B chromosome retrotransposed sequences persist through speciation, contributing to genomic and regulatory innovations in the fish genus Psalidodon (Characiformes, Acestrorhamphidae)
Lucas F. Lasmar, Mateus R. Vidal, Pamela C. F. Nadai, Rodrigo Zeni dos Santos, Raquel da Costa Machado, Ricardo Utsunomia, Francisco J. Ruiz-Ruano, Alexander Suh, Claudio Oliveira, Duilio M. Z. A. Silva, Fausto Foresti, James Crainey, James Crainey, James Crainey

TL;DR
This study explores how B chromosomes in Psalidodon fish retain and evolve retrotransposed gene copies, influencing gene regulation and contributing to genomic diversity.
Contribution
The study reveals that B chromosomes in Psalidodon fish retain and diversify retrotransposed pseudogenes, impacting gene regulation and genomic innovation.
Findings
Retrotransposition of sbno2 and simc1 genes occurred in an ancestral B chromosome, with distinct evolutionary trajectories in different species.
High-copy B chromosomes show multiple sbno2-B and simc1-B pseudogene copies, suggesting structural diversity.
Underexpression of sbno2 and simc1 in ovaries of Psalidodon paranae suggests B chromosome pseudogenes influence gene regulation via non-coding RNA.
Abstract
B chromosomes are supernumerary genetic elements rich in repetitive DNA. Many species within the fish genus Psalidodon possess a large metacentric B chromosome that exhibits signs of recent retrotransposon activity, resulting in truncated pseudogenic copies of standard A chromosomes genes, specifically sbno2 and simc1. We aimed to characterize the structure of the B chromosome pseudogenes sbno2-B and simc1-B and their evolutionary history in four B chromosome variants of three Psalidodon species, as well as their expression patterns in Psalidodon paranae individuals with a single B chromosome (1B). Our findings suggest that the retrotransposition process of each gene occurred in an ancestral B chromosome, which later diverged into distinct trajectories within each species following speciation. In the high-copy B chromosomes, the sbno2-B pseudogene shows dozens of interspersed copies,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
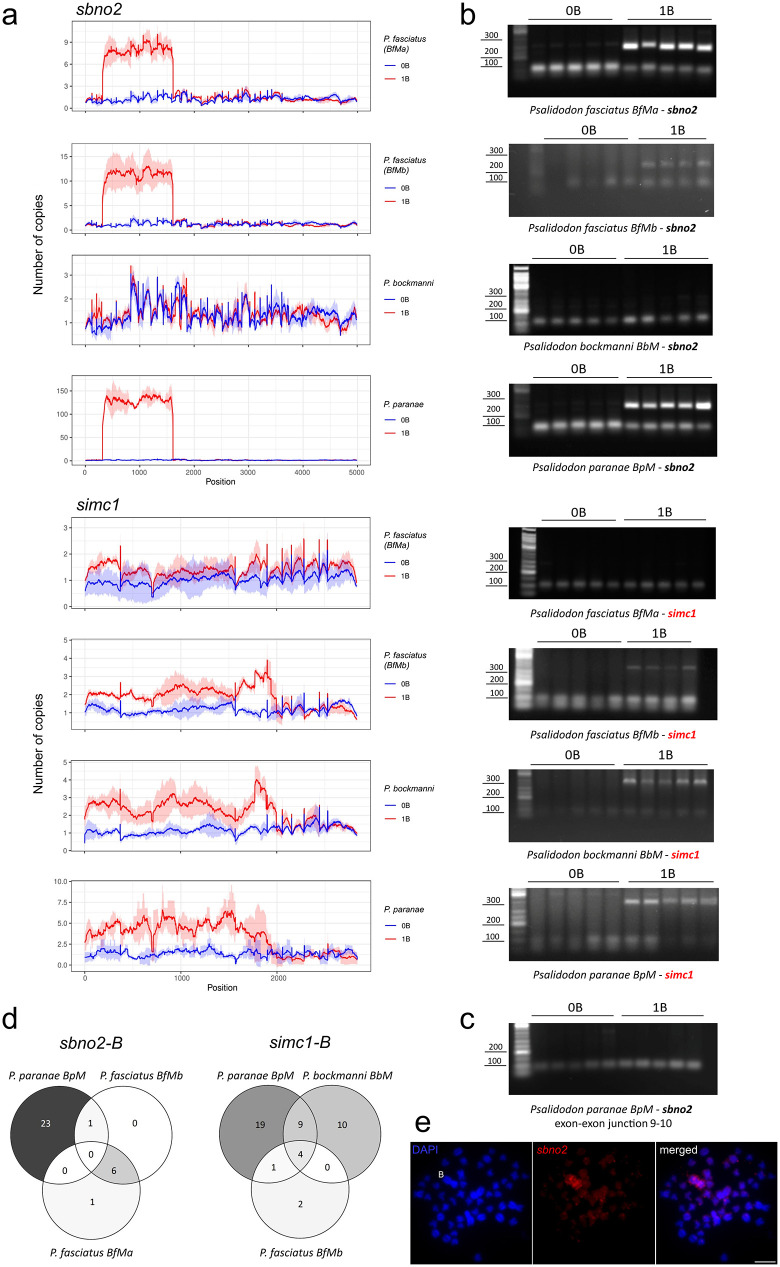 Fig 1
Fig 1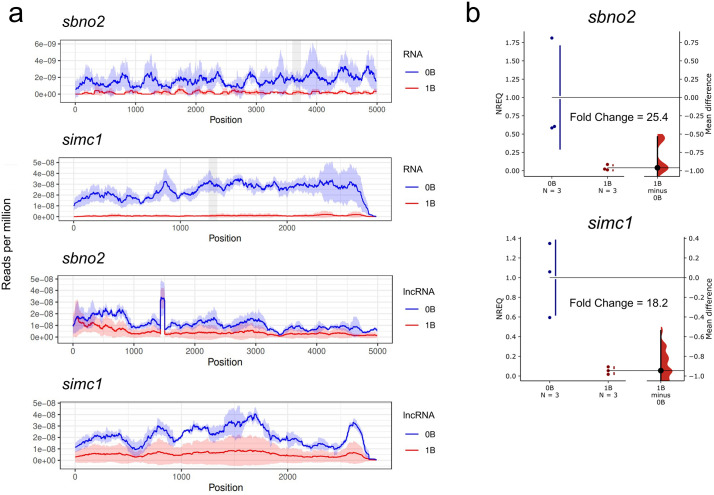 Fig 2
Fig 2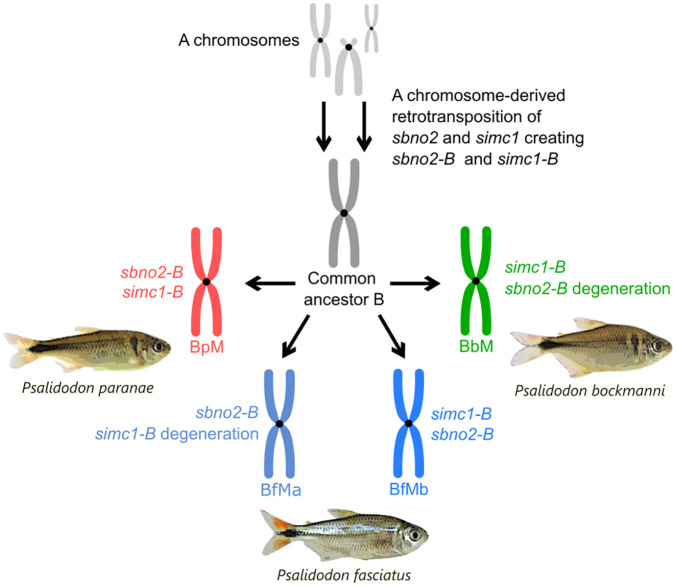 Fig 3
Fig 3- —http://dx.doi.org/10.13039/501100001807Fundação de Amparo à Pesquisa do Estado de São Paulo
- —http://dx.doi.org/10.13039/501100001807Fundação de Amparo à Pesquisa do Estado de São Paulo
- —http://dx.doi.org/10.13039/100010663H2020 European Research Council
- —http://dx.doi.org/10.13039/100010663H2020 European Research Council
- —http://dx.doi.org/10.13039/501100004359Vetenskapsrådet
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromosomal and Genetic Variations · Developmental Biology and Gene Regulation · Fish biology, ecology, and behavior
