Eucommia ulmoides extract attenuates oxidative stress and promotes melanogenesis via Wnt/β-catenin signaling in B16 cells and mice
Xiaojin LIU, Yaqian QIU, Xiaobing LV, Lei CHANG, Tiancheng JI, Yefan GU, Shuyue CHEN

TL;DR
Eucommia ulmoides extract reduces oxidative stress and boosts melanin production in cells and mice by activating a key signaling pathway.
Contribution
The study reveals a novel mechanism by which EUE activates the Wnt/β-catenin pathway to combat oxidative stress and promote melanogenesis.
Findings
EUE reduced ROS levels and improved melanocyte survival under oxidative stress.
EUE activated the Wnt/β-catenin pathway, increasing MITF and melanogenic enzymes.
Topical EUE application protected hair follicles from H2O2-induced depigmentation in mice.
Abstract
Oxidative stress is a major contributor to melanocyte dysfunction and hair graying by impairing key signaling pathways. Eucommia ulmoides bark extract (EUE), rich in antioxidant phytochemicals, has shown potential in combating oxidative damage. This study investigated the protective and promelanogenic effects of EUE under hydrogen peroxide (H2O2)-induced oxidative stress, with a focus on the Wnt/β-catenin signaling pathway. An oxidative stress model was established using B16 cells and a C57BL/6 mouse hair follicle model. EUE significantly improved melanocyte survival and reduced intracellular reactive oxygen species (ROS). Mechanistically, EUE activated the Wnt/β-catenin pathway, leading to upregulation of the microphthalmia-associated transcription factor (MITF) and its downstream melanogenic enzymes (TYR, TRP-1, TRP-2), thereby enhancing tyrosinase activity and restoring melanin…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
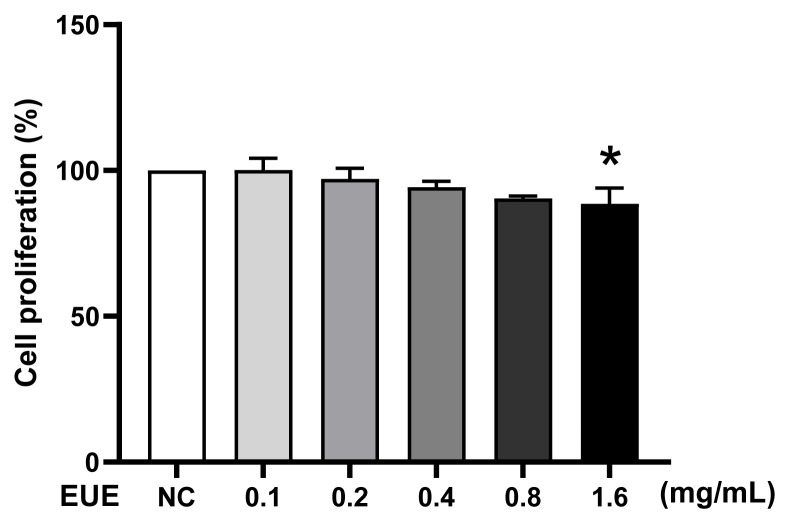 Figure 1
Figure 1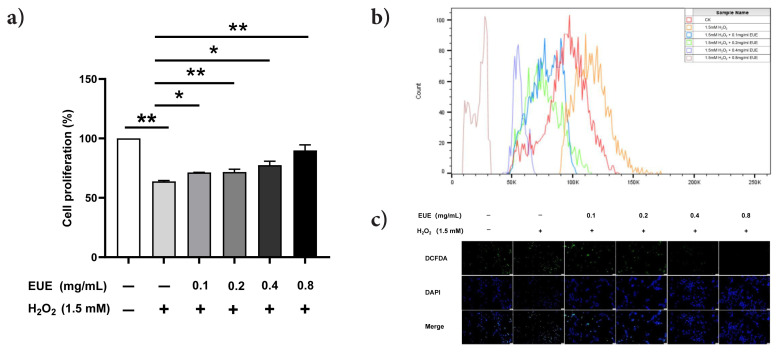 Figure 2
Figure 2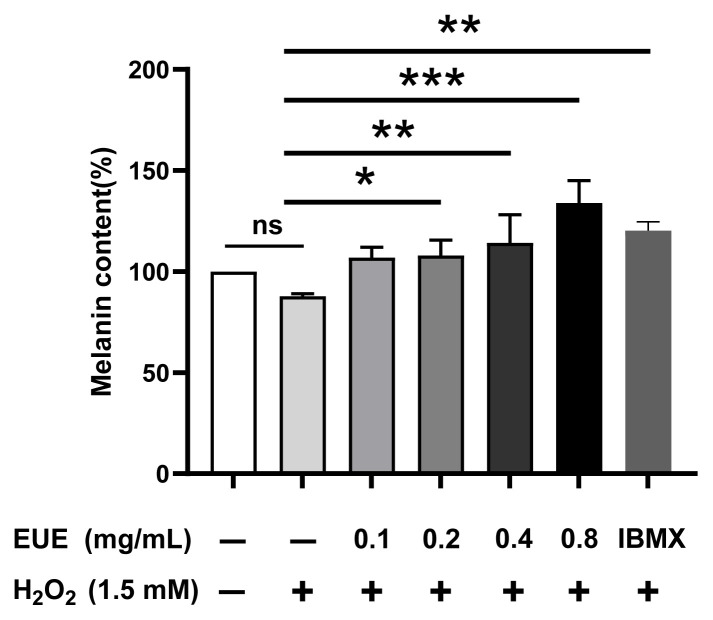 Figure 3
Figure 3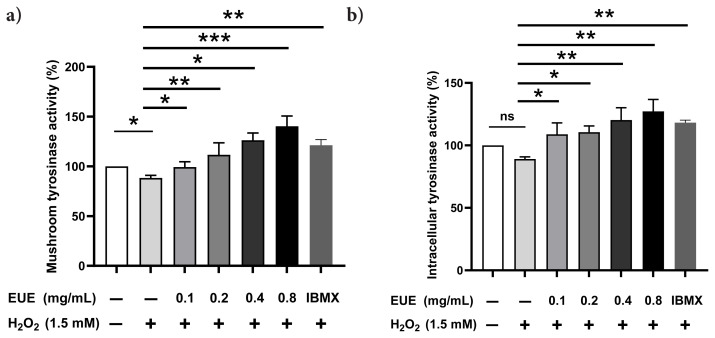 Figure 4
Figure 4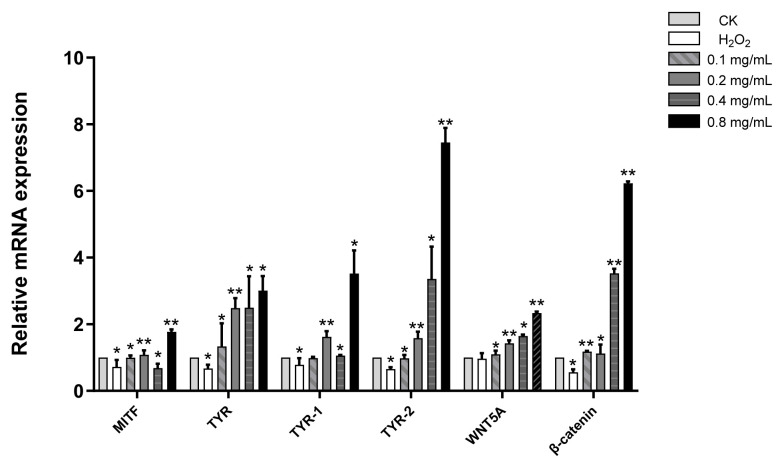 Figure 5
Figure 5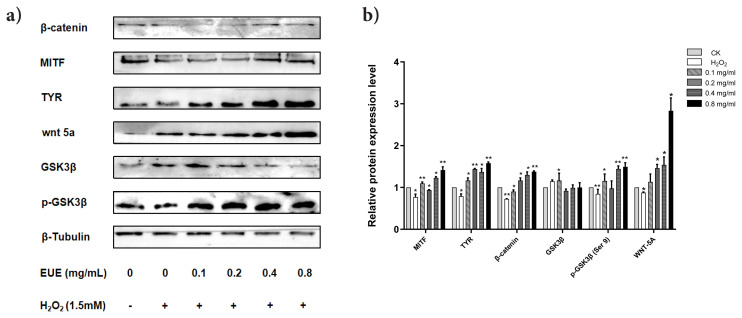 Figure 6
Figure 6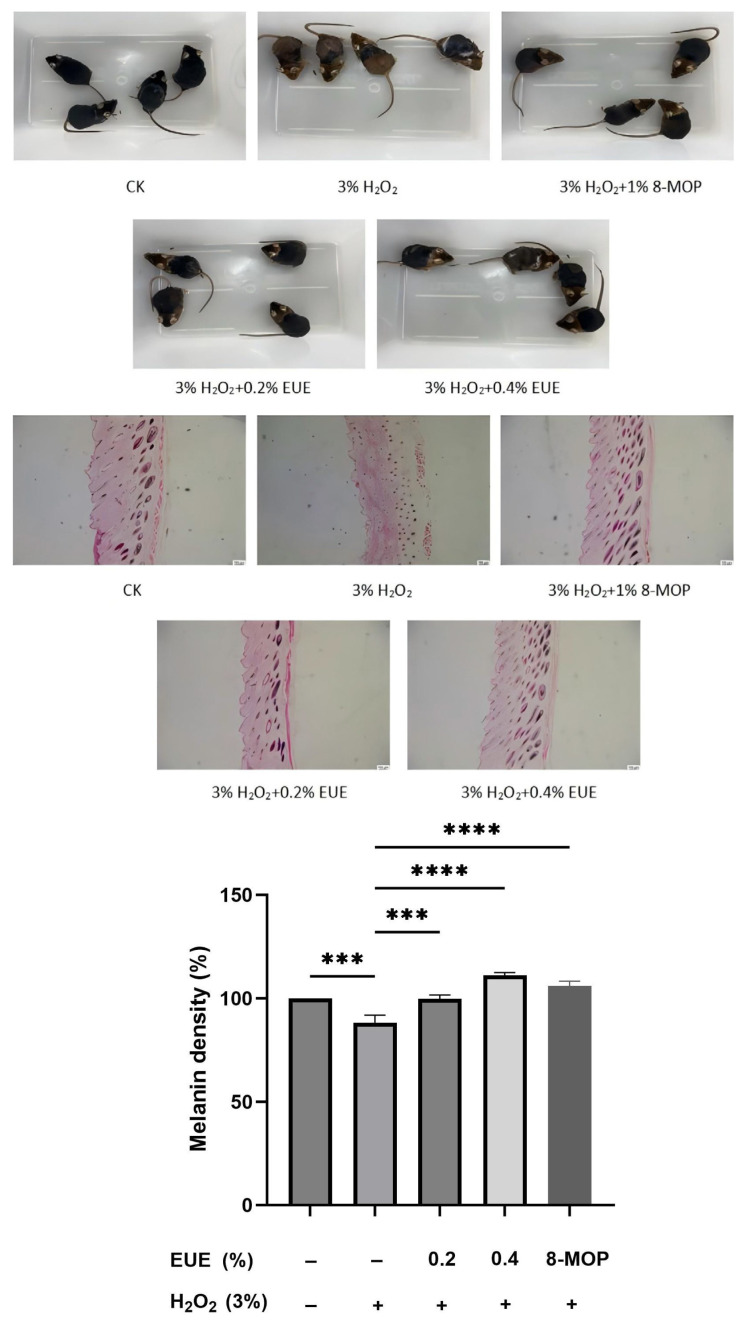 Figure 7
Figure 7- —Wuhu Sci-Tech Program Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHair Growth and Disorders · melanin and skin pigmentation · Natural Compound Pharmacology Studies
Introduction
Thick, dark hair is widely regarded as a symbol of youth and vitality across cultures, while graying hair often marks aging. Currently, no effective medical treatments exist for repigmenting white hair, leaving chemical hair dyes—some containing carcinogenic compounds—as the primary solution (Nishimura et al., 2002; Nishimura et al., 2005; Bolt and Golka, 2007; Wood et al., 2009; Yale et al., 2020; He et al., 2022). In human hair follicles (HFs), melanogenesis is governed by tyrosinase (TYR), tyrosinase-related