First Expert Elicitation of Knowledge Conducted in 2023 on Possible Drivers of Observed Increasing Epizootic Haemorrhagic Disease Incidence in Europe
Claude Saegerman

TL;DR
Experts identified key factors driving the rise of Epizootic Haemorrhagic Disease outbreaks in Europe, including climate effects on vectors and lack of vaccines.
Contribution
This is the first expert elicitation study in 2023 to systematically prioritize drivers of increasing EHD incidence in Europe.
Findings
Temperature effects on vector populations were identified as a top driver.
Unavailability of vaccines for Circulating Serotype 8 was highlighted as a critical factor.
Animal movements from third and neighboring countries were ranked as significant contributors.
Abstract
Epizootic haemorrhagic disease (EHD) is a viral noncontagious arthropod‐borne disease transmitted by blood‐sucking midges of the genus Culicoides. Its causative agent, the EHD virus (EHDV), belongs to the genus Orbivirus and is responsible for domestic and wildlife ruminants’ disease outbreaks, especially in North America, Asia, Africa and Oceania. These outbreaks not only cause significant morbidity and mortality but also have welfare, social and economic implications. Recently, EHD has become a real threat to the European Union, with outbreaks confirmed in October and November 2022 in Sardinia and Sicily. To better understand this phenomenon, we investigated the drivers of the observed increasing EHD incidence in Europe through expert knowledge elicitation. We listed 51 possible drivers grouped in eight domains and elicited 41 European experts to (i) allocate a score per driver, (ii)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
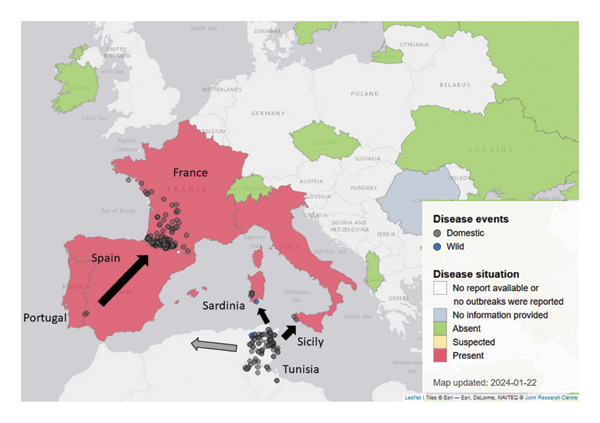 Figure 1
Figure 1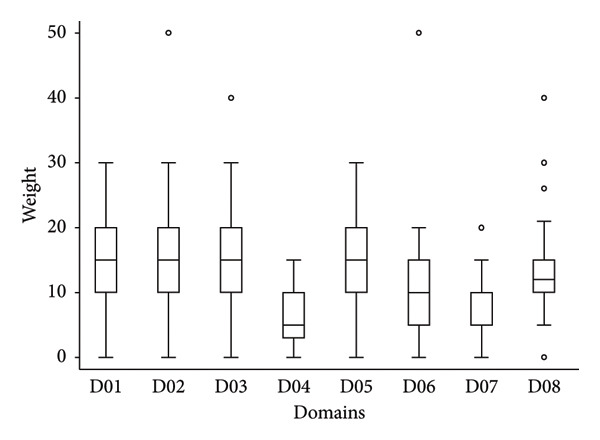 Figure 2
Figure 2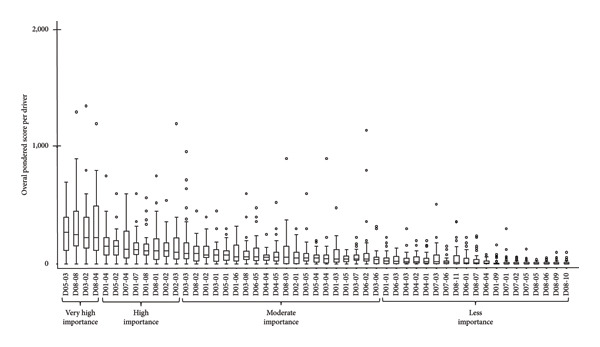 Figure 3
Figure 3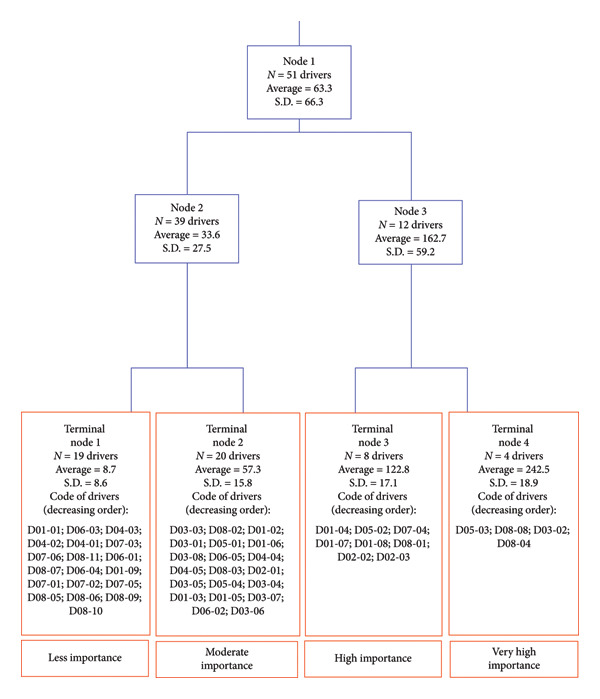 Figure 4
Figure 4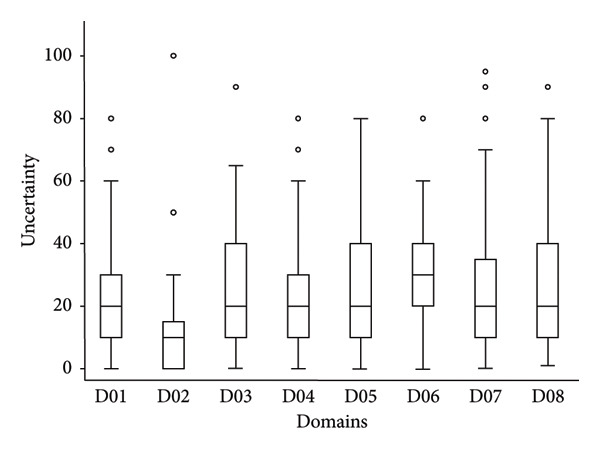 Figure 5
Figure 5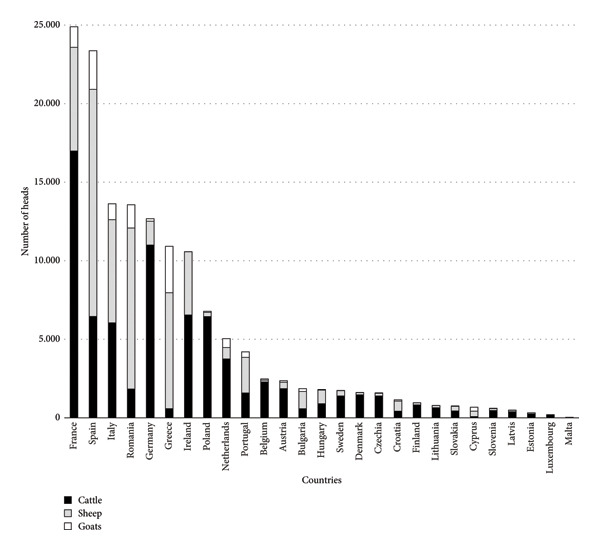 Figure 6
Figure 6| Country | First event | Last event | Domestic ruminants | Wild ruminants(a) | ||||
|---|---|---|---|---|---|---|---|---|
| Cattle | Sheep and goats | Domestic deer | Reed deer ( | Roe deer ( | Fallow deer ( | |||
| Spain | 15/11/22 | 15/11/23 | 251(b) | 0 | 20(d) | ?(b) | 1 | 0 |
| France | 04/09/23 | 17/11/23 | 3764(c) | 0 | 0 | 0 | 0 | 0 |
| Italy (Sardinia) | 28/10/22 | 19/10/23 | 8(e) | 1(e) | 0 | 1 | 0 | 1 |
| Italy (Sicily) | 25/10/22 | 09/12/22 | 2 | 0 | 0 | 0 | 0 | 0 |
| Portugal | 13/07/23 | 13/09/23 | 71 | 0 | 2 | 0 | 0 | 0 |
| Total | 25/10/22 | 07/11/23 | 4096 | 1 | 22 | 1 | 1 | 1 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-Borne Animal Diseases · Animal Disease Management and Epidemiology · Viral Infections and Vectors
1. Introduction