Introduction
B chromosomes and retrotransposons are distinct genetic elements that share a remarkable ability to selfishly take advantage of biological processes, thereby increasing their transmission to future generations through different molecular mechanisms [1,2]. B chromosomes are supernumerary genetic elements that primarily cheat by preferentially being incorporated into generative cells during meiosis [2]. However, in some species, B chromosomes are also known to exhibit drive (biased transmission) mechanisms at pre-meiotic and post-meiotic stages [2]. In contrast, retrotransposons are genomic sequences that over-replicate during cell cycles [1]. Retrotransposons jump to new genomic locations through a copy-and-paste mechanism, utilizing the cell’s transcriptional machinery [3], and some retrotransposons can accidentally retrotranspose mRNAs of host genes [4]. Because retrotransposition involves an mRNA intermediate, retrotransposed genes typically lack introns and regulatory elements [5]. A key feature of the retrotransposed sequences is the presence of exon-exon junctions in the genomic DNA (gDNA) copies. Intriguingly, retrotransposons can hitch a ride on B chromosomes, contributing to the latter’s increased accumulation in populations [6–8].
With the increasing accessibility of technologies for high-throughput sequencing methods, pseudogenes have been identified on B chromosomes of several eukaryote species [9,10]. On the other hand, few studies have shown the presence of retrotransposed pseudogenes on the B chromosome. The only research conducted on this topic thus far is by Carmello et al. [11], who demonstrated in fishes, the hnRNP Q-like pseudogene was retrotransposed onto the B chromosome of Astatotilapia latifasciata.
The genus Psalidodon Eigenmann, 1911 within the order Characiformes includes about 30 species, at least eight of which have B chromosomes [12]. These B chromosomes are highly morphologically diverse from micro to macrochromosomes with varying shapes. The most common variant in the genus is the large metacentric B chromosome [12]. The metacentric B chromosome in at least four Psalidodon species is believed to have originated from a common ancestor approximately 4 million years ago [13]. These B chromosomes share several genes related to their evolutionary success, including those involved in cell-cycle regulation, chromosome segregation, and sexual development [13]. Additionally, they also contain truncated pseudogenic copies of functional genes, such as the amhr2 [13]. A deeper look into the data of Silva et al. [13], revealed that short-read sequences from B chromosome-carrying samples (1B) in Psalidodon paranae, mapped to exon-exon junctions of the genes sbno2 and simc1, suggesting the presence of retrotransposed pseudogenes (sbno2-B and simc1-B) on the B chromosome.