protein-1 (TRP-1), and tyrosinase-related protein-2 (TRP-2), all regulated by the microphthalmia-associated transcription factor (MITF) (D’Mello et al., 2016). The Wnt/β-catenin signaling pathway plays a critical role in melanocyte differentiation, survival, and pigmentation by modulating MITF and TYR expression (Mulholland et al., 2005; Zang et al., 2019).
The concept of oxidative stress, defined as an imbalance between reactive oxygen species (ROS) and antioxidant defenses, was first introduced by Helmut Sies in 1985 (Sies, 1985; Sies et al., 2017). It contributes to melanocyte apoptosis and hair graying, and has been implicated in numerous degenerative conditions. Factors such as psychoemotional stress, inflammation, and environmental challenges elevate oxidative stress, leading to melanocyte apoptosis and hair depigmentation, often termed the “free radical theory of aging” (Melov, 2002; Arck et al., 2006; Jadkauskaite et al., 2017). Hydrogen peroxide (H_2_O_2_) accumulation exacerbates this process, and its reduction has been shown to mitigate cell damage and support hair health (Forman and Zhang, 2021). Importantly, oxidative stress can suppress Wnt/β-catenin signaling, reducing β-catenin stability, impairing MITF activation, and ultimately downregulating melanogenesis, thereby accelerating pigment loss from the follicular niche (Jadkauskaite et al., 2017; Zang et al., 2019).