Epizootic haemorrhagic disease (EHD) is a noncontagious arthropod‐borne viral disease transmitted by specific species of blood‐sucking midges of the genus Culicoides [1]. Its causative agent, the EHD virus (EHDV), has 10 double‐stranded ribonucleic acid (dsRNA) segments (seven structural [VP1–VP7] and four nonstructural [NS1, NS2, NS3/NS3A, NS4] proteins), and belongs to the family Sedoreoviridae, genus Orbivirus [2]. There are at least seven serotypes of EHDV recognized so far, with putative serotypes recently described. Ibaraki virus is the prototype member of the EHDV serogroup (Serotype 2) [2]. EHDV is transmitted by specific species of biting midges belonging to the family Ceratopogonidae, genus Culicoides, which act as biological vectors. Culicoides biting midges are hematophagous insects approximately 2–4 mm in length, which are usually active around sunrise and sunset, with activity peaks in early evening. They can bite outdoors (mostly exophilic) but also indoors (e.g., species of the Obsoletus group) when animals are housed in barns [3–6]. Larvae of many livestock‐associated species develop in warm semi‐aquatic microhabitats, especially around facilities housing ruminants [7]. For instance, according to Ref. [8], larvae of species of the Obsoletus group have been collected from dung, waterlogged soil, compost and leaf litter, while those of the Pulicaris group have generally been found in waterlogged soil.
Culicoides dispersal is described as stratified, due to the combination of dispersal processes occurring actively at short distances, and passively or semi‐actively at long distances [9]. Recent population genetics studies have demonstrated a high dispersal capacity over lands [10]. Mark‐release‐recapture studies on Culicoides species showed that the postrelease dispersal distance travelled for two nights ranges from 1 to 2.5 km and is linked to the gradual search for hosts or oviposition sites [11]. The maximum recapture distance recorded was 6 km for Culicoides mohave in a particular desert landscape [12]. In 2017, Sanders et al. attempted to quantify Culicoides dispersal over land and demonstrated through a capture–mark–recapture study that 84.4% of flights of more than 1 km took place downwind, while only 15.6% of flights were made upwind [13]. In addition, the dispersal happened over distances of at least 3 km. Many studies have reported a correlation between disease movement and wind‐borne transport of Culicoides during outbreaks [9, 14–18]. The introduction of bluetongue virus (BTV) serotypes by wind‐borne infected Culicoides has been demonstrated from northern Africa to southern Europe [19], from Kenya to Southwest Indian Ocean islands [20], from Sardinia to the Balearic Islands [17], from Corsica to France [18], from the south of England to Ireland [21] and from the northern coast of France and Belgium to the United Kingdom [22]. Most of these studies were supported by modelling analyses in which dispersion trajectories were assessed using overwater atmospheric dispersion models [10]. Climatic parameters considered appropriate for the survival of midges carried on wind are temperatures of 15°C–35°C, relative humidity above 25% and wind velocity of up to 10 m/sec (35 km/h) [23–26].
