Efflux pump-associated antimicrobial resistance genes in Staphylococcus spp. from dairy and meat samples
Meryem Burcu KÜLAHCI, Sumru ÇITAK

TL;DR
This study examines antibiotic resistance genes in Staphylococcus bacteria from dairy and meat, finding that efflux pumps play a key role in resistance to antibiotics and biocides.
Contribution
The study identifies and validates the role of efflux pump genes in antimicrobial resistance among Staphylococcus spp. in food samples.
Findings
Reserpine reduced MIC values in cefuroxime-resistant isolates, indicating efflux pump involvement.
Common efflux pump genes like smr, efrA, and mdeA were frequently detected in the isolates.
Resistance to tetracycline and cefuroxime was most prevalent among the tested isolates.
Abstract
This study aimed to investigate the phenotypic resistance and distribution of efflux pump-associated antimicrobial resistance genes in Staphylococcus spp. isolated from dairy and meat samples. Antimicrobial resistance in foodborne bacteria increases with antibiotic exposure and biocides, particularly through efflux mechanisms. Thus, monitoring potential genetic reservoirs in the food chain is very important. A total of 132 dairy and meat samples were collected for the study, and Staphylococcus spp. were isolated using Mannitol salt phenol red agar. Antimicrobial susceptibility was evaluated using the Clinical and Laboratory Standards Institute’s microdilution method. Twenty-six resistant isolates were identified by 16S rDNA sequencing. The effect of reserpine on MIC values was evaluated using microdilution tests to assess the role of efflux pumps in antibiotic resistance and biocide…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
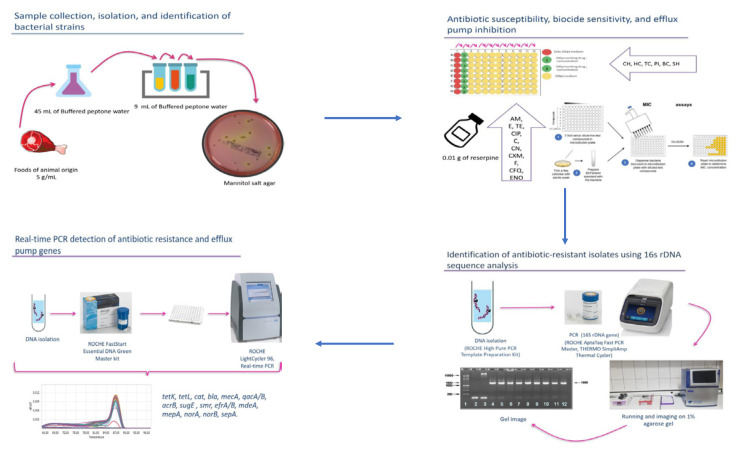 Figure 1
Figure 1- —Scientific Research Projects of the Gazi University
- —Republic of Türkiye Ministry of Agriculture and Forestry, General Directorate of Agricultural Research and Policies, Department of Research and Development (R&D)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Antibiotic Resistance in Bacteria · Antimicrobial agents and applications
Introduction
The development of antimicrobial resistance in bacteria is a global challenge. It is largely driven by the inappropriate use of antibiotics for treatment or preventive purposes in animal infections and their use as productivity enhancers in animal nutrition. This means that antibiotics enter the food chain, and the use of related antibiotics in human medicine trigger defense mechanisms in bacteria, ultimately leading to the development of antibiotic resistance (Lange and Brokking, 2005; Yılmaz et al., 2016; Abbas et al., 2024). Studies have shown that colonization of pathogenic or normal bacteria carrying these resistance genes to the animal and human microbiome through food is an increasing threat to animal and human health (Salam et al., 2023).
Staphylococci are among the most commonly isolated contaminant bacteria from various food samples. Staphylococcus aureus causes various diseases, especially mastitis in animals and food poisoning in humans (Yurdakul et al., 2013). Although coagulase-negative staphylococci are considered commensal and contaminant bacteria, they are known to have pathogenic effects in humans (Rodrigues et al., 2017).
