Metarhizium pingshaense infection reverses insecticide resistance in Anopheles gambiae sensu lato by altering energy reserves and gene expression
Doubé Lucien Lamy, Francesco Baldini, Mafalda Viana, Iván Casas Gómez-Uribarri, Duangkamon Loesbanluechai, Erin S. Johnston, Meshach Lee, Najat Feruzi Kahamba, Edounou Jacques Gnambani, Issiaka Saré, Souro Abel Millogo, Moussa Namountougou, Abdoulaye Diabaté, Etienne Bilgo

TL;DR
A fungus called Metarhizium pingshaense can reverse insecticide resistance in malaria-carrying mosquitoes by depleting their energy and affecting gene activity.
Contribution
This study reveals how Metarhizium infection reverses insecticide resistance by altering energy reserves and gene expression in mosquitoes.
Findings
Metarhizium infection reduced survival of insecticide-resistant mosquitoes by depleting energy reserves.
Fungal infection suppressed detoxification gene expression, including GSTe2 and CYP6Z1.
Resistant mosquitoes became more susceptible to insecticides after fungal exposure.
Abstract
Entomopathogenic fungi like Metarhizium are emerging as effective biopesticides against malaria vectors. They reduce mosquito survival, fecundity, and flight ability, and reverse insecticide susceptibility in resistant Anopheles gambiae sensu lato strains. To elucidate the unclear underlying mechanisms, this study investigates the effects of fungal infections and insecticide exposure on the mosquito’s energy reserves and the expression of key metabolic and immune genes. Three mosquito types: (i) pyrethroid-resistant An. gambiae sensu lato and two laboratory colonies: (ii) pyrethroid-resistant An. coluzzii VKPER and (iii) insecticide-susceptible An. gambiae sensu stricto Kisumu were used. They were infected with Metarhizium pingshaense S10 strain at a concentration of 10⁷ spores/mL (treatment groups) and with solvent only (0.05% Tween® 80; control groups). Live mosquitoes were collected…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
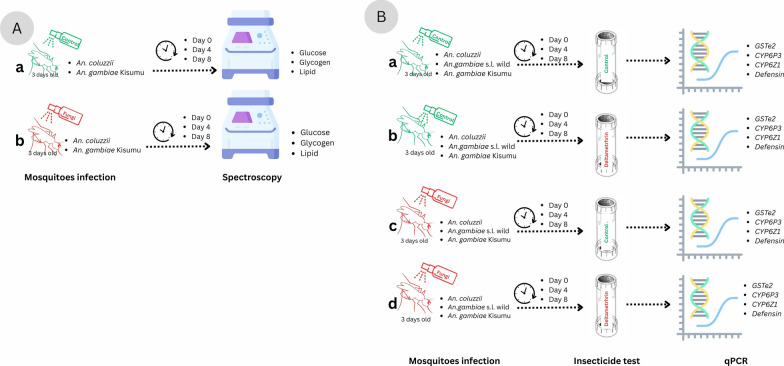 Figure 1
Figure 1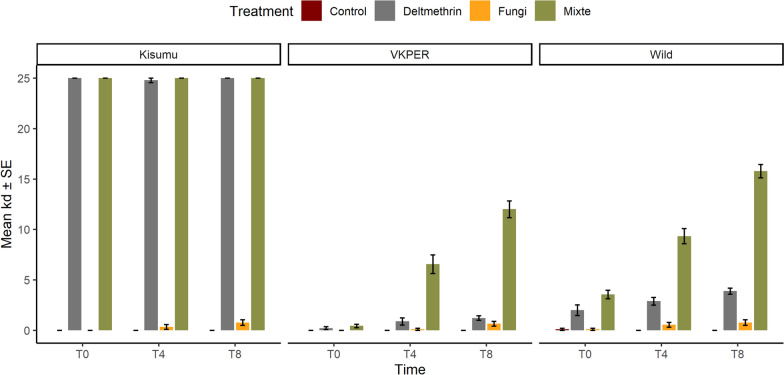 Figure 2
Figure 2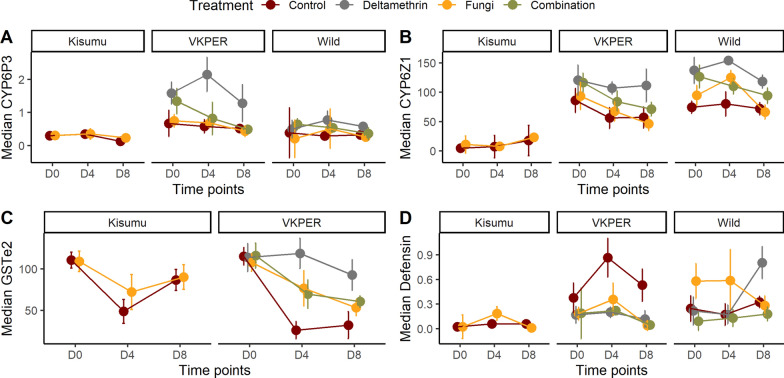 Figure 3
Figure 3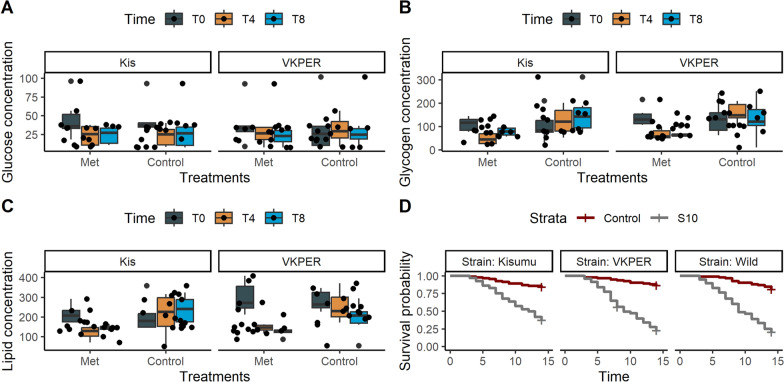 Figure 4
Figure 4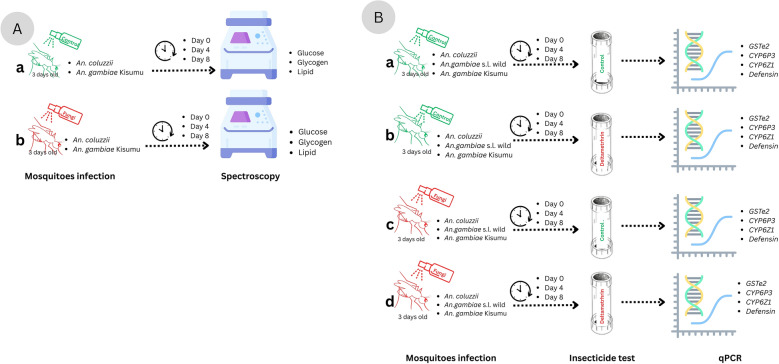 Figure 5
Figure 5- —https://doi.org/10.13039/100010269Wellcome Trust
- —Ministry of Higher Education Research and Innovation of Burkina Faso
- —Academy Medical Sciences Springboard Award
- —European Research Council, European Union’s Horizon 2020 Research and Innovation Programme
- —National Institute for Health Research (NIHR), using the UK’s Official Development Assistance (ODA) Funding
- —African Research Initiative for Scientific Excellence (ARISE)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Insect symbiosis and bacterial influences · Insect Resistance and Genetics
Background