Eucommia ulmoides, documented in the Chinese Pharmacopoeia since the Han Dynasty over 2000 years ago, is a medicinal plant traditionally used in Asia to treat aging-related conditions, including hypertension, diabetes, and osteoporosis (Zhou et al., 2009; He et al., 2014; Huang et al., 2021). Eucommia ulmoides bark extract (EUE) is rich in bioactive compounds with antioxidant properties. This study aimed to determine whether EUE can protect melanocytes from H_2_O_2_-induced oxidative stress and restore pigmentation by modulating the Wnt/β-catenin pathway. Using both B16 murine melanoma cells and a mouse hair follicle model, we assessed melanin production, TYR activity, and key melanogenic signaling molecules, providing mechanistic evidence for EUE as a candidate therapeutic for oxidative stress-induced hair depigmentation.
Materials and methods
2.1. Biological materials, chemicals, and reagents
B16 melanoma cells were obtained from Sciencell Research Laboratories (Carlsbad, CA, USA). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM; Gibco, Thermo Fisher Scientific, USA) supplemented with 10% fetal bovine serum (FBS; Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37 °C in a humidified atmosphere containing 5% CO_2_. EUE was sourced from Shanghai Zhina Biotechnology Co., Ltd. CCK-8 was purchased from Yeasen (Shanghai, China). 3-Isobutyl-1-methyl-xanthine (IBMX) was purchased from BOSF Biotechnology Co., Ltd (Hefei, China). L-3,4-dihydroxyphenylalanine (L-DOPA) was purchased from Hefei Bomei Biotechnology Co., Ltd. Enhanced chemiluminescence (ECL) reagent was obtained from Beijing Labgic Technology Co., Ltd. Poly-vinylidene fluoride (PVDF) membranes were supplied by Beyotime Biotech Inc. Antibodies were purchased from Proteintech Group, Inc. Male C57BL/6 mice were purchased from Henan Sikebas Biotechnology Co., Ltd., China. The Leica DMi8 inverted microscope was purchased from Leica Microsystems (Germany), the BD FACSCanto II flow cytometer was purchased from Becton Dickinson (USA), and the Spark 10M multimode microplate reader was purchased from Tecan Group Ltd. (Männedorf, Switzerland; Asset ID: 17104551).
2.2. Cell viability assay
Cell viability was assessed using the CCK-8 kit. B16 melanoma cells were seeded in 96-well plates at a density of 1 × 10^4^ cells/well and incubated overnight. Cells were first exposed to 1.5 mM H_2_O_2_ for 0.5 h in the dark at 37 °C, and then treated with EUE (0, 0.1, 0.2, 0.4, or 0.8 mg/mL) for 48 h. Optical density (OD) was measured at 450 nm using a microplate reader. Each experiment was performed in triplicate. The doses and exposure times of EUE and H_2_O_2_ were determined based on our lab’s preliminary cytotoxicity tests, confirming moderate cell viability (~80%) under oxidative stress.
2.3. Intracellular ROS measurement
Intracellular ROS levels were quantified using 2,7′-dichlorodihydrofluorescein diacetate (DCFH_2_-DA) staining, as described by Hseu (Hseu et al., 2019). B16 cells were treated with 1.5 mM H_2_O_2_ to induce oxidative stress, followed by the addition of EUE (0.1, 0.2, 0.4, or 0.8 mg/mL). Cells were incubated with DCFH_2_-DA (10 μM) at 37 °C for 30 min in the dark, counterstained with DAPI for 5 min, and analyzed using an inverted fluorescence microscope (200 × magnification) and a flow cytometer.
2.4. Measurement of intracellular melanin content
Intracellular melanin content in melanoma cells was measured following established protocols (Huang et al., 2020; Chen et al., 2021; Liu et al., 2024). Cells were exposed to 1.5 mM H_2_O_2_ for 0.5 h to induce oxidative stress. Subsequently, cells were treated for an additional 48 h with either EUE (0.1, 0.2, 0.4, or 0.8 mg/mL) or IBMX (35 μM), a protein kinase inhibitor. A blank control was included without H_2_O_2_, EUE, or IBMX. Posttreatment, cell pellets were collected and solubilized in 1 M NaOH at 60 °C for 60 min. Melanin levels were quantified spectrophotometrically at 405 nm, and absorbance was measured using a multimode microplate reader.
