Evaluation of culture- and PCR-based methods for detecting Burkholderia pseudomallei in soil samples in Thailand
Didtawa Suttisak, Charlene Mae Salao Cagape, Rathanin Seng, Natnaree Saiprom, Wongwarut Boonyanugomol, Kamolchanok Rukseree, Paul J. Brett, Mary N. Burtnick, Narisara Chantratita, Elsio Wunder Jr, Elsio Wunder Jr, Elsio Wunder Jr

TL;DR
This study compares methods to detect a soil bacterium linked to a deadly disease in Thailand, finding that enrichment culture improves detection rates.
Contribution
The study evaluates and compares culture and PCR methods for detecting Burkholderia pseudomallei in soil, highlighting the effectiveness of enrichment culture.
Findings
Enrichment in ACER broth followed by culture on Ashdown agar detected B. pseudomallei more effectively than direct culture.
Real-time PCR detected B. pseudomallei in only 2 out of 20 culture-positive soil samples.
No significant association was found between environmental detection of B. pseudomallei and human seroprevalence.
Abstract
Burkholderia pseudomallei is an environmental bacterium that causes melioidosis, a life-threatening disease prevalent in tropical regions. Accurate detection of B. pseudomallei in the environment is essential for identifying areas that pose an infection risk. This study aimed to evaluate culture- and real-time polymerase chain reaction (PCR)-based methods for detecting B. pseudomallei directly in soil samples, and to assess its environmental prevalence in relation to antibody levels in healthy individuals from Amnat Charoen, Northeast Thailand. Initial studies used spiked soil samples to evaluate culture-based methods including Ashdown agar, phosphate-buffered acidic erythritol (ACER) agar, Threonine-basal salt solution with colistin 50 (TBSS-C50) broth, TBSS-C50 based erythritol broth, and ACER broth, and real-time PCR assays targeting BPSS1187 and TTS1-orf2. Hemolysin coregulated…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
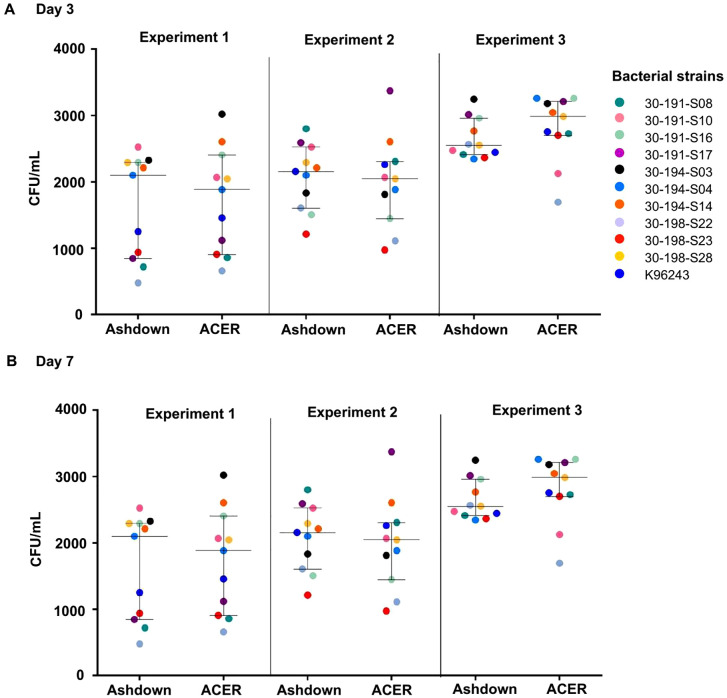 Fig 1
Fig 1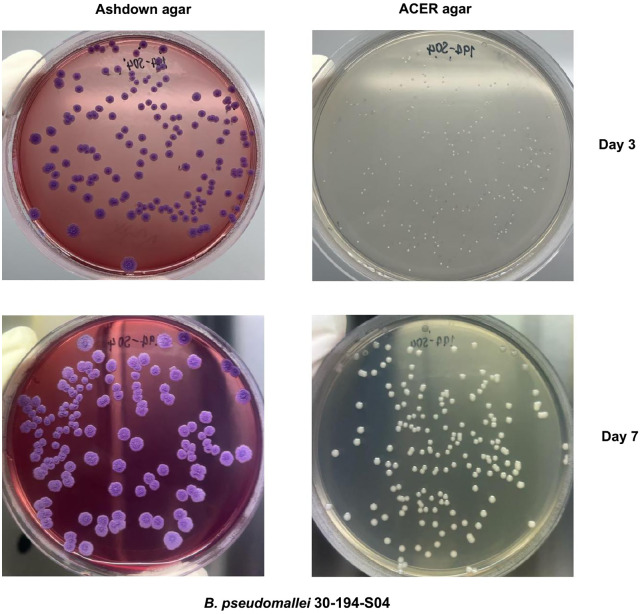 Fig 2
Fig 2 Fig 3