In the early twenty-first century, malaria gained recognition worldwide as a priority global health issue, prompting intensive control efforts. These efforts have led to substantial progress in reducing Plasmodium falciparum infections in humans, with an estimated 50% decline in Africa and a 40% reduction in clinical malaria incidence between 2000 and 2015 [1, 2]. Vector control measures, which remain a pivotal public health strategy for malaria control, have been credited for this success [3], particularly two insecticidal interventions: long-lasting insecticide-treated nets (LLINs), which are bed nets treated with synthetic pyrethroid, and indoor residual spraying (IRS) [1, 3]. However, the intensive use of these tools has led to insecticide resistance to pyrethroids, the only class of insecticides approved for use in bednets, posing a significant threat to the progress of malaria control [4–6]. In addition to insecticide resistance, other factors such as limited access to healthcare, changing environmental conditions, and parasite resistance to antimalarial drugs also contribute to the persistent burden of malaria. As a result, the disease continues to cause hundreds of millions of cases and hundreds of thousands of deaths each year [7]. According to the latest World Malaria Report [8], malaria caused 246 million cases and claimed 569,000 lives in Africa in 2023 [9]. This underscores the need to develop innovative tools for sustainable malaria control, not only as alternatives or complements to existing strategies [6, 10], but also to understand the mechanisms of action for better integration.
Two main insecticide resistance mechanisms have been described in African malaria vectors: (i) target-site resistance, which prevents insecticide binding by altering insecticide target sites [11–14], and (ii) metabolic resistance, characterized by the overexpression of enzyme systems that enhance the detoxification or sequestration of insecticides [15–17]. This latter mechanism involves the action of cytochrome P450-dependent monooxygenases (CYPs), carboxylesterase, and glutathione S-transferases (GSTs) [18, 19]. Insecticide synergists, which are non-toxic chemicals added to insecticides to increase their lethality, or more generally, their effectiveness, have been proposed as a management strategy for metabolic insecticide resistance [20]. For example, piperonyl butoxide (PBO) is currently used in LLINs to improve the efficacy of pyrethroid-treated bed nets by inhibiting mosquito detoxification enzymes, ultimately increasing insecticide effectiveness [21, 22]. In malaria-endemic regions with insecticide resistance, PBO-treated nets can reduce malaria cases by more than 33% compared with conventional LLINs, making them a key control strategy [23–25]. However, PBO-LLINs may not have a significant effect in areas where metabolic resistance is not prevalent [26] or if mosquitoes develop high levels of P450 enzyme activity [23, 27].