The goal of this study was to investigate the evolutionary history of these two pseudogenes, sbno2-B and simc1-B, on the B chromosomes in Psalidodon species, as well as their potential effects on gene expression. Our results show that the sbno2-B and simc1-B pseudogenes were retrotransposed onto an ancestral Psalidodon B chromosome. These pseudogenic B copies survived through speciation events and followed independent evolutionary paths in each species. Furthermore, we observed that the functional canonical sbno2 and simc1 genes are underexpressed in 1B individuals of P. paranae, suggesting that the pseudogenic copies on the B chromosome influence the expression of the canonical genes of the A chromosomes, possibly interfering with molecular pathways for their own benefit.
Materials and methods
Sampling and cytogenetic analyses
We analyzed samples of P. paranae, P. bockmanni, and P. fasciatus from four populations with B chromosomes from São Paulo State, Brazil. The samples, localities, and B chromosome types are described in the S1 Table in S1 Table. After analysis, we deposited the specimens in the fish collection of the Fish Biology and Genetics Laboratory (LBGP) at UNESP, Botucatu, São Paulo, Brazil under vouchers described in S1 Table in S1 Table.
The animals were collected in accordance with the precepts of Law 11,794 of October 8, 2008, with Decree 6,899 of July 15, 2009, as well as guidelines set forth by the National Council for the Control of Animal Experimentation (CONCEA), and was approved by the Ethics Committee on the Use of Animals of the São Paulo State University/Institute of Biosciences (IBB/UNESP) at the meeting on 02/09/2022.
Mitotic chromosomes were obtained following the protocol established by Foresti et al. [14]. The chromosome morphology was determined based on the arm ratio method described by Levan et al. [15], classifying the chromosomes as metacentric (m), submetacentric (sm), subtelocentric (st), and acrocentric (a). To detect the partially heterochromatic B chromosomes in the individuals of each population, we used the C-banding technique originally described by Sumner [16], with some adaptations.
Genomic DNA extraction
We extracted genomic DNA from muscle and fin clips. For muscle, we used the NucleoSpin® Tissue Columns Kit with residual RNA removal by RNase A (20 mg/ml – Invitrogen). For fin clips, we used the “Hot Shot” protocol proposed by Meeker et al. [17]. The samples are described in the S2 Table in S1 Table.
Sequencing and coverage analysis
Genomic DNA (gDNA) short reads were obtained from 0B (animals without B chromosome) and 1B samples of P. bockmanni and specimens from two populations of P. fasciatus using the BGISEQ-500 platform (BGI Shenzhen Corporation, Shenzhen, China). The sequencing information and used libraries are summarized in S3 Table in S1 Table. For P. paranae, we used the gDNA short reads obtained by Silva et al. [13]. Long non-coding RNAseq reads of P. paranae were obtained from dos Santos et al. [18].
For the DNA and RNA coverage analysis, the reads were first checked for quality and adapters were removed with Trimmomatic [19], then the mapping of the sbno2 and simc1 sequences was carried out as described in Silva et al. [13] with the SSAHA2 software [20] and a minimum alignment score of 40 and 80% of minimum identity. Finally, we used customized scripts to count the number of reads mapped as a measure of abundance and to generate the coverage graphics (https://github.com/fjruizruano/ngs-protocols/blob/master/mapping_blat_gs.py; https://github.com/fjruizruano/ngs-protocols/blob/master/coverage_graphics.py). RNA coverage analysis was performed only for P. paranae using previously obtained RNAseq libraries from Silva et al. [13].
Search for in-tandem repeated sequences and mobile elements
Due to the sbno2-B and simc1-B copy number amplification on the B chromosomes, we checked if those sequences were repeated in tandem. For this, we first selected pairs of reads showing similarity with the sbno2-B and simc1-B sequences (12853 pairs for sbno2 and 12552 pairs for simc1). Then, the selected reads were input into TAREAN [21] using standard parameters, for k-mer decomposition and reconstruction of possible tandemly repeated sequences. Additionally, we checked whether the sbno2-B and simc1-B sequences have similarities with the P. paranae satellite DNAs described in the literature [22] by running a homology search using RepeatMasker (https://github.com/fjruizruano/ngs-protocols/blob/master/rm_homology_v2.py) and a de novo assemble using Geneious Prime 2024.0.7.
Finally, to check the pseudogenes structure in long reads, we analyze PacBio long reads from a 1B sample [18] (SRA accession number: SRR11678219) similar to the sbno2-B and simc1-B sequences by a BLASTn search with e-value of 1e-10 and minimum query cover of 70%. Then, we selected the regions from each read including the pseudogene and 10 kb from each flank for annotation using Geneious. To check if the pseudogene sequences occupy different locations on the B chromosome, we selected 1000 bp of each flank of the pseudogenes on each PacBio read and performed a high sensitivity de novo assembly using Geneious software. If the flanking sequences were similar, we assumed the PacBio reads came from the same genome location. If they were distinct, we assumed they came from different locations of the B chromosome. The flanking sequences (4000 bp on each side) of the PacBio reads recovered for the simc1-B pseudogene were screened for repeats using CENSOR [23]. We did not screen the sbno2 PacBio reads due to the high number of reads obtained that would not be informative.