European Culicoides were found to be biting a wide range of mammals [27]. However, many Culicoides species feed preferentially on horses and domestic and wild ruminants but have been found in pig farms, and blood meal analysis demonstrates that they can feed on pigs [28, 29]. Potential vectors of EHDV in northern Europe have been shown to bite susceptible species of deer in Spain [30].
Female Culicoides of competent species become infected with EHDV and can transmit the virus to susceptible ruminants after an extrinsic incubation period of approximately 10–14 days [31]. As in the case of BTV infection, viraemia can be prolonged in EHDV‐infected ruminants because of the virus association with ruminant erythrocytes [32–34].
The incubation period of EHD in animals is estimated to be 2–10 days. The clinical signs of EHD manifest as a haemorrhagic disease in some species of deer, but domestic ruminants may also be infected [35, 36]. EHD was first described in 1955, in white‐tailed deer (Odocoileus virginianus) of New Jersey (USA) [37]. This species and other cervid species are severely affected by the virus, often resulting in high levels of mortality associated with high fever, lethargy, oedema, ulcerations of the dental pad and oral mucosa, haemorrhages of the heart, lungs, major blood vessels and other tissues [35]. Frequently less severe or asymptomatic and sometime mild‐to‐severe (e.g., Refs. [38–40]) EHDV infections are observed in cattle, which are considered the reservoir host for the virus [31, 41]. The clinical signs and patterns of the disease and the mode of replication of EHDV are similar to that of BTV [42, 43]. Photographs of EHD affected animals are available in the Atlas of Transboundary Animal Diseases edited by the World Organization for Animal Health (WOAH), which is intended to assist Veterinary Service field staff involved in animal disease surveillance and diagnostics to identify important transboundary diseases of livestock [43].
Economic losses due to EHD have only been properly assessed in Israeli dairy herds during an EHDV‐7 outbreak in 2006 [42]. Firstly, a high correlation was observed between EHDV seroprevalence and milk loss (average milk loss for herds with seropositivity of 26%–50%, 51%–75% and 76%–100% was 84, 133 and 204 kg of milk/milking cow, respectively), indicating the importance of active surveillance to predict the losses [38]. Secondly, a 1.42% excess mortality was observed in herds with seroprevalence above 50%. Thirdly, an average loss of USD 26.5 per cow and total losses for the dairy cattle industry ranging from USD 1,591,000 to USD 3,391,000 were estimated. Due to its economic impact in livestock, EHD was introduced as a notifiable disease to the World Organization of Animal Health in May 2008, and it is notifiable under the EU Animal Health Law, Commission Implementing Regulation 2018/1882/EU.
In North America, Asia, Africa and Oceania, EHD regularly causes large regional disease outbreaks that affect domestic and wild ruminants, mainly some species of deer, antelope and cattle. These outbreaks cause severe morbidity and mortality but also have welfare, social and economic repercussions [2, 44]. At the end of September 2021, a new strain of EHDV‐8 was reported from cattle farms in central and western Tunisia. The virus subsequently spread rapidly to the northern and eastern regions in October and November, with more than 200 confirmed outbreaks recorded [36, 45, 46]. In autumn 2022, the same virus strain was found in symptomatic cattle in both Italy (Sardinia and Sicily) and Spain (Andalusia), posing a serious threat to the European livestock industry since no vaccine is currently available [47]. During 2023, the virus spread throughout Spain, reportedly reaching Portugal on 13 July 2023 and southwest France on 4 September 2023 [48, 49].
Wind‐borne dispersal of Culicoides is an important process governing both the arrival of BTV into new areas and its subsequent spread [50–52]. Winds were hypothesized to be the primary cause of the long and medium distance distribution of EHDV during the 2006 Israeli epidemic [53]. It is possible that the recent and simultaneous appearance of EHDV‐8 in multiple southern territories of the EU was caused by wind flows over seawater derived from Africa’s northern region [2] (Figure 1).
Geographical distribution of EHD confirmed outbreaks in the European Union and neighbouring countries, situation at January 2024 (source: https://animal-diseases.efsa.europa.eu/EHDV/#Geographicaldistribution). Legend: black arrows indicate the direction of spread of EHD. Grey arrow: EHDV‐8 could have circulated without notification in other western countries of the Maghreb [36, 45, 46].
The aim of this study was to investigate, for the first time, possible drivers of the observed spread of EHD in Europe, using expert elicitation of knowledge. A multicriteria decision analysis (MCDA) method was chosen because it allows the systematic integration of information from a range of sources [54] and aims to improve repeatability and transparency [55].
2. Materials and Methods
The process of this expert knowledge elicitation is identical to the previously published methodology utilized by the UREAR‐ULiege for other diseases that are emerging or zoonotic [56–59]. Briefly, we enumerated 51 possible drivers that were organized into eight different domains and each driver had a description that was specific to their definition (four or five modalities) (Appendix A1). Forty‐one European experts (Appendix A2) were asked to (i) rate the drivers’ capabilities, (ii) weight the rating within each domain [60] and (iii) calculate the degree to which each domain was capable of providing information and how this information was aggregated and then weighted (Appendix A3). A weighted average score per driver was determined. If all the drivers were considered equivalent by the experts, each of them would have received the same score. If not, the expert attributes more weight on the most important drivers contributing to the observed increasing EHD incidence in Europe. Finally, drivers with similar scores were clustered in several terminal nodes using a regression tree analysis (minimizing the standard error in each node). In addition, the degree of uncertainty was queried at the domain level for each expert. To identify whether the ranking of drivers of the observed increasing EHD incidence in Europe was influenced by the choice of experts or the country of origin of experts, a sensitivity analysis was performed.