Biopesticides such as the fungus Metarhizium offer a promising alternative to synthetic chemical insecticides for the control of malaria vectors. These agents are safer for the environment and can help mitigate the growing issue of insecticide resistance among mosquitoes [28–30]. Unlike synthetic chemical insecticides, biopesticides are generally host-specific and safer for non-target organisms, thereby reducing environmental impact [31]. Metarhizium infection has been shown to affect mosquito fitness [32, 33]. For example, Metarhizium anisopliae can disrupt the mosquito life cycle by shortening its lifespan and reducing fecundity [34], ultimately affecting population dynamics. Previous studies have also shown that Metarhizium can complement pyrethroids and potentially mitigate their resistance in mosquito populations [35–38]. Recently, we investigated the effects of combining these fungi and deltamethrin on insecticide-resistant mosquitoes and found that exposing Metarhizium-infected mosquitoes to deltamethrin significantly increased mortality compared to that in the non-infected group, suggesting that fungal infection can reverse insecticide resistance [35]. However, the mechanism has not yet been elucidated.
Mosquito fitness is closely related to energy levels and immunity. Mosquitoes emerge with a limited energy reserve, known as teneral energy, and quickly seek additional sources, such as nectar and blood meals, to survive and reproduce [39, 39–42]. Excess energy is stored as proteins, glycogen, and lipids, which support essential activities such as reproduction and flight [43–45]. For example, glycogen primarily fuels short-term flights, whereas lipids sustain longer flights [46]. Here, we investigated the mechanisms underlying insecticide resistance reversal by exploring changes in metabolic insecticide resistance, immune gene expression, and energy reserves in fungus-infected mosquitoes.
Methods
Mosquito strain
For the bioassays, we used three mosquito types: (i) pyrethroid-resistant An. coluzzii VKPER strain; (ii) insecticide-susceptible An. gambiae s.s. Kisumu, and (iii) the wild-caught strain An. gambiae s.l. The laboratory pyrethroid-resistant An. coluzzii VKPER colony was established from mosquitoes collected at Valley du Kou in Burkina Faso and selected continuously to maintain the kdr gene fixed [13, 47]. All strains were maintained and made available at the Institut de Recherche en Science de la Santé (IRSS), Burkina Faso. Larvae of An. gambiae s.l. were collected in October and November 2023 from several natural breeding sites in Soumousso and Valley du Kou and brought to the insectary at the IRSS in Bobo Dioulasso, where they were reared to adulthood. The larvae were fed with Tetramin fish food daily. Upon emergence, the adults were morphologically identified using an identification key, and only An. gambiae s.l. were maintained. All mosquito strains were maintained in the insectary under the same optimal conditions at 27 ± 2° C, 80 ± 10% relative humidity, and a photoperiod of 12:12 (L:D) at IRSS. Only 3–5-day-old, non-blood-fed females were used for the bioassays.
Mosquito infection
The Metarhizium pingshaense S10 strain used in all infections in this study was previously isolated from wild An. gambiae s.l. collected in an inhabited house in Soumousso (11°04ʹN, 4°03ʹW), Burkina Faso [28]. Here, mosquito infections were carried out using spore suspension of 14–30 day culture suspended in 0.05% Tween (Polysorbate Tween^®^ 80) at a final concentration of 10^7 ^spores/ml. Conidial concentration was determined using a Neubauer hemocytometer. The fungal suspension was mixed vigorously by using a vortex mixer, and then 0.25 ml of the suspension was sprayed on 80 mosquitoes in the treatment group, whereas the control group was sprayed with 0.25 ml of conidia-free 0.05% Tween solution using an atomizer. Mosquitoes were first anesthetized at -20 °C for 15 s and then sprayed with the fungal solution or the same volume of 0.05% Tween 80 solution for the control before their recovery. After the infection, mosquitoes were allowed to wake up in cups, transferred to small cages, and provided with 5% glucose solution.
Sampling and experimental procedures
For each of the three replicates, 15 mosquitoes were randomly selected from each treatment group (Metarhizium-infected and control) at 1 h, 4 days, and 8 days post-infection. Mosquitoes were then briefly anesthetized at − 20 °C for 5 min, transferred to dry, labeled Falcon tubes, and immediately stored at − 80 °C for subsequent energy reserve quantification assays. A sampling protocol was conducted for An. coluzzii and An. gambiae Kisumu (Fig. 1A) but not wild An. gambiae s.l.Fig. 1. Schematic representation of the experimental design. A: illustrates energy quantification experiment; a uninfected control, b Metarhizium control group. B: illustrates gene expression experiment; a control, b deltamethrin only, c Metarhizium only, and d combination of Metarhizium and deltamethrin
In addition to Metarhizium infection, mosquitoes used for gene expression quantification were also exposed to deltamethrin at 1 h, 4 days, and 8 days post-infection using the WHO insecticide susceptibility tests. This involved exposing 20–25 Metarhizium-infected mosquitoes to deltamethrin or control for 1 h duration. The mosquitoes were then transferred to insecticide-free observation tubes for an additional hour [48], after which mortality was recorded. Surviving mosquitoes were then pooled into groups of five and placed into 1.5 ml tubes containing 1 ml of RNAlater for subsequent gene expression analysis. This timeframe was chosen to preserve RNA integrity for the subsequent qPCR analysis of gene expression responses. The tubes were stored at 4 °C for 24 h to allow RNAlater to penetrate the tissues before being stored at -80 °C until RNA extraction (Fig. 1B). In both experiments, samples collected at 1 h (day 0) were used as the baseline for metabolite and gene expression quantification.