2.5. Assay of mushroom TYR and intracellular TYR activity
Mushroom TYR and intracellular TYR activities were assayed following previously described methods (Huang et al., 2020; Chen et al., 2021; Liu et al., 2024). Briefly, melanoma cells were exposed to 1.5 mM H_2_O_2_ for 0.5 h to simulate oxidative stress, followed by treatment with EUE (0.1, 0.2, 0.4, or 0.8 mg/mL) or IBMX (35 μM) for 48 h. A blank control was included without H_2_O_2_, EUE, or IBMX. Posttreatment, cell extracts (100 μL) were mixed with freshly prepared 0.1% L-DOPA solution (in PBS), incubated at 37 °C for 30 min, and the absorbance was measured at 490 nm to assess TYR activity.
2.6. Gene expression analysis
Quantitative real-time polymerase chain reaction (qRT–PCR) was performed to evaluate the expression levels of melanogenesis-related genes, including Wnt5a, β-catenin, MITF, TYR, TRP-1, and TRP-2. All experimental methods, primers, and instruments were adapted from the report by Liu et al. (Liu et al., 2022; Liu et al., 2024).
2.7. Western blot analysis
B16 cells were lysed in PBS containing proteinase inhibitors at 4 °C for 20 min. Proteins (30 μg) were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and transferred to a PVDF membrane. The membrane was blocked with 5% nonfat milk in PBST for 1 h, then incubated overnight at 4 °C with primary antibodies against MITF, TRP1, TRP2, TYR, GSK3β, p-GSK3β, Wnt5a, β-catenin, and β-tubulin. After washing, the membrane was incubated with horseradish peroxidase (HRP)-conjugated goat antimouse secondary antibody (1:7500) at room temperature for 2 h. Protein bands were visualized using ECL reagent and quantified with Multi Gauge 3.0 software, normalized to glyceraldehyde-3-phosphate dehydrogenase (β-tubulin) levels.
Primary antibodies were obtained from Proteintech Group (Chicago, IL, USA): MITF (catalog number 13092-1-AP, 1:1500), TYR (catalog number 31291-1-AP, 1:1000), Wnt5a (catalog number 55184-1-AP, 1:1500), β-catenin (catalog number 51067-2-AP, 1:6000), GSK-3β (catalog number 82061-1-RR, 1:10,000), and p-GSK-3β (catalog number 67558-1-Ig, 1:5000). β-tubulin (catalog number 10094-1-AP, 1:6000) was used as the internal control. The secondary HRP-conjugated goat antimouse IgG antibody (catalog number SA00001-1, 1:7500) was used.
2.8. In vivo evaluation of EUE effects on pigmentation under H2O2-induced stress
The effects of EUE on hair pigmentation under oxidative stress were evaluated using C57BL/6 mice, with all procedures approved by the Academic Ethics Committee of Anhui Normal University (Permit: AHNU-ET2022064). Twenty-seven-week-old healthy male mice (23~25 g) were housed in a temperature-controlled room with a 12-h light/dark cycle and provided food and water ad libitum. The dorsal skin (2 cm × 2 cm) of each mouse was shaved to induce the hair follicle transition from telogen to anagen. Mice were randomly divided into five groups (n = 4 per group):
Negative control: 200 μL of 3% H_2_O_2_ applied at 9 am and 4 pm daily.
Blank control (CK): 200 μL of water applied at 9 am daily.
Three to five treatment groups: 200 μL of 3% H_2_O_2_ applied at 9 am, followed by 200 μL of 0.2% or 0.4% EUE, or 1% 8-MOP (positive control) at 4 pm daily.
Treatments were administered topically to the shaved area once daily for 21 days. Hair pigmentation and growth were subsequently assessed via histological analysis. Following hematoxylin and eosin (H&E) staining, histological sections were imaged under a light microscope. To semiquantitatively assess melanin density, these images were analyzed using ImageJ software (National Institutes of Health, USA)1. Briefly, for each hair follicle cross-section, the area containing melanin was manually selected as the region of interest (ROI). The integrated density (IntDen) of the ROI was measured. The average IntDen value of the blank control group was set as 100%, and the data from all other groups were normalized and expressed as a percentage relative to the control.
2.9. Statistical analysis
Data were expressed as mean ± standard deviation (SD). Statistical significance was assessed using one-way analysis of variance (ANOVA) followed by Dunnett’s test for pairwise comparisons. Significance levels were defined as *p < 0.05, **p < 0.01, ***p < 0.001 and ***** p < 0.0001.
