proteins.
There was a decrease in metabolic activity for S. mutans after tyrosol application, while an increase in cell count was observed in MIC concentrations using the CFU test. This finding suggests that despite being metabolically suppressed after the application of the active substance, the cells are still alive and cultivable. An alternative hypothesis is that the release of planktonic cells into the environment due to the dissolution of the biofilm structure during the CFU test may have led to an increase in cell count, but this may not have been observed in the XTT test, which is biofilm-based and measures only metabolic activity. The presence of viable but uncultivable cells (VBNC) in the environment may also be a helpful factor in understanding infections; however, this relationship between metabolic testing and VBNC viability in culture must be confirmed using methods such as ATP measurement or live/dead staining.
Our findings support S. mutans having a tolerance to tyrosol and that this increases under dual conditions. This result is consistent with reports in the literature that S. mutans can resists active agents through EPS modulation and gene regulation pathways (Senadheera et al., 2007; Nan et al., 2009). Our CFU and XTT findings indicate that S. mutans increases in a dual biofilm structure and has bacterial dominance under stress conditions. Collaborative metabolisms between species or interactions related to EPS are also thought to contribute to this resilience (Silverman et al., 2010; Ellepola et al., 2019; Lobo et al., 2019; Wu et al., 2020).
The increase in AMP resistance in the dual biofilm structure was due to the microbial interaction between the two species and the unique characteristics of the biofilm environment. Compared to biofilms formed by single species, more complex resistance mechanisms are found in synergistic dual biofilms. For example, changes in pH, oxygen, or environmental factors in the environment, or the presence of metabolically slow-growing persister cells, can reduce the effectiveness of antibiotics such as AMP. The dense and complex matrix structure of the dual-species biofilm may significantly limit antibiotic diffusion or reduce its effectiveness. Alternatively, S. mutans cells may be protected from the antibiotic effects due to their physical positioning between C. tropicalis hyphae as a result of bacteria–yeast contact. Another possible mechanism is that antibiotic resistance genes may be induced through the QS mechanism between the two species, and further in-depth studies are needed to elucidate all these complex mechanisms.
CLSM results confirmed increased cell death in tyrosol-treated samples but also showed paradoxical increases in biofilm depth in dual cultures at high concentrations. This may be due to EPS overproduction, a defensive response reported previously (Falsetta et al., 2014; Li et al., 2023).
In biofilm studies, the changes in hydrophobicity observed after antimicrobial application are closely related to the EPS composition of the biofilm. Since the EPS structure determines the adhesion, protective properties, and hydrophobicity of the biofilm, the applied agent can reduce EPS production or change its composition. When the ratio of molecules in EPS changes, the hydrophilic or hydrophobic character of the surface changes. Therefore, not only the amount of biofilm but also the changes in chemical composition should be carefully considered in the evaluation of hydrophobicity changes. In support of this, Bourroubey et al. (2024) showed that changes in hydrophobicity directly affect EPS formation and matrix structure, suggesting a dynamic interplay between surface properties and biofilm development. Our hydrophobicity tests showed increased contact angles after tyrosol treatment in several groups, suggesting altered surface adhesion and matrix composition (Chandra et al., 2005; Khoury et al., 2020; Bourroubey et al., 2024).
In our study, NIH/3T3 fibroblast cells were used to assess the cytotoxicity of tyrosol. The results showed a concentration- and time-dependent biphasic response. At concentrations up to 4 mg/mL, tyrosol suppressed cell proliferation, whereas at higher concentrations, it unexpectedly promoted cell viability. Taken together, these results suggest that tyrosol has a narrow therapeutic window, where its biological impact depends critically on concentration and exposure time. These findings imply a hormetic effect and highlight the need for further investigation to clarify the underlying mechanisms of this dual behavior. Further mechanistic studies, such as caspase activity assays and mitochondrial membrane potential analysis, are warranted to elucidate the exact pathways involved in this concentration-dependent dual response. Although tyrosol is a phenolic compound with weak antioxidant properties, it can have proliferative effects at high doses under certain conditions. In our study, the increase in cell proliferation caused by high doses of tyrosol (4.5–20 mg/mL) may have been achieved by reducing the oxidative stress level of cells in a stressful environment and promoting proliferation. On the other hand, the application of high doses of tyrosol to cells may have triggered an adaptive protective response. There are studies in the literature showing that high doses of tyrosol can have a proliferative effect on some cell lines (Zhang et al., 2019; González-Acedo et al., 2023). However, further detailed studies on this subject are needed. In the study conducted by Loru et al. (2009), the cytoprotective effects of hydroxytyrosol and tyrosol against oxidative damage were evaluated in LLC-PK1 renal epithelial cells (Loru et al., 2009). Cell viability assays were performed after 24-hour exposures to concentrations ranging from 25 to 2500 μM. While hydroxytyrosol significantly reduced cell viability at concentrations above 500 μM, tyrosol had no apparent cytotoxicity across the tested range, suggesting a more favorable safety profile. Similarly, Lee et al. (2016) reported that tyrosol inhibited the growth of KB human oral cancer cells in a time- and dose-dependent manner, indicating that its biological effects may vary depending on the cell type and experimental conditions (Lee et al., 2016).