Genes expression
RNA extraction
Total RNA was extracted from 12 pools of five mosquitoes per treatment group using the PureLinkTM RNA Mini Kit (Invitrogen) following the manufacturer’s instructions. RNA concentration was measured using a Tecan Infinite PRO fluorescence microplate reader. Samples with RNA concentration exceeding 100 ng/µl were diluted to 100 ng/µl before using them for reverse transcription.
Complementary DNA synthesis
For cDNA synthesis, a reverse transcription reaction was performed with 2 μl of buffer, 0.8 μl of dNTP mix, and 2 μl of random primers. Then, 1 μl of reverse transcriptase enzyme and 1 μl of RNase inhibitor were added. The volume was adjusted to 10 μl with nuclease-free water. An RNA sample of 10 μl was then added, resulting in a final reaction volume of 20 μl. The thermal protocol included primer annealing at 25 °C for 10 s, DNA polymerization at 37 °C for 2 min, and reverse transcriptase deactivation at 85 °C for 5 s, with a final hold at 4 °C. The resulting cDNA was diluted 1:10 before use in qPCR assays.
qPCR assays
The expression levels of four genes, CYP6P3, CYP6Z1, GSTe2, associated with insecticide metabolic resistance, and defensin associated with mosquito immune defense against pathogens, including bacteria and fungi, have been quantified [16]. The housekeeping gene encoding ribosomal protein S7 (RPS7) [16] was measured and used as a reference for overall gene expression. The qPCR reactions were run in volumes of 15 μl, containing 5 μl of template nucleic acid extract and 10 μl of master mix of specific primers. Primers were supplied by Integrated DNA Technologies, Inc. (1710 Commercial Park, Coralville, IA 52241, USA). All reactions were performed on Applied Biosystems, ViiA™ 7 Real-Time PCR System with 96-Well Block, Thermo Fisher Scientific, in MicroAmp^®^ Fast 96-Well Reaction Plate (0.1 ml) from Life Technologies Corporation^™^. The thermal protocol started with an initial denaturation at 95 °C for 1 min, followed by 40 cycles of 95 °C for 5 s, 60 °C for 10 s, and 72 °C for 30 s. A final extension at 65 °C for 30 s completed product synthesis. The procedure ended with a melt curve analysis: 95 °C for 30 s, 65 °C for 30 s, and 95 °C for 30 s, to confirm amplification specificity and result accuracy. The primers used in this study are listed in Table 1. These primers were designed specifically to target the conserved regions of each gene across both species. The corresponding AGAP gene identifiers for each primer pair are listed in Table 2. Table 1. Primer sequences of target genes in Anopheles gambiae and Anopheles coluzziiGeneCodeForwardReverseS7AGAP009613GAGTCGGAGGAGGAAAAGGCCAACAGTGGGCGACAGTTTTCYP6P3AGAP002865CTGTCTGTACGAGCTGGCAAGTCGTACGTCACCTCTCCACCYP6Z1AGAP008219GCGGCCAATGTGTTTCTGTTGCCTCAGCATTGTGCGTAAGGSTe2AGAP009194CCAAAGCATTGGGCTTGGAGAGCACCGGGATCGTATGTTGDefensinAGAP007200AACATTCACTGTCGCGGGTACTTTCATCGCTGCCGGTTTGTable 2Pairwise comparison of mortality rate in wild An. gambiae s.l. and An. coluzzii after exposure to either control, deltamethrin, fungi, or mix (deltamethrin & fungi together)ComparisonsOdds ratioSEp-valueControl/deltamethrin0.0080.008 < 10^–3^Control/metarhizium0.0460.0470.006Control/mix0.0010.001 < 10^–3^Deltamethrin/metarhizium5.4181.360 < 10^–3^Deltamethrin/mix0.1480.018 < 10^–3^Metarhizium/mix0.0270.006 < 10^–3^
Energy reserves quantification
Extraction of lipid, glycogen, and sugar from mosquito pools
Energy reserves were quantified in 18 pools of five mosquitoes per treatment group according to the Van Handel protocol [49–51]. This protocol requires the separation of lipids, glucose, and glycogen. Pools of mosquitoes were homogenized in 200 µl of sodium sulfate using plastic pestles until no identifiable parts remained. A 1200 µl solution of chloroform/methanol (1:1) was then added to the homogenate and mixed. The homogenate was centrifuged at 3000g for 1 min. The supernatant containing lipids and glucose was transferred to a clean centrifuge tube, and the pellet was retained in the tube for glycogen analysis. Distilled water (600 µl) was added to the supernatant and mixed before centrifugation at 3000g for 1 min. The top phase was used for glucose quantification, whereas the bottom phase was used for lipid analysis. For heating purposes, both phases were transferred to glass tubes marked at the 5 mL level.
Glucose analysis
Glucose extracts were heated at 110 °C to evaporate the water/methanol solvents down to 200 µl before adding anthrone up to the 5 mL level mark. The solution was mixed, heated for 17 min at 110 °C, and removed from the heating block for cooling. The absorbance of the resulting solution was measured at 625 nm in duplicate. The glucose concentration was determined by comparing the absorbance to that of a standard made from 100 mg/100 ml of anhydrous glucose in deionized water.