Results
3.1. Effect of EUE on B16 melanoma cell viability
The cytotoxicity of EUE on B16 melanoma cells was evaluated using the CCK-8 assay (Figure 1). Cells were treated with EUE (0.1, 0.2, 0.4, 0.8, or 1.6 mg/mL) for 24 h or 48 h. After 24 h, cell viability relative to the negative control (NC) was 97.34%, 94.61%, 92.90%, 89.76%, and 84.78% (*p < 0.05 at 1.6 mg/mL), with similar trends observed at 48 h. Higher concentrations (≥1.6 mg/mL) reduced viability below 85%, indicating mild cytotoxicity. Thus, EUE concentrations below 1.6 mg/mL (i.e. ≤0.8 mg/mL) were selected for subsequent experiments.
3.2. EUE attenuates H2O2-induced oxidative damage in B16 melanoma cells
To assess EUE’s protective effect against oxidative stress, B16 melanoma cells were pretreated with EUE (0.1–0.8 mg/mL) for 48 h, followed by 1.5 mM H_2_O_2_ exposure. The CCK-8 assay (Figure 2A) showed that EUE mitigated H_2_O_2_-induced cytotoxicity, with 0.8 mg/mL yielding the greatest protection. Flow cytometry (Figure 2B) revealed elevated intracellular ROS levels after H_2_O_2_ treatment compared with the control (CK), and EUE reduced these levels in a dose-dependent manner. Fluorescence microscopy with DCFH_2_-DA staining (Figure 2C) revealed strong green fluorescence intensity after H_2_O_2_ exposure, indicating elevated ROS levels. EUE treatment reduced fluorescence intensity in a dose-dependent manner, suggesting effective ROS scavenging.
3.3. Effect of EUE on melanin content in B16 melanoma cells
EUE treatment dose-dependently increased melanin content in B16 melanoma cells under 1.5 mM H_2_O_2_-induced oxidative stress (Figure 3). After 24 h, melanin levels at EUE concentrations of 0.1, 0.2, 0.4, and 0.8 mg/mL were 101.02%, 110.46% (*p < 0.05), 124.81% (**p < 0.01), and 132.88% (***p < 0.001) relative to the H_2_O_2_-treated group, respectively. Compared with the positive control IBMX (35 μM), 0.8 mg/mL EUE yielded 120.20% (**p < 0.01) of the melanin content, while H_2_O_2_ alone reduced it to 89.03% of the untreated baseline. These results indicate that EUE (>0.1 mg/mL) counteracted H_2_O_2_-induced depigmentation, with higher doses (0.4 and 0.8 mg/mL) surpassing IBMX in stimulating melanogenesis.
3.4. Effects of EUE on mushroom and intracellular TYR activity in B16 melanoma cells
EUE treatment significantly enhanced both mushroom and intracellular TYR activities in B16 melanoma cells under 1.5 mM H_2_O_2_-induced oxidative stress (Figure 4). For mushroom TYR (Figure 4A), 24 h treatment with EUE at 0.1, 0.2, 0.4, and 0.8 mg/mL increased activity to 105.40% (*p < 0.05), 125.22% (**p < 0.01), 134.58% (*p < 0.05), and 152.21% (***p < 0.001) relative to the H_2_O_2_-treated group, respectively, compared with 121.41% (**p < 0.01) for IBMX (35 μM). H_2_O_2_ alone reduced activity to 91.29% (*p < 0.05) of the negative control (NC). For intracellular TYR (Figure 4B), EUE at the same concentrations elevated activity to 117.59% (*p < 0.05), 115.81% (*p < 0.05), 124.97% (***p < 0.001), and 129.93% (****p < 0.0001) relative to the H_2_O_2_ group, compared with 115.81% (**p < 0.01) for IBMX, with H_2_O_2_ reducing activity to 90.21% (*p < 0.05) of NC. IBMX served as a positive pigmentation standard, while H_2_O_2_ acted as a depigmentation control. These results demonstrate that EUE (>0.1 mg/mL) counteracted H_2_O_2_-induced inhibition, with the highest dose (0.8 mg/mL) outperforming IBMX in stimulating both TYR activities.
3.5. EUE upregulates melanin synthesis-related gene expression in B16 cells
In the anagen phase of HFs, melanocytes produce melanin and transfer it to keratinocytes via melanosomes, a process regulated by MITF and the Wnt/β-catenin pathway (Paus et al., 2013; Huang et al., 2020). To assess EUE’s effect on this pathway, mRNA levels of six melanin synthesis genes in B16 melanoma cells were measured by qRT–PCR under 1.5 mM H_2_O_2_ stress (Figure 5). After 25 min of H_2_O_2_ exposure, messenger ribonucleic acid (mRNA) levels decreased to MITF (0.72), TYR (0.67), TRP-1 (0.78), TRP-2 (0.65), Wnt5a (0.97), and β-catenin (0.56) relative to the blank control. Treatment with EUE (0.1, 0.2, 0.4, or 0.8 mg/mL) for 24 h dose-dependently increased the mRNA levels compared with the negative control (NC), with fold changes peaking at 0.8 mg/mL: MITF (1.77), TYR (3.00), TRP-1 (3.52), TRP-2 (7.45), Wnt5a (2.33), and β-catenin (6.23). These findings suggest that EUE promotes melanogenesis by activating the Wnt/β-catenin signaling pathway under oxidative stress.