Fig 3 Fig 4
Fig 4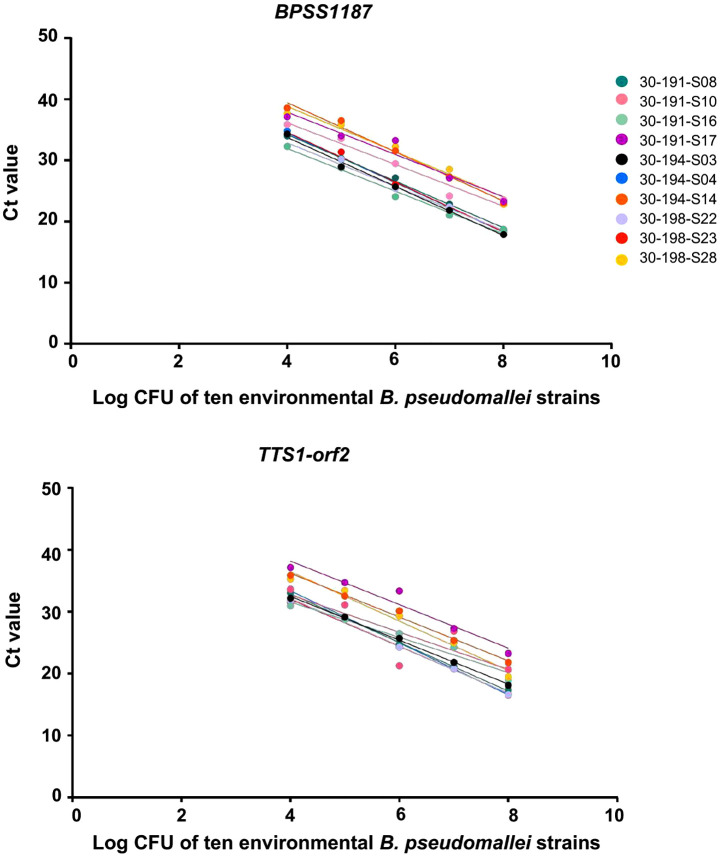 Fig 5
Fig 5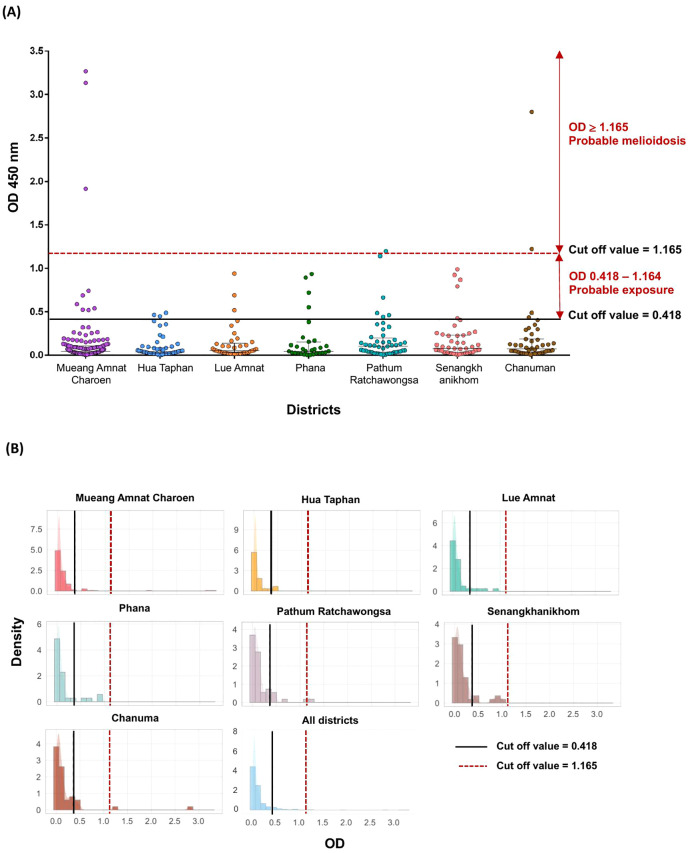 Fig 6
Fig 6 Fig 7
Fig 7 Fig 8
Fig 8- —http://dx.doi.org/10.13039/100000774Defense Threat Reduction Agency
- —http://dx.doi.org/10.13039/100010269Wellcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBurkholderia infections and melioidosis · Research on Leishmaniasis Studies · Veterinary medicine and infectious diseases
Introduction
Burkholderia pseudomallei is a Gram-negative, saprophytic environmental bacterium that causes melioidosis, a potentially fatal infectious disease affecting humans and animals [1]. This pathogen is predominantly found in moist soils, surface and ground water, and the rhizosphere in tropical and subtropical regions [2]. B. pseudomallei infections typically occur through inoculation, ingestion, or inhalation of aerosolized bacteria, with most cases reported during the wet season or following severe weather events such as tropical storms [3]. Thailand reports approximately 7,000 cases of human melioidosis annually [4] with mortality rates of approximately 25–40% [5]. The clinical manifestations of melioidosis vary in severity, ranging from asymptomatic cases to localized skin infections, pneumonia, and septicemia [6,7]. In severe cases, the disease progresses rapidly, leading to acute septicemia, septic shock, and death. Consequently, prompt and accurate diagnosis, along with timely treatment, is crucial for effective disease control and prevention.
Culture-based methods remain the gold standard for detecting B. pseudomallei in clinical samples and are also recommended for environmental detection [8]. Ashdown agar, a selective medium, is effective for isolating B. pseudomallei in regions where the disease is endemic [9]. Traditionally, detection of B. pseudomallei in soil involved enrichment in threonine-basal salt solution with colistin (TBSS-C50), followed by subculture on Ashdown agar. However, this culture method has limited sensitivity, with detection rates of approximately 9.8% after 48 hours and 