Glycogen analysis
Anthrone was added to the pellet at the 5 ml level, then mixed and vortexed. The homogenate was heated to 110 °C for 17 min. The absorbance of the resulting solution was measured at 625 nm in duplicate. The glycogen concentration was obtained by comparing the absorbance to a standard consisting of 100 mg per 100 ml of anhydrous glucose in deionized water.
Lipids analysis
Lipid extracts were heated at 110 °C to evaporate the chloroform completely. Once dried, 200 µl of sulfuric acid was added, and the solution was heated at the same temperature for 10 min. Vanillin was then added to the 5 ml mark and mixed. The tube was then removed from the heating block and allowed to cool to room temperature. The absorbance of the resulting solution was measured at 625 nm in duplicate. The lipid concentration was determined by comparing the absorbance with that of the vegetable oil.
Statistical analysis
The susceptibility of infected mosquitoes to deltamethrin was analyzed using a binomial Generalized Linear Model (GLM). The dependent variable was the proportion of mosquitoes dead within 1 h of exposure and treatment, with mosquito strain and infection duration included as explanatory variables. For this analysis, we considered the inclusion of replicates as a random effect and interaction term among explanatory variables, but they were not supported, and hence, were not included in the final models.
Energy reserves and gene expression were analyzed using Linear Mixed Models (LMM). The initial models included treatment, infection duration, mosquito strain, and the interaction between treatment and infection duration as explanatory variables, as well as random replicates. Because of repeated amplification failure of GSTe2 in wild An. gambiae s.l., the data from this line were insufficient for analysis and were removed from the final model. Mosquito survival analysis was performed using the Cox proportional hazards (Coxph) model with treatment and mosquito strain as fixed effects and replicates as a random effect. All models were fitted to R (v4.3.3) using the package lme4 (v1.1.35.2) for LMM, and survival (v3.5.8) for Coxph. Standard tests were conducted to verify the model assumptions using the DHARMa package (v0.4.6). Inferences were made using likelihood ratio (LRT) tests.
Results
Metarhizium infection reverses insecticide resistance
To assess the phenotypic effects of Metarhizium infection on Anopheles mosquito susceptibility to insecticides, we infected Anopheles coluzzii VKPER, An. gambiae Kisumu and wild An. gambiae s.l. with Metarhizium. On days 0, 4, and 8 post-infection, the mosquitoes were exposed to deltamethrin for 1 h. Mortality was recorded 1 h after the insecticide exposure. The binomial GLM model showed significant effects of treatment, time, and strain on mosquito mortality (p < 0.001 for all the factors). Combination of deltamethrin and Metarhizium resulted in the highest mortality rate 27.5% (95% CI 24.9–30.2) followed by deltamethrin 5.4% (95% CI: 4.3–6.6), then Metarhizium 1% (95% CI: 0.6–1.6) (Fig. 2). Pairwise comparisons revealed statistically significant differences between treatments (Table 1), confirming the efficacy of the combined treatment compared to the individual applications.Fig. 2. Proportion of dead mosquitoes in the insecticide-susceptible strain An. gambiae s.s. Kisumu (left panel) and two insecticide-resistant strains An. coluzzii VKPER (middle panel) and wild An. gambiae s.l. (right panel). Mosquitoes were exposed to either a Tween solution (Control), Metarhizium suspension only (Fungi), deltamethrin 0.05% only (Deltamethrin), a Metarhizium suspension, or a combination of Metarhizium and deltamethrin (Mix)
Gene expression has complex associations with mosquito strains, treatment, and infection duration.
CYP6P3 expression
To investigate the role of CYP6P3 in the reversal of insecticide resistance by Metarhizium infection, we analyzed its expression at three time points during the course of Metarhizium infection. The best model for CYP6P3 transcription included mosquito strain (ꭓ^2^ = 18.22, df = 2, p < 0.001), time (ꭓ^2^ = 8.77, df = 2, p < 0.01), and treatment (ꭓ^2^ = 12.81, df = 3, p < 0.05). CYP6P3 transcription was significantly upregulated in An. coluzzii VKPER by 58.9% (95% CI: 28.3–89.5, p < 0.001) compared to An. gambiae Kisumu. Although deltamethrin exposure alone increased CYP6P3 transcription by 52.4% (95% CI: 21.1–83.7, p < 0.01), deltamethrin in combination with Metarhizium infection reduced CYP6P3 levels by 36.3% (95% CI: 10.9–61.7, p < 0.01) compared to deltamethrin only, suggesting that infection modulates the metabolic response to the insecticide (Fig. 3A).Fig. 3. Median expression levels (± SE) of A CYP6P3, B CYP6Z1, and C GSTe2 detoxification genes, along with D median defensin, an immune gene in Anopheles gambiae Kisumu (susceptible), Anopheles coluzzii VKPER (resistant), and wild Anopheles mosquitoes. Mosquitoes were exposed to four treatments: Control (red), Deltamethrin (gray), Metarhizium (yellow), and a combination of deltamethrin + Metarhizium (green). Measurements were taken at three time points: Day 0 (baseline for gene expression), Day 4, and Day 8 post-treatment
CYP6Z1 expression
To investigate the role of CYP6Z1 in the reversal of insecticide resistance by Metarhizium infection, we analyzed its expression at three time points after Metarhizium infection. Mosquito strain (ꭓ^2^ = 53.99, df = 2, p < 10^–6^) and treatment (ꭓ^2^ = 19.1, df = 3, p = 0.0002) had significant effect on CYP6Z1 expression level. CYP6Z1 transcription was respectively 47.7% (95% CI: 30.9–64.5, p < 0.001) and 67.3% (95% CI: 50.5–84.1, p < 0.001) higher in An. coluzzii and wild An. gambiae s.l. than in An. gambiae Kisumu, suggesting a potential role of this gene in metabolic insecticide resistance.