Quantitative Reverse Transcription PCR (RT-qPCR)
To confirm the RNA coverage analysis, we measured the expression levels of the sbno2 and simc1 genes by RT-qPCR. Quantification of expression levels of sbno2 and simc1 genes was performed using ovaries from 0B and 1B females with three biological replicates per group. Tissue manipulation, RNA extraction and RT-qPCR reactions were performed as in Silva et al. [13,24] using the primers described in the S4 Table in S1 Table. The target and reference genes were simultaneously analyzed in duplicate across two independent samples. The normalized relative expression quantity (NREQ) was calculated using the 2^-∆∆Cq^ method [25], with hypoxanthine phosphoribosyltransferase 1 (hprt1) serving as the reference gene, and results were subsequently calibrated to the mean expression level of the 0B group. Two-group comparisons were performed by the Gardner-Altman estimation plot method devised by Ho et al. [26] following Gardner and Altman’s design [27], as implemented on https://www.estimationstats.com/.
Exon-exon junction analysis
To check for the presence of exon-exon junctions in sbno2 and simc1 sequences we extracted the mapped reads in the previous coverage analysis using a custom script (https://github.com/fjruizruano/ngs-protocols/blob/master/mapping_blat_gs.py). The selected reads were then mapped against the sequence of each gene with the Geneious® 7.1.3 software using the Geneious mapper algorithm. Then, we manually checked the results for the presence of reads mapping at both sides of the exon-exon junctions. To validate this analysis, we designed primers for use in multiplex PCR reactions using a modified version of Primer3 2.3.7 [28] implemented in the Geneious® 7.1.3 software. The sets of primers for both the sbno2 and simc1 genes contain a forward primer within an exon and two reverse primers, one within an exon and another one in the exon-exon junctions (S1 Fig) (S4 Table in S1 Table). With this approach, we expect a short single amplicon in 0B samples from the A copies and two amplicons in the 1B, the short one from the A and B copies, and another longer one exclusive of the B copies. The primer pairs sets were designed to generate 84 bp and 239 bp amplicons for sbno2 and 80 bp and 319 bp amplicons for simc1. We also developed a set of primer pairs for the exon-exon junction 9–10 of the sbno2-B pseudogene for use in a regular PCR to confirm the presence of this junction in the 1B and 0B samples (S4 Table in S1 Table, S1 Fig). The expected amplicon size for this 9–10 exon junction was 76 bp (S1 Fig). PCR reactions were carried out using 15 ng of DNA, 1X PCR buffer, 1.5 mM MgCl_2_, 200 μM dNTPs, 0.1 μM of each primer and 1U Taq DNA polymerase in a final volume of 12.5 μl. Thermocycling conditions were initial denaturation at 94°C for 2 min, 35 cycles at 94°C for 45 s, 58°C for 45 s, 68°C for 30 s and a final extension step at 68°C for 10 min. Only for P. fasciatus (BfMa) from the Piratininga stream, the annealing temperature used was 66°C.
Search for B-specific Single Nucleotide Polymorphisms (SNPs) in Psalidodon species
The search for SNPs in the pseudogenes of the B chromosomes was performed using customized scripts (https://github.com/fjruizruano/whatGene and https://github.com/fjruizruano/ngs-protocols). For P. paranae we used four gDNA and six RNA libraries from Silva et al. [13] and six lncRNA libraries previously obtained from dos Santos et al. [18]. Additionally, we used six libraries for both P. fasciatus and P. bockmanni (12 in total) obtained in the present study (S3 Table in S1 Table). We mapped each library against sbno2 and simc1 sequences using “mapping_blat_gs.py” script (https://github.com/fjruizruano/ngs-protocols/blob/master/mapping_blat_gs.py), with SSAHA2 option (v.2.5.5) [20]. Then, using SAMtools (v.1.3.1) [29] we SNP calling to obtain putative SNPs exclusive from the 1B libraries. After putative merged the bam files in each respective group, i.e., 0B gDNA, 1B gDNA, 0B RNA, 1B RNA, 0B lncRNA, and 1B lncRNA; and separately for each species. After merging, we used the “bam_var_join.py” script (https://github.com/fjruizruano/whatGene/blob/master/scripts/bam_var_join.py) to obtain the counts per position of each nucleotide, and insertions/deletions. Then, with the “snp_calling_bchr.py” script (https://github.com/fjruizruano/whatGene/blob/master/scripts/snp_calling_bchr.py) we performed SNP selection for each group, we extracted counts of the Ref (reference) and Alt (alternative) alleles for each library. Then, for each type of library (gDNA, RNA and lncRNA) we used two filters in the putative SNPs. In the first filter, we only kept the SNPs with the occurrence of Alt alleles in all 1B individuals of the group. For the second filter, we obtained the mean value of occurrence in each position for the 0B libraries. Considering the presence of two copies of each gene on the A chromosomes, and at least one copy on the B, we discarded all the Alt alleles with occurrence of less than half of the 0B mean.
Fluorescence in situ Hybridization (FISH)
The FISH probe for the sbno2 gene was generated by PCR including digoxigenin-11-dUTP (Roche Applied Science) in the reaction with the primers designed by Silva et al. [13] (S4 Table in S1 Table). FISH experiments were carried out as in Pinkel et al. [30] pin, using high stringency conditions, and the signals were detected with anti-digoxigenin-rhodamine (Roche Applied Science). Chromosomes were counterstained with 4’,6-Diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, CA). Images were captured with a digital camera (Olympus DP90) attached to an Olympus BX61 epifluorescence photomicroscope and acquired using CellSens Dimension (Olympus). Image treatment, optimization of brightness, and contrast were performed using the Adobe Photoshop CS4 program. Our investigation approach is summarized in S2 Fig.