3.6. EUE enhances melanogenesis-related and signalling protein expression in B16 cells
Western blot analysis showed that EUE treatment under 1.5 mM H_2_O_2_ stress increased the expression of MITF, TYR, β-catenin, GSK-3β, p-GSK-3β, and Wnt-5a in B16 melanoma cells (Figure 6). After 24 h treatment with EUE (0, 0.1, 0.2, 0.4, or 0.8 mg/mL), protein levels relative to the control were MITF (0.77, 1.10, 0.94, 1.23, 1.42), TYR (0.79, 1.17, 1.44, 1.37, 1.58), β-catenin (0.73, 0.90, 1.17, 1.30, 1.38), GSK-3β (1.05, 1.12, 0.92, 0.99, 1.00), p-GSK-3β (0.85, 1.16, 0.98, 1.45, 1.49), and Wnt-5a (0.88, 1.14, 1.46, 1.54, 2.83). The stimulatory effect was most pronounced at 0.8 mg/mL, with significant upregulation of MITF, TYR, β-catenin, GSK-3β, p-GSK-3β, and Wnt-5a, while GSK-3β levels remained unchanged. These findings suggest that EUE promotes melanogenesis via the Wnt/β-catenin pathway under oxidative stress, with minimal impact on GSK-3β signalling. Protein expression patterns in Figure 6A and Supplementary Figure were consistent with the corresponding densitometric analysis in Figure 6B, and the trends were reproducible across three independent experiments.
3.7. Hair growth and histological analysis in EUE-treated depilated mice
Hair growth and pigmentation in EUE-treated C57BL/6 mice were evaluated via photography under 3% H_2_O_2_-induced oxidative stress (Figure 7A). The H_2_O_2_ group exhibited yellow hairs in pigment islands, indicating depigmentation, whereas the EUE/H_2_O_2_ and 8-MOP (positive control) groups showed black hairs, suggesting EUE counteracted H_2_O_2_-induced depigmentation. H&E staining of dorsal skin sections further revealed that H_2_O_2_ reduced melanin content in HFs and shafts, while EUE treatment increased melanin staining compared with the H_2_O_2_-treated controls (Figure 7B). To provide quantitative support for these histological observations, we performed a semiquantitative analysis of melanin density in hair follicle cross-sections using ImageJ. The results demonstrated that H_2_O_2_ treatment significantly reduced follicular melanin density compared with the blank control group (**p < 0.0001). Importantly, topical application of both 0.2% and 0.4% EUE effectively attenuated this H_2_O_2_-induced reduction in a dose-dependent manner (***p < 0.001 and ****p < 0.0001, respectively), with the higher concentration of EUE (0.4%) even elevating melanin levels above baseline. The positive control, 8-MOP, also exhibited a significant restorative effect. These quantitative data, presented in Figure 7C, strongly corroborate our visual findings and confirm the potent protective effect of EUE against oxidative stress-induced hair follicle depigmentation. By day 21, EUE-treated mice displayed faster hair growth and thicker skin than the H_2_O_2_ group, trends consistent with enhanced follicular health. These findings indicate EUE’s potential to promote hair growth and pigmentation, warranting further investigation into its efficacy for the treatment of androgenetic alopecia.
Discussion
Oxidative stress is a fundamental driver of hair graying through ROS-mediated damage to melanocytes and disruption of regulatory networks that sustain pigmentation (Nishimura et al., 2002; Van Neste and Tobin, 2004; Nishimura et al., 2005; Paus et al., 2013; Liu et al., 2022; Zhang et al., 2022). In this study, EUE protected B16 melanocytes from H_2_O_2_-induced injury and preserved follicular pigmentation in a mouse model. Beyond its antioxidant capacity, EUE increased melanin content and TYR activity, and upregulated key melanogenic regulators, including MITF, TRP-1, and TRP-2.
Notably, EUE elevated β-catenin levels and increased GSK-3β phosphorylation, a modification known to reduce β-catenin degradation. This suggests that EUE may inhibit β-catenin turnover, thereby sustaining melanocyte activity. This is in agreement with reports showing that GSK-3β inhibition promotes melanogenesis (Zang et al., 2019), and it supports a mechanistic model in which EUE stabilizes β-catenin and promotes MITF-dependent transcription of melanogenesis genes under oxidative stress.