Polymicrobial interactions have complex interspecies dynamics that significantly influence biofilm formation and development. Interspecies interactions between non-albicans Candida species and S. mutans and biofilm structures are attracting increasing attention, particularly due to their role in exacerbating dental caries. Currently, natural compounds, antimicrobial peptides, or nanomaterials are being extensively researched as treatment strategies. Tyrosol, like farnesol, is a QS molecule that regulates filamentation in C. albicans. While farnesol inhibits biofilm formation by facilitating the transition from the hyphal form to the yeast form, tyrosol initiates hyphal conversion in yeast but suppresses the QS system at high doses, thereby reducing biofilm formation. Other proposed mechanisms for tyrosol’s antibiofilm effects include inducing oxidative stress, suppressing virulence and adhesion genes, or directly damaging intracellular components. Tyrosol can inhibit Candida biofilms in a concentration-dependent manner, but this effect is particularly promising when applied before biofilm formation. At very high doses, it has toxicity. The effect of tyrosol on S. mutans works through more indirect mechanisms, such as weakening the biofilm structure, but it is ineffective on preformed biofilms. In dual-species biofilms, tyrosol is not sufficiently effective on its own compared to powerful antibiofilm agents such as farnesol or polyphenols, but its effectiveness can be increased through combined approaches. In particular, various studies have reported synergistic effects when combined with antifungal agents such as fluconazole or amphotericin. Additionally, the risk of resistance development with tyrosol is significantly lower compared to antibiotics. Natural compounds such as quercetin and resveratrol, on the other hand, can provide comprehensive antibiofilm effects at lower concentrations than tyrosol and through mechanisms such as inhibiting the QS system or suppressing virulence genes.
This study provides important insights into how tyrosol affects single and dual biofilms and planktonic cultures of C. tropicalis and S. mutans, but it is important to recognize some limitations. Despite the remarkable and promising results, one of the main limitations of the study is the small number of isolates included. On the other hand, the data obtained in vitro do not fully reflect natural biofilm ecosystems. There are many variables such as inoculation rate, incubation time, medium selection, surface type in dual biofilm studies and it is difficult to standardize these parameters. In order to support the data obtained in future research, studies in this field should be expanded with more isolates, the genes leading to resistance should be identified by molecular analysis and our data should be confirmed by in vivo experiments.
On the other hand, clinical strains of the same species can exhibit different resistance profiles in both types. While some microorganisms are resistant to the value recommended in CLSI guidelines, others can tolerate much higher concentrations. Similarly, some may be resistant to one or two antibiotics, whereas others may show resistance to several. Here, serotypic and phenotypic homogeneity differences in S. mutans affect the resistance pattern; in C. tropicalis, clonal structure, mutations, and gene expression levels can create different antifungal susceptibility profiles between strains. Therefore, strains with different resistance spectra must also be studied in detail.
As the first study to evaluate the effects of tyrosol on C. tropicalis–S. mutans biofilms, our findings highlight the potential for therapeutic application in the management of mixed oral biofilms, while underlining the need for further mechanistic and clinical research. Despite these limitations, it can be concluded that tyrosol has an inhibitory effect on oral biofilms and has the potential to be used as a natural alternative candidate for the prevention of dental caries.