Cleaning and disinfection are of great importance to prevent microorganism contamination of food and the resulting foodborne infections (Pakdel et al., 2023). Biocides are used during raw food production. They are used to sterilize food production facilities and equipment, thereby inactivating bacteria that cause food spoilage (Marquez et al., 2017). There are concerns about whether the widespread use of biocides, and other antimicrobial agents, is driving the development resistance against them (Coombs et al., 2023). Bacterial resistance to biocides was detected in the 1950s and 1960s. This was when biocides started to be used for bacterial sanitation in the food industry. Although studies on biocide resistance are not as numerous as those on antibiotic resistance, it has become an issue of great importance in recent years (Lavilla Lerma et al., 2013).
Biocides used in the food industry are disinfectants containing chlorine, cationic quaternary ammonium compounds, alcohol-based disinfectants, biguanides, iodinated compounds, hexachlorophene, and triclosans (Kampf, 2018). However, the widespread and unconscious use of biocides in this sector leads to the acquisition of resistance genes by microorganisms or the adaptation of initially sensitive microorganisms, as with antibiotics. In addition, there is a growing danger of cross-resistance between antibiotics and biocides (Larsson and Flach, 2022).
One of the mechanisms known to cause antimicrobial resistance in bacteria is the overexpression of efflux pump protein genes. Efflux pumps play an important role in antimicrobial resistance. Recently, a growing number of multidrug and drug-specific efflux pumps have been characterized in different kinds of bacteria. There are 5 families of efflux pump proteins: the ATP-binding cassette (ABC) family, the multidrug and toxic compound extrusion (MATE) family, the major facilitator superfamily (MFS), the resistance-nodulation-division (RND) family, and the small multidrug resistance (SMR) family (Huang et al., 2022).
To prevent efflux pump-based resistance and to increase pump sensitivity, efflux pump inhibitors are an interesting class of compounds in the fight against resistance. Reserpine is an antihypertensive herbal efflux pump inhibitor known to inhibit P-glycoprotein (Sun et al., 2014; Aygül, 2015). Studies show that antibiotic resistance can be prevented by using efflux pump inhibitors (Fernandes-Fuentes et al., 2014; Sezgin, 2025).
In aim of the present study was to determine the status of antibiotic and biocide tolerance in Staphylococcus spp., in addition to evaluating the presence of reserpine-sensitive efflux pump and genes associated with antimicrobial resistance and efflux pumps. We focused on isolates obtained from dairy and meat samples collected from diverse markets within Ankara, Türkiye.
Materials and methods
2.1. Sample collection, isolation, and identification of bacterial strains
We purchased 132 dairy and meat samples, including 60 cheese samples, 13 raw milk samples, 18 chicken samples, and 41 minced meat samples, from different markets in Ankara, Türkiye. Approximately 5 g or 5 mL of each sample were mixed with 45 mL of sterile buffered peptone water (BPW, Merck, Darmstadt, Germany, catalog number: 107228) and homogenized. The bacterial suspensions were serially diluted in sterile BPW solution and spread on Mannitol salt phenol-red agar (Merck, catalog number: 105404) plates (Turkish Food Codex Microbiological Criteria Regulation, 2011)1. The plates were incubated aerobically at 37 °C for 48 h. From each plate, typical pink and yellow staphylococci colonies were isolated and cultured. The preliminary identification was carried out using basic biochemical tests. Isolates were stored in 20% glycerol at −80 °C (Chajecka-Wierzchowska et al., 2015).
2.2. Antibiotic susceptibility test
Staphylococcus isolates were analyzed for antimicrobial resistance using the broth microdilution method according to the Clinical and Laboratory Standards Institute guidelines (Clinical and Laboratory Standards Institute, 2018) for the following antibiotic agents obtained from Oxoid (Wesel, Germany): ampicillin, erythromycin, tetracycline, ciprofloxacin, chloramphenicol, gentamicin, cefuroxime, nitrofurantoin, cefquinome, and enrofloxacin. Staphylococcus epidermidis ATCC 35984 and S. aureus ATCC 29213 were used as control strains.