Results
When mapping short-read sequence data against the A chromosomes, the sbno2 and simc1 genes show regions with higher gDNA coverage in 1B samples across all B chromosome variants analyzed, except for the sbno2 gene on the BbM variant (metacentric B chromosome of P. bockmanni) and the simc1 gene on the BfMa (metacentric B chromosome variant “a” of P. fasciatus) (Fig 1a, S3 Fig). These high-coverage regions span portions of each of the two genes, suggesting these B-variants have truncated pseudogenic copies, referred to as sbno2-B and simc1-B, of the canonical genes on the A chromosomes. The high coverage regions among species are coincidental for each gene, i.e., the copies on the B chromosomes are truncated in the same regions for each gene (Fig 1a). Furthermore, the coverage graphs show that different B chromosome variants have distinct copy numbers of the pseudogenes (Fig 1a), ranging from eight on the BfM to 125 on the BpM for sbno2-B, and from two on the BfMb and BbM to three on the BpM for simc1-B (Fig 1a).
Detection of pseudogenes on the B chromosomes of Psalidodon species.a coverage graphs of the pseudogenes on the B chromosomes of Psalidodon species (the P. paranae graph was adapted from Silva et al., 2021). b electrophoresis agarose gels showing the amplification of B-specific pseudogenic sequences (upper band) of the genes sbno2 and simc1 on samples with B chromosomes (1B) by multiplex PCR. The lower band on all samples is a positive control (see methods section). c amplification of the exon-exon junction 9-10 in both 0B and 1B samples by regular PCR. d Venn diagram showing the number of B-specific SNPs shared among the B chromosome variants of the Psalidodon species analyzed in this study. e Metaphase of P. paranae after FISH with a probe for the sbno2 gene. Bar = 10 µm.
We also analyzed Psalidodon B chromosomes for complete copies (with introns) and non-truncated retrogene copies (without introns) of the sbno2 and simc1 genes. To do this, we mapped gDNA reads of 0B and 1B samples of the four Psalidodon populations analyzed here onto the sbno2 and simc1 full gene sequences, including introns. This mapping revealed no coverage differences in intronic regions between 0B and 1B groups across any species (S4a, b Fig), except for small areas composed of fragments of genes (S4c, d Fig, S5 Table in S1 Table). These results suggest that the Psalidodon B chromosomes only contain retrotransposed truncated copies of the sbno2 and simc1 genes, with no full gene sequences.
TAREAN clustering analysis did not detect the typical satellite DNA clusters for the sbno2 and simc1 sequences, indicating that the amplified copies of these genes on the B chromosome of P. paranae are not tandemly arranged (Supplementary Material S1, S2 Files). PacBio long read analysis confirmed this for the sbno2-B pseudogene (S5a Fig). However, for the simc1-B pseudogene, PacBio reads revealed an in-tandem structure consisting of three gene copies, consistent with the copy number estimated by short reads coverage (S5b Fig, Fig 1a). Comparison of the sbno2-B and simc1-B sequences to the P. paranae satellitome (Silva et al., 2017) [22] revealed no significant sequence similarities.
To determine whether the pseudogene sequences are located in different regions of the B chromosome, we recovered 169 PacBio long reads encompassing the sbno2-B pseudogene. Analysis of the gene flanks (1000 bp on each side) revealed more than 70 distinct sequences. Additionally, about 70 sequences assembled into a single contig, while other small groups formed separate contigs. This suggests that the sbno2-B sequences originate from different paralogs from distinct regions of the B chromosome. FISH mapping further supported this, confirming the presence of the sbno2 gene at several locations on the P. paranae B chromosome (Fig 1e), with prominent clusters symmetrically positioned on the arms and smaller clusters scattered throughout. For the simc1-B pseudogene, only five PacBio reads were recovered which assembled into a single contig with three paralog copies (S5b Fig) indicating that they derive from the same genomic sequence. FISH did not show visible signals for the simc1 gene, likely due to its low copy number on the B chromosome. Repeat screening of the simc1-B and *sbno2-*B flanking sequences did not identify the presence of repetitive elements in or near the pseudogenes with high confidence.
gDNA read mapping also revealed reads spanning the exon-exon junctions in high-coverage regions of the sbno2-B and simc1-B pseudogenes in 1B samples (S6a, c Fig) as detailed in Table 1, confirming the absence of introns on the B chromosome copies of these genes. Multiplex PCR reactions further confirmed the presence of these exon-exon junctions in the 1B samples (Table 1, Fig 1b). Additionally, gDNA reads spanning exon-exon junctions 8–9 and 9–10 were detected in a 0B individual of P. paranae (S6b Fig). Although these reads were present in only one 0B individual, PCR assays with five 0B individuals confirmed that junction 9–10 was present in all samples (Fig 1d). These PCR results, obtained from different individuals than those used for Illumina sequencing, rule out sequencing errors such as index swapping [31], confirming that in 0B samples, a region containing exons 8–10 without intervening introns was also retrotransposed to some part of genome A.
Table 1: Summary of read mapping and multiplex PCR results for exon-exon junctions in the high-coverage B-regions of the sbno2 and simc1 genes in 1B and 0B samples.
If the 0B samples have more copies of the fragments including exons 8–10, we would expect higher coverage in this region relative to its flanking regions in the coverage graphs. As expected, we observed a slightly higher coverage of this region compared to its flanks (S7 Fig).