In mosquitoes exposed to deltamethrin alone, CYP6Z1 transcription increased by 35.8% (95% CI: 18.6–52.9, p < 0.001) compared to An. gambiae Kisumu. However, in those previously infected with Metarhizium, subsequent deltamethrin exposure increased CYP6Z1 transcription only by 21.8% (95% CI: 4.4–39.3, p < 0.05), highlighting that Metarhizium can alter the metabolic response to the insecticide exposure (Fig. 3B).
GSTe2 expression
To investigate the role of the GSTe2 gene in the reversal of insecticide resistance by Metarhizium infection, we analyzed its expression at three time points during the course of Metarhizium infection. GSTe2 transcription was impacted by mosquito strain (ꭓ^2^ = 5.10, df = 1, p = 0.02) and the interaction between treatment and infection duration (ꭓ^2^ = 12.91, df = 6, p = 0.04). The insecticide-resistant An. coluzzii, GSTe2 levels were lower by 17.4% (95% CI: -32.5–2.4, p < 0.05) compared to the reference insecticide-susceptible strain An. gambiae s.s. Kisumu.
In mosquitoes exposed to deltamethrin only, GSTe2 transcription was 73.2% (95% CI: 28.0–118.3, p = 0.0033) and 34.8% (95% CI: -10.3–79.9, p = 0.15) higher on days 4 and day 8, respectively, than their corresponding control groups (control days 4 and control day 8). However, when mosquitoes were infected with Metarhizium and exposed to deltamethrin, GSTe2 transcription was only 39.0% (95% CI: -6.2–84.2, p = 0.11) and 5.7% (95% CI: -39.5–50.8, p = 0.81) higher on days 4 and 8 compared to the within group control day 0, suggesting that Metarhizium infection modulates GSTe2 expression (Fig. 3C).
Defensin expression
To investigate the role of defensin in the reversal of insecticide resistance by Metarhizium infection, we analyzed its expression at three time points during the course of Metarhizium infection. The best model for defensin transcription included mosquito strain (ꭓ^2^ = 20.92, df = 2, p < 0.001) and treatment (ꭓ^2^ = 9.16, df = 3, p = 0.02). The defensin gene was overexpressed in An. coluzzii VKPER, and wild An. gambiae s.l. compared to An. gambiae Kisumu. The estimates were 35.4% (95% CI: 18.6–52.2, p = 0.00006) and 37.9% (95% CI: 21.1–54.8, p = 0.000018), respectively. In contrast, defensin transcription was reduced in mosquitoes treated with a combination of Metarhizium and deltamethrin by 22.7% (95% CI: 5.2–40.2, p = 0.0123742) compared to untreated An. gambiae Kisumu, similarly to deltamethrin alone (Fig. 3D).
Fungal infection depletes energy reserves
To assess the effect of Metarhizium infection on mosquito fitness, we quantified key energy levels and reserves, including glucose, glycogen, and lipids. The experiments were conducted using insecticide-resistant An. coluzzii VKPER and insecticide-susceptible An. gambiae Kisumu at the three time points.
Glucose quantification
Only infection duration (time) was retained as significant in the final model (ꭓ^2^ = 11.98, df = 2, p < 0.005). Across both treatments (control and fungi) and mosquito strains (Kisumu and VKPER), a consistent decline in glucose concentration was observed over time. Specifically, glucose concentration decreased by 15.5% (SE = 5.425, p = 0.005) 4 days and 17.6% (SE = 5.425, p = 0.001) 8 days after Metarhizium infection compared to the baseline (day 0). These results indicate that Metarhizium infection does not affect glucose concentration, even during infection (Fig. 4A).Fig. 4. Changes in metabolite concentrations (± SE) in Anopheles mosquitoes following Metarhizium infection. A Glucose, B Glycogen, C Lipid concentrations were measured in the insecticide-susceptible strain Anopheles gambiae Kisumu (left panels) and the insecticide-resistant strain Anopheles coluzzii VKPER (right panels) after Metarhizium infection, and D mosquito survival over 14 d after Metarhizium infection. Metabolite levels were assessed at three time points: Day 0 (baseline for metabolite concentration), Day 4, and Day 8 post-treatment. The x-axis represents the treatments, and the y-axis indicates metabolite concentrations (µg/µL). Error bars denote the standard error of the means
Glycogen quantification
We found a strong interaction between fungal infection and time on glycogen concentration in mosquitoes (ꭓ^2^ = 11.288, df = 2, p = 0.003). In both the susceptible and resistant strains, the mean glycogen levels in infected mosquitoes declined significantly as the infection progressed. In the control group, glycogen levels remained the highest, averaging 118.58 µg/L (95% CI: 90–147, p < 0.00001). However, in the infected group, glycogen levels decreased significantly by 77.0% (95% CI: 27.6–126.4, p = 0.003) on day 4 post-infection and by 66.5% (95% CI: 17.1–115.9, p = 0.01) on day 8. This revealed a progressive depletion of energy reserves in the mosquitoes as the infection duration increased (Fig. 4B).