25.5% after 144 hours of enrichment [10]. Improved sensitivity has been achieved by varying nitrogen and carbon sources in TBSS-C50. For example, NH_4_H_2_PO_4_ can be used as a substitute for nitrilotriacetic acid, and erythritol can replace threonine since it is a unique carbon source that is used by B. pseudomallei but not by other Burkholderia species [11]. Recently, a novel phosphate-buffered acidic erythritol (ACER) medium has been shown to enhance growth of B. pseudomallei compared to TBSS-C50 and TBSS-C50-based erythritol media in a study conducted in the north-central region of Vietnam [12]. Even with these improvements, culture methods remain limited by their low sensitivity and labor-intensive nature. To overcome these limitations, molecular methods such as real-time polymerase chain reaction (PCR) and Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) [13] have been employed for the detection of B. pseudomallei in environmental samples.
The current species-specific target used for the detection of B. pseudomallei by PCR is the orf2 gene, which is located within the type III secretion system 1 (TTS1) gene cluster [14,15]. A real-time PCR assay targeting TTS1-orf2 gene demonstrated 100% specificity in spiked human blood samples [15]. In a study using 200 soil samples collected from rice paddies in Northeast Thailand, TTS1-based real-time PCR showed the detection rate of 76.5%, and when BPSS0745 was added as a secondary target the detection rate increased to 90% [16]. Real-time PCR has consistently shown higher detection rates for B. pseudomallei in environmental samples compared to culture methods using TBSS-C50 enrichment broth [17]. Recent advancements in real-time PCR assays for B. pseudomallei includes the incorporation of an internal control to ensure the efficiency of DNA extraction and the assessment of additional target genes including BPSS0087, BPSS1187, BPSS1498 (hcp1), and BPSS1492 (bimA) [18]. Among these, the assay targeting BPSS1187 exhibited the highest sensitivity (76.3%) and specificity (100%) using plasma samples [18].