The number of B-specific SNPs for the sbno2-B and simc1-B pseudogenes in gDNA in the high-coverage regions for each species is summarized in the Venn diagrams in Fig 1d (S8-S15 Tables in S1 Table). For both the sbno2-B and simc1-B pseudogenes, P. paranae exhibited the highest number of SNPs (24 and 33, respectively). For the sbno2-B pseudogene, P. fasciatus (BfMa) and P. fasciatus (BfMb) shared the highest number of B-specific SNPs (7 each), with no B-specific SNP common to all species. For the simc1-B pseudogene, P. paranae and P. bockmanni shared the highest number of B-specific SNPs (13), with four of these also present in P. fasciatus (BfMb). Both populations of P. fasciatus had the lowest number of B-specific SNPs for both pseudogenes.
The RNA coverage analysis for mRNA and lncRNA reads for the sbno2 and simc1 genes in 0B and 1B ovaries of P. paranae showed that both genes are underexpressed in 1B samples (Fig 2a). In contrast, RNA coverage in 0B samples was uniform throughout the coding sequences for both genes. RT-qPCR for mRNA confirmed the expression patterns for both genes (Fig 2b, S6, S7 Tables in S1 Table) with high 0B/1B fold change values, FC = 25.4 and 18.2 for the sbno2 and simc1 genes, respectively. Because of the low number of B-specific SNPs, expression levels were measured for all copies (A + B) per gene together.
Gene expression analysis of the pseudogenes on the B chromosomes of Psalidodon species.a coverage graphs show underexpression of Psalidodon genes sbno2 and simc1 in samples with B chromosomes by RNAseq and lncRNAseq analysis. The shaded areas indicate the RT-qPCR amplicon. b Gardner-Altman estimation plots showing sbno2 and simc1 transcription levels in 0B and 1B individuals, analyzed by RT-qPCR. Both groups are plotted on the left axes, and the mean difference (effect size) is plotted on a floating axis on the right as a bootstrap sampling distribution. The mean difference is depicted as a black dot, and the 95% confidence interval is indicated by the ends of the vertical error bar. NREQ = normalized relative expression quantity.
The SNP analysis using lncRNA reads did not show B-specific SNPs for the sbno2 and simc1 genes, indicating that the B chromosome does not express long non-coding sequences with B-specific variation (S16, 17 tables in S1 Table). We analyzed the mRNA reads and detected three SNPs for the sbno2 gene in the high-coverage region and zero SNPs for the simc1 gene. This strongly indicates that mRNA transcripts of the sbno2 gene from the B chromosome are expressed (S18, 19 tables in S1 Table). Considering that our filters were very stringent for this analysis (see methods section), we cannot rule out that copies of the simc1 gene are expressed from the B chromosome, since one individual exhibited SNPs on the high-coverage region of this gene, but in a low proportion compared to the 0B reference copies (S19 Table in S1 Table).
We created a quick protocol for identifying individuals bearing B chromosomes extracting DNA with the HotShot protocol and doing multiplex PCRs with A-specific and B-specific primers combined (full protocol detailed in the Methods section). Using the primers set for the sbno2 or simc1 genes, the protocol can be applied to all the B-variants analyzed here. Using this approach, the genotyping time was 3 hours, from tissue collection to results.
Discussion
Here we demonstrate that four B chromosome variants of three Psalidodon species have retrotransposed copies of the sbno2 and simc1 genes that were acquired in a common ancestor and maintained across speciation events. These findings were based on the presence of exon-exon junctions for the B copies of these genes at DNA level, confirmed by bioinformatic analyses of gDNA reads and multiplex PCR. Furthermore, the coverage analyses for the sbno2 and simc1 genes showed that the pseudogenic B copies of each gene are truncated in the same regions on the B-variants analyzed here. This similarity indicates that the sbno2-B and simc1-B pseudogenes originated on an ancestral Psalidodon B chromosome through retrotransposition events from mRNAs of the copies present on the standard A chromosomes, as illustrated in Fig 3. This evidence of retrotransposition of gene fragments to the B chromosome is reinforced by the absence of introns for the sbno2-B and simc1-B pseudogenes on the Psalidodon B chromosomes (see results). This allows us to discard the possibility of an initial origin or colonization of the B chromosome with full copies of the genes followed by internal retrotranspositions within the B chromosome.
Illustration of the most parsimonious hypothesis on how the B chromosomes of the Psalidodon species acquired and lost (in two cases) the pseudogenes sbno-2-B and simc1-B.
The metacentric B chromosomes of the genus Psalidodon share a common ancestor origin at least 4 million years ago [13]. The number of shared SNPs among each pair of Psalidodon B chromosomes here is correlated with the phylogenetic history of the genus [32], which shows P. paranae and P. bockmanni phylogenetically closer than these species with P. fasciatus. For the sbno2-B, the two P. fasciatus B chromosomes share more SNPs than with P. paranae and for simc1-B, the P. paranae and P. bockmanni B chromosomes share more SNPs than both with the P. fasciatus B chromosomes (Fig 1d, 8-S15 Tables in S1 Table). These findings support the idea that the B pseudogenes sbno2-B and simc1-B originated on an ancestral B chromosome, which persisted through speciation events in the genus, maintaining a relatively conserved structure over a long evolutionary period.
After the initial colonization of the B chromosome by the retrotransposed copies, the pseudogenic sequences likely underwent multiple rounds of amplification to achieve the higher copy numbers observed (Fig 1a). PacBio long-read analysis showed that the flanking regions of the sbno2-B belong to different parts of the B chromosome, indicating the sbno2-B was dispersed during amplification events, which was reinforced by the presence of FISH signals throughout the arms of the B chromosome. For the simc1-B, we found three sequences repeated in tandem in the PacBio long reads, which suggests that they have the potential to evolve into a satellite DNA structure. Also, for simc1-B, we observed just one group of unique sequences flanking of the pseudogene, indicating that the in-tandem sequences of this pseudogene are confined to a specific area of the B chromosome.