2.3. Identification of antibiotic-resistant isolates
Twenty-six antibiotic-resistant isolates (Table 1) were genetically identified by 16S rDNA sequence analysis. Total genomic DNA was extracted from pure culture using the optimized protocol of the commercial Roche High Pure PCR Template Preparation kit (Basel, Switzerland). The 16S rDNA coding region sequence was selected and amplified by PCR. The 20 μL reaction mixture included template DNA, forward primer, reverse primer, water, and Roche AptaTaq Fast PCR Master. 16S-F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 16S-R (5′-ATGGTACCGTGTGACGGGCGGTGTGTA-3′) universal primers were used to amplify the 16S-rDNA gene. Sequence analysis was performed by Macrogen (Amsterdam, Netherlands) (Lange et al., 2015).
2.4. Biocide tolerance and efflux pump inhibition
Minimum inhibitory concentrations (MICs) were determined by the broth microdilution method in 96-well microtiter plates according to the recommendations from the Clinical and Laboratory Standards Institute (2018). The biocides used for these assays—sodium hypochlorite (active chlorine 10–15%), povidone-iodine, hexachlorophene [2,2′-methylenebis(3,4,6-trichlorophenol)], chlorhexidine, benzalkonium chloride, and triclosan—were obtained from Sigma-Alrich (Madrid, Spain). Briefly, serial 2-fold dilutions of each substance were incubated with bacterial suspensions adjusted to 5 × 10^5^ CFU/mL in Mueller–Hinton broth (Merck, catalog number: 110293). Microtiter plates were incubated at 37 °C, and visual readings were taken after 24–48 h of incubation. The effect of the efflux pump inhibitor reserpine (Sigma-Aldrich, Madrid, Spain) on antibiotic and biocide MIC values was evaluated by adding reserpine to the medium at a concentration of 25 μg/mL (Fernandes-Fuentes et al., 2014). A ≥4-fold decrease in MIC in the presence of reserpine suggested that the resistance could be attributed to a reserpine-sensitive efflux pump (Gibbons and Udo, 2000).
2.5. Real-time PCR detection of antibiotic resistance and efflux pump genes
The presence of efflux pumps and antibiotic resistance genes was determined by real-time PCR amplification using the LightCycler system (Roche). Antibiotic resistance primers for tetK and tetL for tetracycline, bla and mecA for β-lactam, cat for chloramphenicol resistance, and efflux pump primers qacA/B, acrB, sugE, smr, efrA, efrB, mdeA, mepA, norA, and sepA were designed using information obtained from the NCBI PubMed database2 for this study. norE and norB primer sequences were taken from Swick et al. (2011) and Patel et al. (2010), respectively (Table 2). The Roche FastStart Essential DNA Green Master kit was used for the experiment. The study used primer pairs for the targeted gene region and SYBR Green dye on the LightCycler 96 (Roche Diagnostics, Mannheim, Germany).
Results
From 132 food samples, 77 Staphylococcus spp. isolates were recovered (Table 1). Antibiotic resistance to one or more antibiotic was found in 26 isolates by the MIC method (Table 3). Isolates were sensitive to the erythromycin, ciprofloxacin, gentamicin, cefquinome, and enrofloxacin antibiotics. Of the 26 resistant isolates, 18 (69.2%) were resistant to tetracycline, 10 (38.5%) to cefuroxime, three (11.5%) to chloramphenicol, two (7.7%) to nitrofurantoin, and one (3.9%) to ampicillin (Table 3). These isolates were selected for genotypic identification of antibiotic resistance and efflux pump genes (Table 3). For biocide tolerance, 12 different concentrations were tested for all biocides. Unlike other biocides, sodium hypochlorite and povidone-iodine concentrations were prepared using the doses used in commercial products (Table 4). MIC ranges of six biocides are given in Table 4.
The efflux pump inhibitor reserpine reduced the MIC value between 4- and 256-fold in all cefuroxime-resistant bacteria (Table 3). When reserpine was added, the MIC value decreased 4-fold or more in 5 (27.8%) of the tetracycline-resistant isolates and in both nitrofurantoin-resistant isolates, while there was no change in the MIC value in chloramphenicol- and ampicillin-resistant isolates. The effect of reserpine on biocide MIC values is shown in Table 4. Reserpine reduced biocide tolerance to triclosan (46.7%) and povidone-iodine (32%) most effectively.