The details of the methodology are described in Appendix A4 and comprising six sections: (i) the questionnaire design; (ii) the expert elicitation to assess the drivers of observed increasing cases of EHD in Europe; (iii) the scoring, weighting system and level of uncertainty; (iv) the calculation of an overall weighted score for each driver and ranking process; (v) the cluster analysis and (vi) the sensitivity analysis to test the robustness of the expert elicitation.
3. Results
3.1. Response Rate and Field of Expertise Mobilized by the Experts
Sixty‐eight professionals with scientific knowledge and/or field knowledge or experience regarding EHD and EHDV were contacted, i.e., respectively, 16, 20 and 32 from Belgium (boundaries shared with France that experienced EHD outbreaks), France (country currently with cases of EHD) and other European countries concerned by the future EHD spread. The fields and diversity of expertise are summarized in Appendix A2. Among all professionals contacted, 13, 12 and 16 responded and originated from Belgium, France or other European countries, respectively. The participation rate was 60.3%.
3.2. Estimating the Overall Weighted Score and Ranking of the Observed Increasing EHD Incidence in Europe
The medians of the weight between the eight domains of drivers as well as for the different 51 drivers were not equal according to the nonparametric Kruskal–Wallis test (chi‐squared test = 71.5 with 7 d.f. and α = 0.05, p value = 0.0001; and chi‐squared test = 744.2 with 50 d.f. and α = 0.05, p value = 0.0001, for the weights between domains and weights of the different drivers, respectively) (Figure 2). The median of the weights of domains D04 (p value < 0.001), D06 (p value = 0.034) and D07 (p value < 0.001) were significantly lower than the median of D01 as a reference (median regression).
Boxplot of the relative importance of the eight domains of possible drivers of the observed increasing cases of EHD in Europe (N = 41 European experts). Legend: The bold line represents the median of the score distribution between the different experts attributed to each domain; the solid lines at the bottom and top of each rectangle represent, respectively, the first and the third quartiles; adjacent lines to the whiskers represent the limits of the 95% confidence interval; small circles represent outside values. The eight domains of drivers are D01, disease/pathogen characteristics; D02, distance to Europe and the expert’s country (spatial–temporal scales); D03, ability to monitor, treat and control the disease; D04, characteristics of European farms; D05, global change; D06, interface with wildlife; D07, human activity; and D08, economic and trade activities.
Twelve drivers out of 51 were ranked in decreasing order as being of very high importance (N = 4) and of high importance (N = 8) in the probability of playing a key role in the observed increasing EHD incidence in Europe. The first most important driver was from the global change domain, the influence of the temperature on the abundance/survival of the vector populations (D05–03). The second and the fourth most important drivers were from the economic and trade activities domain, the legal or illegal movements of live animals from third countries (i.e., countries that are not a member of the European Union as well as countries or territories whose citizens do not enjoy the European Union right to free movement) (D08–08) and from neighbouring or European Union countries (D08–04), respectively. The third most important driver was vaccine availability (D03–02). The following eight drivers, with high importance in the probability of playing a key role, were the ease or speed of the pathogen spread (D01–04); the influence of humidity on the abundance or survival of the vector population (D05–02); live animal transport vehicles (D07–04); the existence of vectors and potential spread (D01–07), the mode of transmission of the pathogen, i.e., via vector (D01–08); the decrease in resources allocated to disease surveillance in animals and/or in the environment (D08–01); the European geographic proximity of the pathogen or disease to the expert’s country (D02–02); and recently reported cases in Europe (D02–03) (Figure 3).
Ranking of the overall weighted score for each potential driver of the observed increasing epizootic haemorrhagic disease incidence in Europe (boxplot based on input from 41 European experts). Legend: X axis represents the drivers with the following codification: D01–D08 refer to the eight domains of drivers and D01–01 to D08–11 refer to a specific driver (for the codification, see Appendix A1). Drivers were grouped as having ‘very high importance’, ‘high importance’, ‘moderate importance’ and ‘less importance’ according to the cluster analysis of their weighted scores.
3.3. Cluster Analysis
Results of the regression tree analysis revealed four significantly different clusters of drivers (Figure 4) according to the nonparametric Kruskal–Wallis test (chi‐squared test = 44.2 with three degrees of freedom (d.f.) and α = 0.05; p value = 0.0001). Four drivers were assigned the category of ‘very high importance’, eight drivers received the category of ‘high importance’, twenty drivers received the category of ‘moderate importance’ and 19 drivers received the category of ‘less importance’.
Aggregation of drivers of the observed increasing epizootic haemorrhagic disease incidence in Europe into four homogenous groups using a regression tree analysis. Legend: N = Number, S.D. = Standard deviation.
3.4. Sensitivity Analysis of the Impact of Experts on the Final Ranking of the Observed Increasing EHD Incidence in Europe
Firstly, the results of two different sensitivity analyses indicated that, regardless of the expert excluded, removing some experts only had very limited and no significant effects on the ranking compared to the reference (all experts elicited). Secondly, using 10 bootstraps of 30 experts among 41, the Pearson correlation coefficient between bootstraps against the ranking of 41 experts as a reference was very high (value between 0.974 and 0.994, with a p value of < 0.0001). Thirdly, when comparing the ranking of drivers between either Belgian experts (N = 13), French experts (N = 12) or other experts (N = 16) and all experts as a reference, the Pearson coefficient of correlation was of 0.952, 0.974 and 0.970, respectively, with a p value of < 0.0001.