The BbM and BfMa variants lack any copies of the sbno2-B and simc1-B pseudogenes, respectively. The most parsimonious hypothesis to explain this absence is that these sequences would have been lost during the evolutionary history of each B chromosome within each species. Degenerative processes such as random deletions, the formation of satellite DNAs or the insertion of other transposable elements may be responsible for these losses, as observed in sex chromosomes [33–35].Interestingly, a B chromosome variant of another P. bockmanni population has the sbno2-B pseudogene [13], suggesting that the loss of the sbno2-B pseudogene on the BbM variant analyzed here is a recent event.
The finding of reads in the exon-exon junctions 8–9 and 9–10 of the sbno2 gene in 0B individuals of P. paranae indicates the presence of retrotransposed copies of this gene in the chromosomes of the standard A complement. Two hypotheses can explain this finding. The first is that, after retrotransposition of the sbno2-B copies onto the B chromosome from A chromosomes, a pseudogene fragment was moved back to the A chromosomes. The retrotransposition from the B to the A chromosomes could be facilitated by the presence of retrotransposon machinery surrounding the sbno2-B sequences on the B chromosome. This machinery would be a remnant of the initial retrotransposition process. Thus, both processes would be related. In all B chromosomes analyzed, the sbno2-B and simc1-B pseudogenes had high coverage across all exons, suggesting the entire pseudogenes were amplified. In contrast, on the A chromosomes, only two connected exon-exon junctions of sbno2-B were found, indicating that just part of the pseudogene was transferred back to the A chromosomes. The retrotransposition from the B to the A chromosomes could be facilitated by the presence of a retrotransposon machinery sequence surrounding downstream of the sbno2-B sequences on the B chromosome. This machinery sequence would be a remnant of the initial retrotransposition process.
The second hypothesis is that the retrotransposed A copies were created in an independent event of the B copies. This hypothesis is less likely because it relies on two independent retrotransposition capturing fragments of the same gene. However, this could also be facilitated by retrotransposon sequence surrounding the canonical copy of the gene in the A chromosomes. Illumina 0B reads were found mapped in exon-exon junctions just in P. paranae, indicating that the retrotransposition of the sbno2-B pseudogene in 0B animals is exclusive of this species and that it is a later event compared to the retrotransposition to the B chromosome as the B pseudogene was retrotransposed in a common ancestor B chromosome.
In this study, we measured the expression levels of the canonical sbno2 and simc1 gene, both of which showed lower expression levels in the 1B samples. This differential expression may result from the regulation of canonical gene copies by the sbno2-B and simc1-B truncated pseudogenes. Truncated copies could be involved in canonical copy post-transcriptional direct or indirect regulation by mechanisms such as small siRNAs, miRNAs or competition for transcription factors [34,36] SNPs analysis using lncRNA reads did not show expression of B copies with B-specific variation. On the other hand, we observed some B-specific transcripts in mRNA reads for the sbno2-B pseudogene, indicating that this pseudogene can express mRNA truncated copies, which could be involved in canonical copy regulation by RNAi mechanisms. How these retrocopies are expressed in a new location probably lacking their parental regulatory elements, e.g., promoters and enhancers, is an intriguing question. However, retrocopies can benefit from regulatory elements of other genes in their vicinity [37]. We did not find B-specific SNPs in mRNA reads for the simc1-B and we therefore speculate that, if expressed, the simc1-B may produce other types of RNA, such as miRNAs.
If the sbno2-B and simc1-B pseudogenes can influence the expression of their canonical counterparts, we suspect that in the populations there is lacking one of the pseudogenes on the B chromosome, such as P. bockmanni (Segredo waterfall) and P. fasciatus (Alambari stream), the expression of the standard A-copies remains unaffected. This would lead to different effects of each B in the regulatory networks for each population. We did not measure the expression of these genes in the population besides the P. paranae one due to the difficulty of obtaining an adequate sample size for the RT-qPCR analysis. Therefore, identifying populations with favorable conditions to test these assumptions would be valuable for future research.
The underexpression of the sbno2 and simc1 genes in P. paranae 1B individuals may benefit B chromosome transmission across generations by manipulating the cell cycle. As the B chromosomes of Psalidodon species lack homologs to pair during meiosis, they employ an intriguing strategy of auto-pairing and gene expression manipulation to overcome the meiotic checkpoints [23]. Insights into how the sbno2 and simc1 genes might contribute to this process can be hypothesized based on the known roles of these genes in mammals. In mammals, the Sbno2 gene is associated with cancer cell proliferation and survival [38], while the Simc1 gene encoded protein, SIMC1, interacts with the CTBP1 protein, which is involved in the ability of cancer cells to evade cell-cycle checkpoints [39]. Collectively, we propose that the underexpression of both genes in the 1B samples may be crucial for the 1B cells to evade cell-cycle checkpoints and proliferate, ensuring B transmission over generations [24,10].