Lipid quantification
The interaction between fungal infection and infection duration was retained as significant in the final model (ꭓ^2^ = 10.318, df = 2, p = 0.005). In both the susceptible and resistant strains, the mean lipid levels in infected mosquitoes significantly decreased as the infection progressed. Lipid level in the control group remained the highest, averaging 243.34 µg/L (95% CI: 206.6–280.1, p < 0.00001). However, in the infected group, lipid levels decreased significantly by 46.2% (95% CI: 16.1–76.3, p = 0.004) on day 4 post-infection and by 38.4% (95% CI: 8.3–68.5, p = 0.01) on day 8 (Fig. 4C) compared with baseline day 0.
Survival
The Hazard Ratio of dying in mosquitoes infected with fungi was 7.503 (95% CI: 5.38–10.45) times higher compared to non-treated control mosquitoes. HR for laboratory insecticide-resistant An. coluzzii VKPER and field caught mosquitoes were 1.35 (95% CI: 0.98–1.85) and 1.48 (95% CI: 1.08–2.01), respectively. These results indicated a significant increase in hazard compared to the reference insecticide-susceptible An. gambiae ss Kisumu (Fig. 4D).
Discussion
This study provides new insights into the mechanisms underlying insecticide resistance reversal via fungal infections in mosquitoes. In particular, fungal infection impairs the expression of detoxifying genes and reduces mosquito energy reserves, hinting at a dual mechanism by which Metarizhium can re-establish susceptibility, even in highly pyrethroid-resistant mosquitoes.
The results of this study revealed the complex interactions among fungal infections, insecticide exposure, and gene expression in mosquitoes. The upregulation of CYP6P3 and CYP6Z1 in An. coluzzii VKPER, and wild An. gambiae s.l. compared to the insecticide-susceptible An. gambiae ss Kisumu confirmed a strong association between these genes and insecticide resistance in malaria mosquitoes [15, 16, 47, 52, 53]. In contrast, GSTe2 transcription was lower in An. coluzzii VPER compared with An. gambiae s.s. Kisumu. GSTs are typically associated with detoxification of DDT, whereas metabolic resistance to pyrethroids is often associated with cytochrome P450 monooxygenases and esterases [53, 54]. As An. coluzzii VKPER was selected for pyrethroid resistance, it might be useful to confirm whether the VKPER strain exhibits cross-resistance to carbamates or whether it has elevated GST activity despite its selection history. Otherwise, a strain specifically selected for carbamate resistance would be more appropriate.
The upregulation of defensin transcription in insecticide-resistant An. coluzzii VKPER, and wild An. gambiae s.l. than in the susceptible An. gambiae Kisumu suggested that defensin may be involved in the immune response to environmental stressors, including insecticide exposure and fungal infections. Defensin is a key antimicrobial peptide that plays an important role in the immune defense of mosquitoes, particularly in response to microbial infections such as bacterial and fungal infections [55].
Interestingly, Metarhizium infection led to a decrease in CYP6P3 transcription, a key gene involved in insecticide detoxification. Fungal infections potentially weaken the detoxification capacity of mosquitoes, as suggested by Lamy et al. [35]. This hypothesis was supported by Reingold et al., who demonstrated that infection with Metarhizium brunneum affects gene transcription in aphids [56]. Although deltamethrin treatment induced significant increases in CYP6P3 and CYP6Z1 transcription, the combination of Metarhizium and deltamethrin had only a moderate effect. This result suggests that Metharhizium infection inhibits CYP6P3 and CYP6Z1 transcription in mosquitoes, even when they are exposed to deltamethrin.
GSTe2 expression was higher in response to deltamethrin exposure, but the effect was less pronounced in Metarhizium infection. These findings highlight the role of detoxification genes in the insecticide resistance of malaria vectors. These results demonstrate the potential of Metarhizium infection to inhibit metabolic insecticide resistance mechanisms. Metarhizium pingshaense can therefore act as a synergist, enhancing the efficacy of deltamethrin, and making it a valuable tool for integrating malaria vector control strategies in the current context of widespread insecticide resistance.
This study revealed that fungal infection significantly reduced glycogen and lipid levels in mosquitoes as well as their survival. We showed that glycogen and lipid concentrations in Metarhizium-exposed mosquitoes were significantly lower than in control mosquitoes, but this was not the case for glucose concentration, regardless of the mosquito colony. The current results support our hypothesis that Metarhizium infection causes energy depletion in mosquitoes [35]. In Metarhizium anisopliae, the penetration of the insect cuticle is facilitated by high pressure within the appressoria. This is achieved through the degradation of intracellular lipids into glycerol, which accumulates and increases osmotic pressure [57]. This process may explain the observed reduction in the lipid content of the infected mosquitoes.