3.5. Level of Uncertainty per Domain of Drivers
The level of uncertainty per domain of drivers is shown in Figure 5. The medians of the uncertainty between domains of drivers were not equal according to the nonparametric Kruskal–Wallis test (chi‐squared test = 32.5 with 7 d.f. and α = 0.05, p value = 0.0001). Domain D02 (distance to Europe and the expert’s country) had a median uncertainty of 10 and Domain D06 (interface with wildlife) had a median uncertainty of 30, respectively, significantly higher and lower than the median of the other domains, which was equal to 20 (median regression; p value = 0.05). In addition, there is no linear (Pearson coefficient correlation = −0.09; p value = 0.11) or nonparametric (Spearman rank correlation = −0.08; p value = 0.13) relationship between the median weight and the median uncertainty per domain.
Level of uncertainty per domain of drivers. Legend: The bold line represents the median of the level of uncertainty attributed by experts using a scale from 0 (minimal uncertainty in the scoring) to 100 (maximum uncertainty in the scoring); the solid lines at the top and bottom of each rectangle represent, respectively, the first and the third quartiles; adjacent lines to the whiskers represent the limits of the 95% confidence interval; and small circles represent outside values. The eight domains of drivers are D01, disease/pathogen characteristics; D02, distance to Europe and the expert’s country (spatial–temporal scales); D03, ability to monitor, treat and control the disease; D04, characteristics of the European farms; D05, global change; D06, interface with wildlife; D07, human activity; and D08, economic and trade activities.
4. Discussion
Fifty‐one possible drivers of observed increasing EHD incidence in Europe were ranked and aggregated into four homogenous groups according to expert elicitation of knowledge. Only the first 12 most important ranked drivers will be further discussed with a focus on those categorized in the regression tree as ‘very high importance’ (N = 4) and ‘high importance’ (N = 8), respectively. Moreover, the sensitivity analysis showed a limited and insignificant effect of experts involved (using both bootstraps and country‐origin groups of experts), indicating an acceptable robustness of the expert elicitation for drivers included in the first two terminal nodes of the tree. In addition, the median‐level of uncertainty for all domains of drivers was moderate (around 20 on a scale from 0 up to 100) except for Domain D02 (distance to Europe and the country of expert—spatial–temporal scales), which was less (around 10), and Domain D06 (wildlife interface), which was higher (around 30). Regarding the last domain with the highest median uncertainly, more surveillance and research‐action are recommended on European wildlife to assess more properly the susceptibility of deer species regarding to EHDV‐8 and its possible role as the reservoir of the disease [2, 34, 61].
The most important driver for the observed increasing cases of EHD in Europe recognized during this expert elicitation was the influence of the temperature on the abundance or survival of vector populations (D05–03). In Sardinia, EHDV‐8 was detected in whole bodies of C. imicola, C. obsoletus/C. scoticus, C. newsteadi, C. pulicaris ss and C. bysta. Note that simple detection of RNA of EDHV is not the proof of vector competency and capacity. However, since the first four species are also able to transmit BTV, and given the current distribution of BTV in Europe, further spread of EHDV‐8 is expected in Europe [62]. According to a recent modelling study, the habitat suitability of C. imicola will likely expand to higher latitudes due to three main variables identified in decreasing order of importance, i.e., the temperature seasonality, the precipitation of the coldest quarter and the average temperature of the wettest quarter with a 30.3%, 29.5% and 16.5% contribution, respectively [63]. This modelling confirms earlier studies (e.g., Refs. [52, 64]). Indeed, the current suitable habitats for the Afro‐Asian biting midge vector C. imicola are distributed in most of the southern part areas of America, southwestern Europe, most of Africa, the coastal areas of the Middle East, almost all regions of South Asia, southern China, a few countries in Southeast Asia and the whole of Australia [63]. Currently, we do not know that EHDV‐8 can be vectored by Palaearctic species of Culicoides but expects northwards. For example, in France, EHDV‐8 has already spread in areas where C. imicola is absent.
Under experimental conditions, the temperature also has a significant impact on the spring emergence of two Palaearctic species of Culicoides, C. chiopterus and C. dewulfi [65]. In this experiment, the emergence of Culicoides started when the temperature exceeded 20°C for several days [65]. In contrast, Losson et al. [66] recorded a few Culicoides captured during the 2006–2007 winter in Belgian cattle shed, at minimum temperatures of between 6°C and 12°C. Subsequent studies have confirmed the trapping of Palaearctic species of Culicoides at similar or even slightly lower minimum temperatures [67, 68]. Another study demonstrated activity down to 4°C [69].
The temperature has a crucial effect on the extrinsic incubation period, i.e., the time between uptake of virus by the insect vector and its presence in the salivary gland. However, due to the nature of the required experimental work, temperature thresholds have not been systematically investigated for EHDV at this time [70]. Temperature also influences biting rates [71]. Temperature is undoubtedly the main environmental factor influencing the behaviour and survival of these midges; their activity peaks between 13°C and 35°C although these limits vary according to species [72]. In relation to BTV, Culicoides kept at 15°C require several weeks to complete the extrinsic incubation period, while those kept at 30°C may complete incubation in a couple of days [73]. Higher temperatures can reduce the time needed for a vector to digest a blood meal, increasing the frequency of blood‐feeding [8]. All these events increase the opportunities for transmission [8].