Finally, we propose a new protocol for the rapid identification of Psalidodon samples carrying large metacentric B chromosomes. Our results have shown that the HotShot DNA extraction protocol combined with multiplex PCR reactions to be an efficient tool for identifying B chromosomes in the three sampled Psalidodon species. With this tool the genotyping of large numbers of 0B and 1B animals is significantly expedited, reducing the processing time from a few days using classical cytogenetic analysis, to just a few hours. Furthermore, because fin clips are used for DNA extraction, this protocol allows for the minimally invasive genotyping of live specimens, thereby minimizing the impact on the studied populations and opening new avenues for various types of studies.
In conclusion, we demonstrated that the metacentric B chromosomes of three Psalidodon species share two retrotransposed pseudogenes, sbno2-B and simc1-B, which were acquired from an ancestral B chromosome. Canonical copies of the sbno2 and simc1 genes are underexpressed in 1B animals in P. paranae, indicating that pseudogenic B copies influence their expression through non-coding RNA interference mechanisms. Fragments of the sbno2-B pseudogene were identified on the standard A chromosomes, showing that the dispersion of these genes contributes to genomic diversity in both the A and B chromosomes. We did not precisely determine whether these A-copies of the pseudogenes are present in one or more A chromosomes, or which specific chromosomes they might occupy. Future work, including whole-genome assemblies of Psalidodon species and in-depth analysis of transposable elements on the B chromosomes, will be crucial in better understanding how these pseudogenes are organized on both the A and B chromosomes. Our work would benefit from a dendrogram including the sbno2 and simc1 paralogs to investigate the origins of the pseudogenes on the B and A chromosomes. However, the sequences did not exhibit sufficient variation for this analysis.
Supporting information
S1 FigIllustration of the primer scheme for multiplex and regular PCR to confirm the presence of B chromosome-specific sequences on samples with B chromosomes.F1 = forward primer. R1 = reverse primer 1. R2 = reverse primer 2.(PDF)
S2 FigSummary of the investigation approach.(PDF)
S3 FigCoverage graph of the sbno2 gene on the B chromosomes of Psalidodon paranae excluding the high-coverage region to visualize the absence of differences between 0B and 1B samples.(PDF)
S4 FigCoverage graph of the full sbno2 (a, c) and simc1 (b, d) genes sequences of Psalidodon paranae (including introns).The high-coverage peaks observed in c, d were removed from the analysis in a, b for better visualization of the low-coverage regions. The shaded areas indicate the exon boundaries.(PDF)
S5 FigExample of Psalidodon paranae PacBio reads alignment with the pseudogene sequences annotated, sbno2 (green) (a) and simc1 (pink) (b).(PDF)
S6 FigExample of Illumina short reads mapped to a reference sequence.The dotted lines show the exon-exon junctions. Note the presence of reads crossing the exon-exon junctions in all the mappings. In b, the red boxes identify the reads crossing the exon-exon junctions for visualization purposes.(PDF)
S7 FigCoverage graph using 0B reads of two P. paranae individuals.The dotted lines show the exon-exon junctions. The sample highlighted in blue showed reads mapped in the exon-exon junctions. Note the slightly higher coverage in the blue sample on exons present in the 0B genome (green boxes) compared to its flanks (exons 7 and 11) and the red sample.(PDF)
S1 Raw imagesUncropped raw images of the electrophoresis gels used in this study.(PDF)
S1 FileTAREAN clustering results for the sbno2 reads of Psalidodon paranae.(PDF)
S2 FileTAREAN clustering results for the simc1 reads of Psalidodon paranae.(PDF)
S1 TableSupplementary tables for this study.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Werren JH. Selfish genetic elements, genetic conflict, and evolutionary innovation. Proc Natl Acad Sci U S A. 2011;108(Suppl 2):10863–70. doi: 10.1073/pnas.1102343108 21690392 PMC 3131821 · doi ↗ · pubmed ↗
- 2Houben A. B Chromosomes - A Matter of Chromosome Drive. Front Plant Sci. 2017;8:210. doi: 10.3389/fpls.2017.00210 28261259 PMC 5309253 · doi ↗ · pubmed ↗
- 3Boeke JD, Garfinkel DJ, Styles CA, Fink GR. Ty elements transpose through an RNA intermediate. Cell. 1985;40(3):491–500. doi: 10.1016/0092-8674(85)90197-7 2982495 · doi ↗ · pubmed ↗
- 4Carelli FN, Hayakawa T, Go Y, Imai H, Warnefors M, Kaessmann H. The life history of retrocopies illuminates the evolution of new mammalian genes. Genome Res. 2016;26(3):301–14. doi: 10.1101/gr.198473.115 26728716 PMC 4772013 · doi ↗ · pubmed ↗
- 5Pan D, Zhang L. Burst of young retrogenes and independent retrogene formation in mammals. P Lo S One. 2009;4(3):e 5040. doi: 10.1371/journal.pone.0005040 19325906 PMC 2657826 · doi ↗ · pubmed ↗
- 6Camacho JPM. B Chromosomes. In Ryan Gregory T, editor. The Evolution of the Genome (Capítulo 4). 2005. p. 223–86.
- 7Lamb JC, Riddle NC, Cheng Y-M, Theuri J, Birchler JA. Localization and transcription of a retrotransposon-derived element on the maize B chromosome. Chromosome Res. 2007;15(3):383–98. doi: 10.1007/s 10577-007-1135-0 17429746 · doi ↗ · pubmed ↗
- 8Coan RLB, Martins C. Landscape of Transposable Elements Focusing on the B Chromosome of the Cichlid Fish Astatotilapia latifasciata. Genes (Basel). 2018;9(6):269. doi: 10.3390/genes 9060269 29882892 PMC 6027319 · doi ↗ · pubmed ↗