Antibiotic resistance and efflux pump genes determined by real-time PCR are given in Table 3. Antibiotic resistance genes tetK were found in 17 isolates, bla in 5 isolates, and mecA in 1 isolate, while tetL and cat genes were not found in any of the tested isolates. The efflux pump genes identified with high frequency were norE (80.8%), smr (88.5%), efrA (84.6%), efrB (80.8%), mdeA (84.6%), sepA (53.8%), and sugE (50%). The qacA/B gene was not detected in any of the tested isolates.
Discussion
Antibiotic and biocidal agents used for hygiene, treatment, and prophylaxis in the veterinary and food industries exert selective pressure, leading to the development and spread of antimicrobial-resistant bacteria in animals and humans (Endale et al., 2023). Many studies have shown that biocides and antibiotics have similar mechanisms of action on bacteria, and that the resistance mechanisms that develop in bacteria may also be the same. Therefore, biocides can promote the spread of antibiotic-resistant bacteria, and exposure to biocides at subinhibitory concentrations can lead to cross resistance with antibiotics (Sousa et al., 2025).
In this study, resistance to at least one antibiotic was identified in 26 of the 77 Staphylococcus isolates. Since the study aimed to investigate efflux pump proteins, a common antibiotic and biocide resistance mechanism in foodborne Staphylococcus spp., further analysis was performed on these 26 antibiotic-resistant isolates. The antibiotics with the most resistance were tetracycline (69.2%) and cefuroxime (38.5%), consistent with the literature (Ghabbour et al., 2022; Taddese et al., 2025). Of the 18 tetracycline-resistant isolates, 17 had the tetK gene, which encodes a type of efflux pump protein known to be associated with tetracycline resistance and from the MFS family (Roberts., 2005). None of the isolates had the tetL gene. Reserpine reduced tetracycline resistance by 4-fold or more in 5 (27.8%) of the tetracycline-resistant isolates. This suggests that tetracycline resistance in Staphylococcus spp. isolated in our study may be due to the presence of the tetK gene, although a small portion of this resistance is due to reserpine-sensitive efflux proteins.
The bla gene causes resistance in Staphylococcus spp. by producing the beta-lactamase enzyme and enzymatically breaking down antibiotics such as penicillin and cephalosporin. The mecA gene, on the other hand, causes antibiotic resistance by inhibiting the entry of antibiotics into the cell by enabling the production of alternative proteins to penicillin-binding proteins (PBPs), which are the receptors for beta-lactam antibiotics (Tahmasebi et al., 2017). In our study, the bla gene was positive in 5 (50%) of the isolates identified as resistant to cefuroxime, while the mecA gene was detected in only one isolate showing resistance to both cefuroxime and ampicillin. The mecA-positive isolate was negative for the bla gene. This suggests that the beta-lactam antibiotic resistance of this isolate may have developed due to a change in the PBP protein caused by the mecA gene, not in beta-lactamase production. Ghabbour et al. (2022) examined the prevalence of the mecA and blaZ genes in S. aureus strains isolated from retail foods in Egypt. The results showed a lower prevalence of mecA (34%) and a higher prevalence of blaZ (82%). They suggested that some phenotypic beta-lactam resistance may be associated with mecA, even in the absence of blaZ (Ghabbour et al., 2022). The results of this study are similar to and support our findings.
The efflux pump inhibitor reserpine reduced the MIC values of all cefuroxime-resistant bacteria 4- to 256-fold, providing strong evidence that cefuroxime resistance in Staphylococcus spp. isolated from animal food sources originated from the efflux pump system. Similarly, Costa et al. (2013) reported that administration of reserpine to multidrug-resistant S. aureus isolates resulted in 2- to 128-fold reductions in antibiotic MIC values, supporting the role of efflux pump activity in beta-lactam resistance. Similar studies support the hypothesis that cefuroxime resistance in our study is mainly due to efflux pump activity (Huet et al., 2008; Costa et al., 2013). The cat resistance gene, which encodes the chloramphenicol acetyltransferase enzyme (Schwarz et al., 2004), was not detected in any of the 3 chloramphenicol-resistant Staphylococcus spp. isolates in our study. In addition, the chloramphenicol MIC value in these isolates was not affected by reserpine. This indicates that the resistance may not be due to the reserpine-sensitive efflux pump.