In Europe, since June 2023, every month has been the warmest on record for the respective month of the year [74]. There is likely to be an effect of such climate pressure on the abundance, activity and survival of Culicoides and their vector competence. The distribution (map) of different Culicoides species in Europe is frequently updated on the dedicated website of the European Centre for Disease Prevention and Control (ECDC) and European Food Safety Authority (EFSA) at the following address https://www.ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/biting-midge-maps [75]. In the European context, reported EHD outbreaks are more numerous in domestic than in wildlife ruminants (Table 1). This is perhaps because wild ruminant EHD outbreaks are underreported, not because of a lower incidence or impact. In fact, in Europe, mandatory EHD surveillance is based on clinical signs. As mortality may be low at the individual level (one or a few cases by herd) and as wildlife surveillance is more often based on cadaver observation, a surveillance system for wildlife based on clinical signs would likely result in unobserved cases of EHD. In addition, the size of domestic and wild ruminant populations and their connectivity are of prime importance to understanding the current spread of EHD (Figure 6). To better assess the role of wildlife as the possible reservoir of EHDV‐8, we recommend more active surveillance of wildlife.
Ruminant population by EU Member States, expressed in thousand heads (animals) (source: Ref. [77]).
The second and the fourth most important drivers were related to the (il‐)legal movements of live animals from third countries (D08–08) or neighbouring countries of Europe and the European Union (D08–04). In European countries, there are currently fewer movements of live domestic animals originating from third countries than from neighbouring countries or European Union Member States (MS) (see also international trade statistics, available at the following URL address: https://www.trademap.org/tradestat/Index.aspx; accessed on 29 December 2023). Nevertheless, a proper estimation of the relative importance of illegal movements of live animals and their introduction pathways is deemed essential to set up risk‐based awareness, prevention and surveillance programs that correspond to reality [57]. A previous EFSA assessment of the risk of introduction of EHDV into the European Union was developed by considering three possible entrance pathways: (i) via imported infectious animals and (ii) via infectious vectors and (iii) other routes, e.g., via vaccines or germplasm. When quarantine and testing for EHDV are in place, the probability of importing an infectious animal into an EU MS is considered negligible. However, the probability of introducing an infectious animal through illegal livestock movements, illegal wild animal movements or wildlife transit could be high depending on the origin (EHDV circulating area) and the season of movement (vector activity). Indeed, following the introduction of EHDV by an infectious animal, the risk of exposure of EU susceptible animals during periods of vector activity was considered high. Depending on the temperature at the time of introduction (see also the main driver D05–03), the risk of spread was considered moderate or high [78].
The third most important driver was related to the vaccine availability for animals (D03–02). The only commercially available and currently used vaccines are manufactured for EHDV‐2 in Japan and are either live or inactivated vaccines [79]. In addition, there are currently no licenced vaccines available for EHD in the United States (for white‐tailed deer) other than autogenous vaccines [80], but some autogenous vaccines do not result in a high titre for homologous virus serotypes [81]. Currently, in Europe and Tunisia, the same EHDV Serotype 8 circulates with very high nucleotide sequence identity [46, 47] and no specific vaccine exists [44]. Some isolates of EHDV‐6 and EHDV‐8 previously showed some cross‐reactivity [44]. To limit EHDV infections, stop the virus from spreading and stop direct and indirect economic losses, more research is required on the development of vaccines [47, 80]. To encourage vaccine development, an assessment of the ratio of vaccine costs to EHD expenses may be beneficial (e.g., for a basic methodology [82]). If a vaccine is produced, its use could be regulated under the temporary authorization of animal vaccinations that have been previously discussed [83]. Several conditions must be fulfilled before a temporary authorization can be granted, e.g., ‘inactivated vaccines should be used preferentially in order to exclude reversion to virulence and re‐assortment between vaccine viruses and/or field strains of EHDV (and possibly a Differentiating Infected from Vaccinated Animals (DIVA) strategy would be great); decision‐making must be supported by scientific evidence and risk analysis; there must be a complete census of the susceptible animals that were vaccinated; vaccination protocols must be adhered to and there must be a scheme allowing for registration, delivery and follow‐up of vaccination, and monitoring, analysis and, possibly, adjustment of field use of the vaccination’ [83]. This temporary authorization must be replaced by a full authorization as quickly as possible. However, if the circulation of multiple serotypes could occur in the same area, farmers may be reluctant to vaccination against EHDV.
Three drivers were connected, i.e., the fifth (the ease or speed of spread of the pathogen; D01–04), the eighth (the existence of the vectors and potential spread; D01–07) and the ninth (the vector‐borne mode of transmission of the pathogen; D01–08) most important, according to the weighted scores. The basic reproduction ratio (R 0) is currently not known for EHD under European conditions but was estimated as moderate [84]. However, EHD is a ‘bluetongue‐like’ disease of ruminants caused by EHDV, which is also a Culicoides‐transmitted Orbivirus very closely related to BTV [85]. Using the surrogate data of the past BTV‐8 epizootic [86], the median R 0 of EHD can be provisionally estimated to be around 2. The speed of EHDV spread is currently undetermined and depends on the environmental heterogeneity and the presence of susceptible species (Figure 6), but the velocity has been estimated at ranging from 7.9 km/week (BTV‐1 in 2014 in Italy) to 24.4 km/week (BTV‐1 in 2008 in France) [87], up to 39 km/week (BTV‐8 in 2007–2008 in France) [88]. In addition, despite the restriction on animal movements, the effect of vaccination with inactivated vaccine on BTV speed was estimated (BTV‐1 in 2008 in southwestern France) at 3.2 km/day, i.e., a decrease from 5.4 km/day in nonvaccinating municipalities to 1.7 km/day in municipalities with vaccination [89]. The current observed spatiotemporal spread of EHDV‐8 in 2023 in France illustrates well the previous considerations and the competency and capacity of European Culicoides species.