These findings extend prior reports that focused primarily on the antioxidative properties of E. ulmoides by providing functional and mechanistic evidence for modulation of the Wnt/β-catenin-GSK-3β axis under oxidative challenge (Cheng et al., 2022; Park et al., 2025). Regarding phytochemical composition, EUE contains multiple bioactive constituents, including geniposidic acid, lignans, and flavonoids, which have been reported to exert antioxidant and melanogenesis-modulating effects. These components may act synergistically to confer the observed protective outcomes. The dual action—ROS attenuation and signaling restoration—suggests EUE may both reduce oxidative burden and reestablish propigmentary signaling, which is particularly relevant for maintaining melanocyte stem cell function within the follicular niche.
Limitations of the study include the modest in vivo sample size (n = 4 per group), which may reduce statistical power. The present findings should therefore be interpreted as preliminary evidence. Nonetheless, consistent trends were observed across groups, supporting the reliability of the outcomes. Future studies with larger cohorts are warranted to validate these results and enhance the robustness of the conclusions. Additionally, the complex, multicomponent nature of EUE presents both a challenge and an opportunity for future research. While our findings are promising, further validation in human scalp melanocytes and hair follicle organ cultures is necessary to assess translational potential. Additionally, comprehensive safety profiling, skin penetration studies, and formulation optimization will be essential before considering clinical applications. Future studies should (i) isolate and characterize bioactive constituents, (ii) validate effects in primary human scalp melanocytes and human hair follicle organ cultures, and (iii) employ pathway-specific perturbations to establish causality. Additionally, formulation development, penetration studies, and long-term safety profiling will be required before clinical translation. Overall, our data provide a mechanistic basis for further development of EUE-derived strategies to mitigate oxidative stress-related hair depigmentation.
Supplementary material
Supplementary FigureOriginal uncropped western blot images corresponding to Figure 6A. This figure shows the uncropped original western blot bands for MITF, YR, β-catenin, GSK3β, p-GSK3β, Wnt5a, and β-tubulin.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arck PC Overall R Spatz K Liezman C Handjiski B 2006 Towards a “free radical theory of graying”: melanocyte apoptosis in the aging human hair follicle is an indicator of oxidative stress induced tissue damage The FASEB Journal 20 9 1567 1569 10.1096/fj.05-4039 fje 16723385 · doi ↗ · pubmed ↗
- 2Bolt HM Golka K 2007 The debate on carcinogenicity of permanent hair dyes: new insights Critical Reviews in Toxicology 37 6 521 536 10.1080/10408440701385671 17661215 · doi ↗ · pubmed ↗
- 3Chen S Hseu Y Gowrisankar YV Chung Y Zhang Y 2021 The anti-melanogenic effects of 3-O-ethyl ascorbic acid via Nrf 2-mediated alpha-MSH inhibition in UVA-irradiated keratinocytes and autophagy induction in melanocytes Free Radical Biology and Medicine 173 151 169 10.1016/j.freeradbiomed.2021.07.030 34314818 · doi ↗ · pubmed ↗
- 4Cheng S Jia H Zhang Y Zhou J Chen X 2022 Geniposidic Acid from Eucommia ulmoides Oliver Staminate Flower Tea Mitigates Cellular Oxidative Stress via Activating AKT/NRF 2 Signaling Molecules 27 23 8568 10.3390/molecules 27238568 36500666 PMC 9739628 · doi ↗ · pubmed ↗
- 5D’Mello SAN Finlay GJ Baguley BC Askarian-Amiri ME 2016 Signaling Pathways in Melanogenesis International Journal of Molecular Sciences 17 7 10.3390/ijms 17071144 PMC 496451727428965 · doi ↗ · pubmed ↗
- 6Forman HJ Zhang H 2021 Targeting oxidative stress in disease: promise and limitations of antioxidant therapy Nature Reviews Drug Discovery 20 9 689 709 10.1038/s 41573-021-00233-1 34194012 PMC 8243062 · doi ↗ · pubmed ↗
- 7He L Michailidou F Gahlon HL Zeng W 2022 Hair Dye Ingredients and Potential Health Risks from Exposure to Hair Dyeing Chemical Research in Toxicology 35 6 901 915 10.1021/acs.chemrestox.1c 00427 35666914 PMC 9214764 · doi ↗ · pubmed ↗
- 8He X Wang J Li M Hao D Yang Y 2014 Eucommia ulmoides Oliv.: ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine Journal of Ethnopharmacology 151 1 78 92 10.1016/j.jep.2013.11.023 24296089 · doi ↗ · pubmed ↗