Hughes and Ferguson (2017) used 198 clinical S. aureus isolates and the agar dilution method to show the MIC values for chlorhexidine were between 0.5 and 8 mg/L. The MIC values for triclosan were between <0.06 and 16 mg/L. Compared to our study (Table 4), the Staphylococcus isolates had slightly higher MIC values for chlorhexidine. However, our MIC values for triclosan were much higher: 512–64 μg/mL in seven isolates and 32–4 μg/mL in two isolates. These results suggest that biocidal use in the food and livestock sectors is somewhat uncontrolled.
Marquez et al. (2017) screened 80 gram-positive bacteria isolated from various cheese samples produced by small and medium-sized enterprises in Spain, including Bacillus, Enterococcus, and lactic acid bacteria. They found that benzalkonium chloride MIC values were 2.5–50 mg/L, triclosan MIC values were 2.5–7.5 mg/L, and hexachlorophene MIC values were 5–10 mg/L. Although the MIC values for benzalkonium chloride were similar to those in our study, the MIC values for triclosan and hexachlorophene were much higher in our study. Marquez et al. (2017) noted that these values indicate reduced biocide sensitivity. Therefore, the isolates in our study also have significantly reduced biocide sensitivity.
Jomha et al. (2014) examined biocidal resistance of 86 gram-positive bacteria isolated from a hospital in Lebanon. Serial dilutions based on commercially available biocide concentrations were performed. Of the 31 S. aureus isolates, 12.9% were resistant to quaternary ammonium compound-containing disinfectants, 20.1% were resistant to sodium hypochlorite, and none were resistant to povidone-iodine. Of the 31 coagulase-negative Staphylococcus, 6.5% were resistant to quaternary ammonium compound-containing disinfectants and sodium hypochlorite; no resistance to povidone-iodine was observed.
Fernandes-Fuentes et al. (2014) collected 11 Bacillus, 25 Enterococcus, and 10 Staphylococcus isolates from organic foods. The MIC values of various biocidal substances decreased 4-fold or more in four isolates for benzalkonium chloride, 14 isolates for hexachlorophene, 19 isolates for chlorhexidine, and two isolates for triclosan when 25 μg/mL of reserpine was added to the medium. In our study, reserpine most effectively reduced resistance to povidone-iodine and triclosan (Table 4). This indicates that resistance to other biocidal substances, whose MIC values decrease with the addition of reserpine, particularly povidone-iodine and triclosan, may have developed with the reserpine-sensitive efflux pump system.
norA, mdeA, mepA, sepA, smr, and qacA/B genes have been studied in clinical isolates of S. aureus, especially methicillin-resistant/susceptible S. aureus (MRSA/MSSA). The protein products of these genes are efflux pumps, which are often involved in the development of resistance to antimicrobial agents (Hassanzadeh et al., 2020). Other genes acrB, efrA, and efrB have also been identified in gram-negative bacteria (e.g., Escherichia coli and Klebsiella) and contribute to the removal of antimicrobials from the cell (Fuentes et al., 2014).
Hassanzadeh et al. (2017) found the mecA gene in all 60 clinical MRSA isolates they collected in Tehran. They also found the mdeA gene in 43 isolates, the mepA gene in 37, the norA gene in 22, the norB gene in one, the sepA gene in 19, and the smr gene in 11 (Hassanzadeh et al., 2017). The results for genes other than mecA are similar to those in our study (Table 3). Molecular analysis confirmed the presence of multiple efflux pump genes in the resistant isolates. The most prevalent were smr, efrA, efrB, mdeA, and norE; these were detected in over 80% of the tested strains. Similar distributions of these genes have been observed in hospital and foodborne Staphylococcus isolates worldwide (Costa et al., 2013; Hassanzadeh et al., 2020).
PCR/qRT-PCR analyses of S. aureus clinical isolates showed that the prevalence of chromosomal genes norA, norB, norC, mepA, sepA, and mdeA was over 90% (Suma et al., 2023). In comparison, plasmid-borne genes qacA/B (approximately 20%) and smr (8–30%) were less common. These gene clusters (e.g., norA + norB + norC + mepA + sepA + mdeA) have been associated with widespread microbial resistance in MRSA/MSSA strains characterized by ciprofloxacin, vancomycin, and mecA, leading to clinical treatment challenges (Suma et al., 2023). Our research results indicate that efflux pump genes are also frequently observed in animal-based foodborne bacteria, emphasizing that transferring these genes to human pathogens through gene transfer could lead to significant problems.