The sixth most important driver was the influence of humidity on the abundance or survival of the vector population (D05–02). A > 25% relative humidity is a climatic parameter considered appropriate for the survival of midges [23–26]. A recent modelling study has indicated that the precipitation in the coldest quarter is an important predictor of the habitat suitability for C. imicola [63]. Indeed, the larval development of most Culicoides species is optimal in semi‐aquatic microhabitats, consisting mainly of warm, moist or wet substrates, rich in organic matter [90, 91]. In general, the larvae are mainly found in the first 5‐6 cm of the upper layer of the environmental substrate [92]. The pupae are found on the surface of the substrate (mud or water) where larval development takes place [91]. In terms of abiotic factors, temperature, humidity and rainfall are of importance for the vector biology, but temperature seems to be the most important parameter that was illustrated by this expert elicitation and supported by the scientific literature under field and experimental conditions (e.g., Refs. [93, 94]).
The seventh most important driver was live animal transport vehicles (D07–04). As an example, the intra‐EU trade of live animals in 2019, for several reasons, such as slaughter, fattening or breeding, represented around 4.18 million cattle and 3.36 million sheep and goats. For the same year, the extra‐EU trade of life animals represented 3.04 million cattle and 0.98 million sheep, mostly for exportation. Animals are transported primarily by sea and by road [95]. Whatever the direction (export or import), the risk of EHDV infection exists if Culicoides spp. are present in the live animal transport vehicle. Such a risk has not been estimated for EHDV so far. However, a previous risk assessment model has been built to assess the probability of a BTV‐8 outbreak following the introduction of Culicoides via live animal transport vehicles. The mean weighted annual risk for an outbreak caused by the transportation of a single vector from an affected northern European country varied from 1.8 × 10^−7^ to 3.0 × 10^−13^, depending on the country of origin [96]. Indeed, this mechanism represents a significant risk to BTV‐free countries, if a large number of vectors are transported. To date, there is no conclusive evidence that the use of insecticides or repellents singularly reduces the transmission of BTV in the field [97]. In specific scenarios, however, they have been shown to either kill biting midges or reduce host/vector contacts, and hence, they are used as mitigation when vaccines are not available [97], which is the case for EHDV‐8 in Europe at this time. Stabling is also effective in reducing host/vector contact where a high level of containment can be attained [97], but its feasibility under field conditions is of low. Insecticide‐treated meshes settled over stables windows in stables were found to kill biting midges quickly enough to prevent entry; field trials demonstrated substantial reductions in populations found in stables [98, 99]. As these studies mostly focused on horses, the logistics and reduced coverage existing in ruminant facilities may lessen such effects [97]. However, these measures could be applicable to the protection of animals from Culicoides during transport [99]. In addition, the chemical compounds derived from the neem tree (Azadirachta indica A. Juss; Meliaceae) have shown antilanding and antifeeding effects on adults of a colony of C. nubeculosus and field‐collected C. impunctatus [100]. Neem cake is a cheap and ecofriendly by‐product obtained from the extraction of neem oil and was used in a field case study (randomized complete block design with four replications of the treated and untreated plots) in Sardinia, Italy [101]. In this study, a treatment with neem cake at a dose of 100 g/m^2^ was applied on a larval breeding site of Culicoides located on the edge of a pond in a livestock farm. The emergence of Culicoides adults in treated and untreated plots was observed weekly using emergence traps before and after treatment. In neem cake‐treated plots, a significant reduction in the emergence of Culicoides was recorded up to 28 days. C. imicola represented about 10% of all emerged Culicoides adults and were highly sensitive to the neem cake [102]. Further studies are needed to gain more lines of evidence and design appropriate standard operational procedures to secure animal transportation. Furthermore, it is unclear what potential risk of introduction from EHDV‐infected midges exists during transport by airports, ships, trains or ports from countries that are not EHDV‐free. The closeness of the first outbreaks of BTV‐8 in 2006 and BTV‐3 in 2023 to international airports in Belgium and the Netherlands is striking. This raises the question of whether animal‐free transport of fresh products such as fruit or flowers from EHDV‐positive third countries generates a certain risk of EHDV strains entering Central Europe.
The 10th most important driver was the decrease in resources allocated to disease surveillance in animals and/or in the environment (D08–01). The resource attribution at EU level is related to the first pillar of the Animal Health Strategy with the objectives to (i) progressively eliminate animal diseases and to implement disease monitoring measures in the MS and the EU as a whole; (ii) ensure a high level of animal health, public health and consumer protection; and (iii) guarantee a high level of protection of both animal health and public health, to encourage the improvement of the productivity of the livestock sector and to contribute to the economic sustainability of the sectors directly or indirectly affected by an animal disease outbreak [103]. This is a challenging equilibrium especially in the context of recent major events (COVID‐19 and a resurgence of conflict and violence) [104, 105]. For BTV, a decrease of resources allocated to the disease surveillance in animals was observed from 112 million euros in 2009 to 3.8 million euros in 2014 [103]. Sufficient allocation is needed, especially during this winter to ensure a representative picture of the presence of EHDV occurrence in vector and domestic and wildlife compartments using both clinical and active surveillance (serological and molecular testing as the clinical expression of the disease can be mild or asymptomatic) when considering the current spread.
The 11th and the 12th most important drivers were the European geographic proximity of the pathogen/disease to the expert’s country (D02–02) and the recently reported cases in Europe (D02–03). It is important to be aware of the EHD situation and circulating serotype(s) of EHDV close to one’s country and to prepare some mitigation measures in the at‐risk countries. EHD is categorized as D and E under the EU Animal Health Law (Regulation [EU] 2016/429), which impose an obligation for livestock farmers to monitor for the disease and notify outbreaks, as well as restrictions on movements in affected MS. The EHD situation is registered in the EU Animal Diseases Information System (ADIS: https://food.ec.europa.eu/animals/animal-diseases/animal-disease-information-system-adis_en). In addition, the current incursion of EHD in southern Europe demonstrates the need to further promote a ‘global health’ approach, i.e., a long‐term collaboration between south and north countries to anticipate emerging vector‐borne diseases in the context of climate change. In this context, investment in capacity building (e.g., twinning programs for laboratories) and surveillance systems in the south can help northern countries to better anticipate any new event.