These findings align with those of previous studies, demonstrating that Metarhizium infection can impair mosquito behavior and fitness. For instance, Metarhizium-infected Aedes aegypti mosquitoes have been shown to fly shorter distances than their uninfected counterparts, thereby reducing their dispersal capabilities and ability to locate hosts for blood meals [58]. This reinforces the potential of Metarhizium not only as a direct pathogen, but also as a disruptor of essential behaviors that contribute to the reduction of mosquito vectorial capacity. No significant variations in glucose concentrations were observed between the treatment groups. Because both groups were given access to feed from a 5% glucose solution, this may explain the lack of significant differences in glucose levels between the groups. Additionally, glucose may originate from the conversion of glycogen and lipids [59].
The concentrations of glucose, glycogen, and lipids decreased over time, with this decrease being more pronounced in infected mosquitoes. On days 4 and 8, glucose, glycogen, and lipid levels were significantly lower than those on day 0. This suggests that as mosquitoes grow older, their energy reserves diminish, affecting their fitness compared with younger individuals, as previously demonstrated by Somé et al. [60].
Survival in Metarhizium-infected mosquitoes was significantly lower than that in controls. Previous studies have shown a similar biopesticide potential of Metarhizium fungi, including the strain S10 used in this study [28]. Wild Anopheles gambiae s.l., presumed to be resistant to insecticides, were found to be more susceptible to Metarhizium infection than laboratory-reared, insecticide-susceptible An. gambiae s.s. Kisumu strain. No difference was observed in insecticide-resistant An. coluzzii VKPER. The reduced survival observed in wild An. gambiae s.l*.* mosquitoes during this study is likely due to the combined effects of environmental stressors and field-acquired infections. Unlike laboratory-reared mosquitoes, which are bred under controlled conditions that may affect their immune responses and overall susceptibility to pathogens, such as Metarhizium, wild mosquitoes face diverse environmental pressures and pathogen exposure. These external factors may enhance susceptibility profiles and influence survival outcomes [35]. This finding is consistent with that of Howard et al. [29], who demonstrated that insecticide-resistant mosquitoes exhibit increased susceptibility to entomopathogenic fungi compared with insecticide-susceptible strains. Furthermore, in an insecticide-free environment, the kdr mutation, which confers resistance to pyrethroids, was found to enhance mosquito survival by reducing neural and behavioral excitability in resistant individuals [61, 62]. However, this is disadvantageous to Plasmodium falciparum-infected mosquitoes [61, 63].
Conclusions
This study highlights a potential mechanism by which the entomopathogenic fungus Metarhizium significantly reduces mosquito survival and reverses insecticide resistance, thereby diminishing their disease transmission potential. Beyond reducing survival, Metarhizium infection impairs mosquito fitness by depleting key energy reserves, including glycogen and lipids, and by altering metabolic and immune gene expression. Our findings further demonstrate that the longer the infection persists, the greater is the impact on energy reserves, intensifying the physiological effect on the host. These results underscore the multifaceted effects of Metarhizium infection on mosquito biology and offer valuable insights into its potential as a biological control agent. Metarhizium-based strategies may play a pivotal role in the integrated vector management strategies aimed at reducing malaria transmission.
Author contributions
LDL, EB, and FB conceived and designed the study. LDL implemented the study. LDL, MV, FB, and EB analyzed the data and wrote the original draft of the manuscript. EB, NM, and DA revised the final manuscript. All the authors have read and approved the final version of the manuscript.
Funding
This work was supported by the Wellcome Trust grant, Ref 218771/Z, and the National Institute for Health Research (NIHR) (using the UK’s Official Development Assistance (ODA) Funding). LDL is supported by the Ministry of Higher Education Research and Innovation of Burkina Faso and by the Wellcome Trust grant, Ref 218771/Z, and the National Institute for Health Research (NIHR) (using the UK’s Official Development Assistance (ODA) Funding). MV is funded by the European Research Council, European Union’s Horizon 2020 Research and Innovation Programme grant 852957. FB was supported by an Academy of Medical Sciences Springboard Award (ref: SBF007–100094). The proofreading fees for this manuscript were supported by the African Research Initiative for Scientific Excellence (ARISE) grant (ref: ARISE-PP-FA-143) awarded to Dr. Etienne Bilgo. The ARISE programme is funded by the European Union and implemented by the African Academy of Sciences, in partnership with the African Union Commission and the European Commission.
Data availability
Data supporting the conclusions of this article are included within the article and its additional files.
Supplementary Information
Supplementary Material 1Supplementary Material 2Supplementary Material 3Supplementary Material 4
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Clinton Health Access Initiative. The PBO Net Deep Dive Report. Annual, CHAI. https://allianceformalariaprevention.com/wp-content/uploads/2022/03/PBO-net-deep-dive_Summary-of-findings-2021-report.pdf. 2021. Accessed 12 May 2025.
- 2Sibanda MM, Focke WW, Heyman HM, Birkholtz L-M. Physical, chemical and biological strategies to combat malaria. Univ Pretoria. https://www.up.ac.za/media/shared/Legacy/sitefiles/file/44/1026/2163/8121/innovate 8/4045 physical_chemical_and_biological_strategies_to_combat_malariabymthokozisimsibandaetal.pdf. 2018. Accessed 8 Jun 2025.
- 3WHO. Standard operating procedure for testing insecticide susceptibility of adult mosquitoes in WHO tube tests. https://www.who.int/publications/i/item/9789240043831. Accessed 5 Apr 2025.