Optimizing methods for detection of B. pseudomallei is crucial for accurately mapping endemic areas and guiding surveillance efforts that could help to enhance awareness and inform disease prevention strategies. To determine the most effective methods for detecting B. pseudomallei in environmental samples, we initially evaluated both culture- and real-time PCR-based methods using spiked soil samples. We compared the performance of ACER agar with Ashdown agar, and evaluated ACER broth, TBSS-C50-based erythritol broth, and TBSS-C50 broth for culturing B. pseudomallei from spiked soil samples. We also determined the efficiency of real-time PCR assays targeting BPSS1187 and TTS1-orf2 using these samples. We then examined the environmental prevalence of B. pseudomallei in 238 soil samples collected in Amnat Charoen province. These included soil surrounding the houses of individuals that were strongly positive (N = 6) and negative (N = 6) for Hcp1-specific antibodies. Amnat Charoen borders Ubon Ratchathani, a recognized melioidosis endemic area, but currently lacks serological surveillance data.
Materials and methods
Ethics statement
The study was approved by the Ethics Committee of the Faculty of Tropical Medicine, Mahidol University (MUTM 2024-052-01) and the Mahidol University Central Institutional Review Board (MU-CIRB 2023/028.1503). Written informed consent was obtained from all participants or their parents or guardians prior to their enrollment. This study was approved by the Institutional Biosafety Committee of Mahidol University (MU 2024–010).
Bacterial strains
B. pseudomallei isolates were obtained from both human and environmental sources in Northeast Thailand. The reference strain K96243 was isolated from a patient in Khon Kaen province [19]. Ten environmental isolates were obtained from soil samples across different locations in Surin Province, Thailand, including Khwao Sinarin (strains 30–191-S08, 30–191-S10, 30–191-S16, 30–191-S17), Chom Phra (strains 30–194-S03, 30–194-S04, 30–194-S14), and Sanom (strains 30–198-S22, 30–198-S23, and 30–198-S28). Bacterial manipulations were performed using biosafety level 3 (BSL-3) containment and handled according to institutional and national biosafety guidelines. Bacterial cultures were propagated in Luria-Bertani (LB) broth or on LB agar at 37°C unless otherwise specified. For long-term preservation, stocks were maintained in 20% glycerol at -80°C.
Comparison of Phosphate-buffered acidic erythritol (ACER) and Ashdown agar for culturing B. pseudomallei
The efficiency of different culture media for supporting growth of B. pseudomallei was initially evaluated by comparing two agar-based media: Ashdown agar [20] and ACER agar [12]. Bacterial growth was evaluated using ten environmental B. pseudomallei strains (30–191-S08, 30–191-S10, 30–191-S16, 30–191-S17, 30–194-S03, 30–194-S04, 30–194-S14, 30–198-S22, 30–198-S23, and 30–198-S28) and the reference clinical strain K96243. The bacterial strains were obtained from frozen stocks, sub-cultured on Columbia agar (Oxoid, UK), and incubated at 37°C for 24 hours. The resulting colonies were re-suspended in PBS, adjusted to an optical density 600 nm (OD_600_) of 0.18 (approximately 1 × 10⁸ CFU/ml), and diluted to a final concentration of 1 × 10^3^ CFU/ml. One hundred microliters of each of the suspensions was plated onto ACER agar and Ashdown agar in triplicate and incubated at 37°C for 3 and 7 days. The resulting colonies were enumerated, and their morphology was examined.
Comparison of B. pseudomallei recovered from spiked soil samples in different broth media
The efficiency of ACER, TBSS-C50-based erythritol, and TBSS-C50 media for the recovery of eleven different B. pseudomallei strains from spiked soil samples was evaluated as previously described [8,10,12]. Ten environmental strains (30–191-S08, 30–191-S10, 30–191-S16, 30–191-S17, 30–194-S03, 30–194-S04, 30–194-S14, 30–198-S22, 30–198-S23, and 30–198-S28) and the reference clinical strain K96243 were included in this study. Bacterial strains were sub-cultured on Columbia agar (Oxoid, UK) and incubated at 37°C for 24 hours. The resulting colonies were re-suspended in PBS, adjusted an OD_600_ to obtain approximately 1 × 10⁸ CFU/ml. One ml of each strain in PBS was spiked into 5 g of autoclaved soil, followed by the addition of 9 ml of PBS. The samples were then incubated at room temperature for 24 hours. After incubation, 500 µl of each of the soil supernatants was transferred into 10 ml of ACER, TBSS-C50-based erythritol, or TBSS-C50 enrichment broth, in duplicate, and then incubated under static conditions at 37°C with loosely capped tubes for 2, 5, and 9 days. At each time point, 100 µl of each broth culture was serially diluted (10-fold) in PBS, spread onto Ashdown agar in duplicate, incubated at 37°C for 4 days, following which bacterial colonies were enumerated.