The presence of multiple efflux pump genes in isolates from food products could contribute to the spread of antimicrobial resistance throughout the food chain. However, limitations such as the lack of gene expression data and the relatively small sample size in our study should be considered. Future studies that include transcriptomic and efflux activity analyses are needed to further elucidate the contribution of these genes to resistance phenotypes.
Conclusion
This study showed the high prevalence of efflux pump-related antimicrobial resistance genes in Staphylococcus spp. isolated from milk and meat samples in Ankara, Türkiye. The detection of smr, efrA, efrB, mdeA, and norE genes in a majority of resistant isolates, along with the significant reduction in MIC values by reserpine, indicates that efflux-mediated mechanisms play a key role in both antibiotic and biocide resistance. The absence of the qacA/B gene, despite widespread resistance, suggests possible regional or ecological differences in gene dissemination. The frequent occurrence of the tetK and bla genes, and the identification of the mecA gene in one isolate, further reflect the complexity of resistance determinants in foodborne Staphylococcus spp.
The decreased susceptibility to commonly used biocides, especially triclosan and povidone-iodine, provides information about the excessive and unnecessary use of these substances in food-related environments and raises the issue that this situation may contribute to the selection of resistant strains. The findings of the study highlight the need for better regulation and more appropriate use of antimicrobial agents in the veterinary and food sectors.
Although this study provides significant insights into the presence of resistance genes, further research, including gene expression analysis, transcriptomic profiling, and efflux pump functional assays, is essential to clarify the contribution of these genes to phenotypic resistance. Surveillance of efflux pump gene prevalence in foodborne bacteria is critical for anticipating public health risks and guiding evidence-based antimicrobial stewardship strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abbas A Barkhouse A Hackenberger D Wright GD 2024 Antibiotic resistance: A key microbial survival mechanism that threatens public health Cell Host & Microbe 32 6 837 851 10.1016/j.chom.2024.05.015 38870900 · doi ↗ · pubmed ↗
- 2Aygül A 2015 Antibiyotik direncinde dışa atım sistemlerinin ve dirençle mücadelede dışa atım pompa inhibitörlerinin önemi Mikrobiyoloji Bülteni 49 2 278 291 (in Turkish). 10.5578/mb.8964 26167829 · doi ↗ · pubmed ↗
- 3Chajecka-Wierzchowska B Sierpińska A Nalepa W Zadernowska M Łaniewska TrokenheimŁ 2015 Coagulase-negative staphylococci (Co NS) isolated from ready-to-eat food of animal origin phenotypic and genotypic antibiotic resistance Food Microbiology 46 222 226 10.1016/j.fm.2014.08.001 25475289 · doi ↗ · pubmed ↗
- 4Clinical and Laboratory Standards Institute 2018 Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard 10th ed M 07 A 11 Clinical and Laboratory Standards Institute Wayne, PA
- 5Coombs K Rodriguez-Quijada C Clevenger JO Sauer-Budge AF 2023 Current understanding of potential linkages between biocide tolerance and antibiotic cross-resistance Microorganisms 11 8 2000 10.3390/microorganisms 11082000 PMC 1045925137630560 · doi ↗ · pubmed ↗
- 6Costa SS Viveiros M Amaral L Couto I 2013 Multidrug efflux pumps in Staphylococcus aureus: an update The Open Microbiology Journal 7 59 10.2174/1874285801307010059 23569469 PMC 3617543 · doi ↗ · pubmed ↗
- 7Endale H Mathewos M Abdeta D 2023 Potential causes of spread of antimicrobial resistance and preventive measures in one health perspective-a review Infection and Drug Resistance 16 7515 7545 10.2147/IDR.S 428837 38089962 PMC 10715026 · doi ↗ · pubmed ↗
- 8Fernandes-Fuentes MA Abriouel H Morente EO Pulido RP Gálvez A 2014 Genetic determinants of antimicrobial resistance in Gram positive bacteria from organic foods International Journal of Food Microbiology 172 49 56 10.1016/j.ijfoodmicro.2013.11.032 24361832 · doi ↗ · pubmed ↗