Supplementary materials
Table S1Absorbance, cell survival (%) and standard deviation values obtained as a result of 24 h exposure of tyrosol to 3T3 cells at concentration ranges of 1–20 mg/mL.Concentration (mg/mL)OD 450 nmCell survival (%)Standard deviation200.5090.5150.52823.5221.450150.5090.5110.50822.0600.228100.4750.4540.45513.2851.7687.50.4880.4300.44712.1274.45150.4470.4210.4197.3742.3324.50.4520.4380.4309.3851.66240.3980.4070.3992.3160.7363.50.4000.4190.4264.8142.00830.4050.4190.4255.0581.5322.50.3940.3980.3900.9750.5972.40.3810.4150.4132.6202.8482.20.4910.5170.48720.0492.4312.00.5950.6350.39941.3773.6561.80.7330.7790.46067.1544.2051.60.7380.7480.82269.5926.8491.0.6960.7520.82067.1549.2691.20.7550.8250.91881.17012.2061.00.8230.9760.971106.9170.457POZ0.4630.4950.46615.7222.638NEG0.9720.10090.826100.0004.443BLANK0.3680.3780.4200.0004.119
Table S2Absorbance, cell survival (%) and standard deviation values obtained as a result of 48 h exposure of tyrosol to 3T3 cells at concentration ranges of 1–20 mg/mL.Concentration (mg/mL)OD 450 nmCell survival (%)Standard deviation200.4680.4680.4538.0130.680150.5180.5000.48811.7631.185100.4770.4530.4407.4041.4747.50.4390.4150.4284.5830.94350.4570.4270.4245.4171.4334.50.4730.4490.4346.9551.54440.1170.4080.4092.7720.0483.50.4090.4300.4334.2631.02730.4020.4140.4132.8850.5232.50.3990.3960.3931.5710.2362.40.3980.4160.4042.5320.7202.20.4090.4220.4193.5580.5342.00.4140.4130.4374.0061.0661.60.6860.6580.46421.4429.4921.40.7950.10410.102662.8690.7211.20.9040.9440.96553.6542.4331.00.10560.10950.106066.4101.684POZ0.4350.4610.4406.3141.083NEG0.14400.13620.1457100.0003.977BLANK0.3750.3810.3830.0000.327
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arias LS Delbem ACB Fernandes RA Barbosa DB Monteiro DR 2016 Activity of tyrosol against single and mixed-species oral biofilms Journal of Applied Microbiology 120 5 1240 1249 10.1111/jam.13070 26801208 · doi ↗ · pubmed ↗
- 2Azeredo J Azevedo NF Briandet R Cerca N Coenye T 2017 Critical review on biofilm methods Critical Reviews in Microbiology 43 3 313 351 10.1080/1040841 X.2016.1208146 27868469 · doi ↗ · pubmed ↗
- 3Barbosa JO Rossoni RD Vilela SFG De Alvarenga JA Velloso MDS 2016 Streptococcus mutans can modulate biofilm formation and attenuate the virulence of Candida albicans P Lo S One 11 3 e 0150457 10.1371/journal.pone.0150457 26934196 PMC 4774980 · doi ↗ · pubmed ↗
- 4Bourroubey B Chelli N Touil AT Meddah B 2025 Regulation of exopolysaccharide production by Pistacia lentiscus L. to modulate intestinal microbiota in type 2 diabetics Journal of Microbiology, Biotechnology and Food Sciences 15 1 e 11455 10.55251/jmbfs.11455 · doi ↗
- 5Brown GD Denning DW Gow NAR Levitz SM Netea MG 2012 Hidden killers: Human fungal infections Science Translational Medicine 4 165 165rv 13 10.1126/scitranslmed.3004404 23253612 · doi ↗ · pubmed ↗
- 6Chandra J Patel JD Li J Zhou G Mukherjee PK 2005 Modification of surface properties of biomaterials influences the ability of Candida albicans to form biofilms Applied and Environmental Microbiology 71 12 8795 8801 10.1128/AEM.71.12.8795-8801.2005 16332875 PMC 1317330 · doi ↗ · pubmed ↗
- 7CLSI Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard—Third Edition CLSI document M 27 A 3 Wayne, PA Clinical and Laboratory Standards Institute 2008
- 8CLSI Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard – Eighth Edition CLSI document M 07 A 8 Wayne, PA Clinical and Laboratory Standards Institute 2009