Detection of B. pseudomallei from spiked samples using real time-PCR assays
DNA extraction.
For real-time PCR assays, genomic DNA was extracted from 200 µl of bacterial pellet obtained from 1 ml of B. pseudomallei K96243 culture in LB broth or 250 mg of autoclaved soil spiked with ten environmental B. pseudomallei strains using the PowerSoil Pro Kit (QIAGEN) following the manufacturer’s instructions. Briefly, the samples were lysed in Solution CD1, vortexed, and centrifuged. The resulting supernatants were treated with Solution CD2 for inhibitor removal, followed by DNA binding with Solution CD3 using an MB Spin Column. Columns were washed with Solutions EA and C5, centrifuged to remove residual wash buffer, and DNA was eluted with 50 µl of Solution C6. The purified DNA samples were stored at -80°C until needed for real-time PCR analysis.
Real-time PCR assay.
TaqMan real-time PCR assays were evaluated for the detection of B. pseudomallei using the TTS1-orf2 [15] and BPSS1187 [21] as the target genes. The assays were tested using purified genomic DNA from strain K96243 and the ten environmental isolates. The total reaction volumes were 10 µl and contained 5µl of 1X SensiFAST Probe No-ROX kit, 400 nM each of forward and reverse primers, 100 nM of the probe, and 1 µl of total genomic DNA. The amplification conditions for BPSS1187 included an initial denaturation step at 95°C for 5 minutes, followed by 45 cycles of 95°C for 15 seconds and 58°C for 30 seconds. For TTS1-orf2, the conditions consisted of an initial denaturation at 95°C for 8 minutes, followed by 45 cycles of 95°C for 15 seconds and 59°C for 15 seconds. All reactions were performed in a CFX96 Touch Real-time PCR system and analyzed using Bio-Rad CFX Maestro software, version 4.1.2433.1219 (Bio-Rad).
Data analysis was performed using serial 10-fold dilutions of genomic DNA, corresponding to a range of 10⁸ to 1 CFU/ml. Standard curves plotted threshold cycle (C_t_) values against the log of bacterial concentration (CFU/ml). Linear regression analysis was then applied to the data to assess the assay efficiency and dynamic range. PCR efficiency was calculated from the slope of the standard curve using the equation of E = 10^-1/slope^-1 [22].
Determination of the limit of detection of real-time PCR assays.
The limit of detection (LOD) of the real-time PCR assays was evaluated using specific primers targeting TTS1-orf2 and BPSS1187 in a TaqMan real-time PCR format [18]. The assays were performed using genomic DNA from B. pseudomallei K96243 in PBS and ten environmental strains spiked into soil samples. DNA was extracted as described above and concentrations were quantified using a NanoDrop Lite spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Genomic DNA corresponding to CFU/ml ranging from 10⁸ to 1 CFU/ml was analysed by real-time PCR. The LOD was defined as the lowest concentration at which amplification was consistently observed across three replicates and was expressed as CFU/ml.
Enzyme linked immunosorbent assay for detection of Hcp1-specific antibodies in plasma collected from healthy individuals
Hcp1-specific IgG antibodies were measured in 398 plasma samples collected from healthy individuals in a previous study conducted in Amnat Charoen province using a single point (1:2000 dilution) enzyme linked immunosorbent assay (ELISA), as previously described [23]. Two cut-off values were applied to define seropositivity. The first cut-off value calculated as mean OD_450nm_ plus 1.5 standard deviation (OD = 0.418) was used to identify individuals with probable exposure to B. pseudomallei. The second cut-off (OD 1.165), as previously described [24], was used to identify individuals with probable melioidosis.
Soil sampling
A total of 238 soil samples were collected from three districts in Amnat Charoen Province, Northeast Thailand in November 2024. Sampling locations were selected based on seroprevalence data from healthy populations, targeting households of healthy individuals who were strongly seropositive to B. pseudomallei (N = 6), as determined by Hcp1-specific ELISA, while households of seronegative individuals (N = 6) were randomly selected for soil sampling. The sampling sites included Mueang Amnat Charoen district (N = 109), Pathum Ratchawongsa district (N = 60), and Chanuman district (N = 69).