Expert knowledge elicitation may be limited by factors such as cognitive bias, overconfidence or replies that are influenced by recent, well‐publicized research (for a review, see Ref. [103]). The standardization of this expert elicitation (e.g., number of experts, choice of experts, different disciplines involved, protocol followed) and its sensitivity analysis takes partially these limitations into consideration in this elicitation to some extent. Furthermore, expert knowledge elicitation is an addition to empirical research, not a replacement for it [59]. Expert elicitation can be quickly implemented and empirical research that follows transparent and well‐designed protocol needs time. Both are necessary and not in opposition but should be a continuum with iterative process. In addition, as reviewed by prominent scientific journals, the elicitation of scientific and technical judgements from experts, in the form of subjective probability distributions, can be a valuable addition to other forms of evidence in support of public policy decision‐making [106].
The findings are specific to the European context (the purpose of this paper), but well the methodology can be easily translated to other regions and diseases (e.g., Refs. [56–59]).
5. Conclusions
Scientific knowledge on possible drivers of observed increasing animal cases of EHD in Europe was scanned. In this context, expert elicitation of knowledge and MCDA, in addition with clustering and sensitivity analyses together, allowed the identification of 12 drivers of either very high (N = 4) or high importance (N = 8), on which to concentrate future studies in order to increase and refine the understanding of the epidemiology of EHD under specific conditions of Europe and to support decision‐making to reduce exposure to EHDV and its impacts.
This expert elicitation of knowledge identified three main strategic axes of research or action. The first axis is the need for more research development to find a safe and protective vaccine towards EHDV‐8, possibly within the framework of a temporary authorization to vaccinate animals in emergency situations. The second axis is the promotion and operationalization of the ‘global health’ to anticipate the emergence of new pathogens or vectors in Europe, such as zoonotic arboviruses, which are already circulating in southern regions. This may be facilitated by the import of both new vectors and existing vectors attaining new competency capabilities to these new pathogens, in particular, due to climate change (e.g., Ref. [107]). The third axis includes awareness and information campaigns targeting both farmers and practitioners, strategies for the detection of infected animals (including subclinical presentations), systematic epidemiological investigations including vector surveillance and both passive and active surveillance of both susceptible ruminants and wildlife animal populations (especially at the front of EHD spread), restriction of movements and efforts to reduce contact between hosts and vectors [2, 108, 109].
Conflicts of Interest
The author declares no conflicts of interest.
Author Contributions
Conceptualization, methodology, software, validation, formal analysis, investigation, resources, data curation, writing–original draft preparation and writing–review and editing: Claude Saegerman; Experts included in the expert elicitation of knowledge are listed in Appendix A2.
Funding
No funding was received for this manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hoff G. L. and Trainer D. O. , Bluetongue and Epizootic Hemorrhagic Disease Viruses: Their Relationship to Wildlife Species, Advances in Veterinary Science & Comparative Medicine. (1978) 22, 111–132.216247 · pubmed ↗
- 2Jiménez-Cabello L. , Utrilla-Trigo S. , Lorenzo G. , Ortego J. , and Calvo-Pinilla E. , Epizootic Hemorrhagic Disease Virus: Current Knowledge and Emerging Perspectives, Microorganisms. (2023) 11, no. 5, 10.3390/microorganisms 11051339.PMC 1022437937317313 · doi ↗ · pubmed ↗
- 3Rozendaal J. A. , Chapter 1: Mosquitoes and Other Biting Diptera, Vector Control: Methods for Use by Individuals and Communities, 1997, World Health Organization (WHO), Geneva, 6–177.
- 4Zimmer J. Y. , Haubruge E. , Francis F. et al., Distribution of Potential Bluetongue Vectors on Belgium Farms, The Veterinary Record. (2008) 162, no. 21, 10.1136/vr.162.21.700, 2-s 2.0-44949238037.18503076 · doi ↗ · pubmed ↗
- 5Viennet E. , Garros C. , Rakotoarivony I. et al., Host-Seeking Activity of Bluetongue Virus Vectors: Endo/Exophagy and Circadian Rhythm of Culicoides in Western Europe, P Lo S One. (2012) 7, no. 10, 10.1371/journal.pone.0048120, 2-s 2.0-84868101509.PMC 348322123144735 · doi ↗ · pubmed ↗
- 6Barceló C. , del Río R. , and Miranda M. A. , Seasonal and Nocturnal Activity of Culicoides Spp. (Diptera: Cerato-Pogonidae) Adapted to Different Environments in the Balearic Islands, Diversity. (2023) 15, no. 5, 10.3390/d 15050690. · doi ↗
- 7Zimmer J. Y. , Losson B. , Saegerman C. , Haubruge E. , and Francis F. , Breeding Sites and Species Association of the Main Bluetongue and Schmallenberg Virus Vectors, the Culicoides Species (Diptera: Ceratopogonidae) in Northern Europe, Annales de la Société Entomologique de France. (2013) 49, no. 3, 335–344, 10.1080/00379271.2013.854100, 2-s 2.0-84890726726. · doi ↗
- 8Wilson A. J. and Mellor P. S. , Bluetongue in Europe: Past, Present and Future, Philosophical Transactions of the Royal Society B. (2009) 364, no. 1530, 2669–2681, 10.1098/rstb.2009.0091, 2-s 2.0-70349303746.PMC 286508919687037 · doi ↗ · pubmed ↗