Soil sampling was performed as previously described [25]. Briefly, the soil samples were collected from a depth of 30 cm, with sampling points spaced 5–10 meters apart. A steel auger was used for soil collection and was cleaned with bottled water and disinfected with 70% ethanol between each sample to prevent cross-contamination. For each sampling point, 200 g of soil was placed in plastic zip lock bags, stored at ambient temperature, protected from direct sunlight, and transported to laboratory (Faculty of Tropical Medicine, Mahidol University, Bangkok) within two days.
Culture of B. pseudomallei from soil samples
For isolation of B. pseudomallei from soil samples*,* two culture approaches were performed using established protocols: 1) direct plating culture on Ashdown agar [25–27] and 2) culture in ACER enrichment broth [8,10,12,27]. For direct plating on agar, 100 g of soil from each sample was suspended in 100 ml of sterile water, mixed and incubated at room temperature overnight. Subsequently, two aliquots of 10 µl and 100 µl of the soil supernatants were plated onto Ashdown agar in duplicate and incubated at 42°C for 3 days. Bacterial colonies were then counted. Suspected B. pseudomallei colonies were confirmed using latex agglutination [28] and real-time PCR targeting TTS1-orf2 gene [15].
For culture in enrichment broth [12], five grams of soil from each sample was homogenized in 10 ml of PBS, and the suspensions were left to settle overnight at room temperature. A 500 µl aliquot of supernatant was then inoculated into 10 ml of ACER broth and incubated statically with a loose cap at 40°C for 5 and 9 days [12]. Following incubation, 100 µl of broth culture was 10-fold serial diluted, 100 µl of each dilution was plated onto Ashdown agar, and the plates were incubated at 40°C for 4 days. Bacterial colonies were counted and confirmed to be B. pseudomallei [15,28] as described above.
Real-time PCR of soil samples positive for B. pseudomallei by culture-based methods
A total of 20 soil samples that tested positive for B. pseudomallei by culture-based methods were extracted as described above. All samples were subsequently analyzed using real-time PCR assays targeting BPSS1187 and TTS1-orf2 genes.
Statistical analysis
Statistical analyses were performed using GraphPad Prism version 9.5.1 (GraphPad Software, San Diego, CA, USA). Comparisons of bacterial colony counts between different media were conducted using the Wilcoxon signed-rank test. The efficiencies of real-time PCR assays targeting the BPSS1187 and TTS1-orf2 genes were compared using a paired t-test. The association between the prevalence of B. pseudomallei and seroprevalence was assessed using the Fisher’s exact test. The Chi-square test was used to analyze the demographic distribution of individuals with Hcp1-specific antibody positivity.
Results
Comparison of Ashdown and ACER agar for culture of B. pseudomallei
The recovery of B. pseudomallei from PBS was assessed by culturing ten environmental strains and the clinical strain K96243 on Ashdown and ACER agar in three independent experiments (Fig 1). At day 3 post-inoculation, B. pseudomallei colonies were detectable on both media. For all experiments, the median CFU per plate on ACER agar was not significantly different from the median CFU per plate on Ashdown agar (Fig 1A and S1 Table). Colonies on Ashdown agar exhibited type I morphotypes [29], characterized by a rough central surface with irregular margins and a pale purple appearance, with a diameter ranging from 1 to 2 mm. In contrast, colonies on ACER agar appeared smooth, creamy, white, and opaque, with a diameter of approximately 0.1 to 0.5 mm (Figs 2 and S1).
Colony counts of eleven B. pseudomallei strains on Ashdown agar and ACER agar.The number of colonies of B. pseudomallei were counted at (A) 3 days and (B) 7 days of incubation at 37 °C. The experiments were performed in triplicate in three independent assays.
Colony morphology of B. pseudomallei on Ashdown agar and ACER agar.The colony morphology of B. pseudomallei 30-194-S04 was examined at 3 days and 7 days of incubation on Ashdown and ACER agar respectively.
At day 7 post-inoculation, the median CFU per plate on ACER agar was not significantly different from that of Ashdown agar. The number of colonies and morphologies were also the same as on day 3 (Fig 1B and S1 Table). In contrast, the average colony size (diameter) on Ashdown agar had increased to 5 mm but was only 1 mm on ACER agar Figs 2 and S1). Since the recovery of all strains of B. pseudomallei on Ashdown agar was similar and there was a more distinct colony morphology compared to ACER agar, Ashdown was selected for further experiments and used for environmental surveillance of B. pseudomallei.
Comparison of three different enrichment media for the recovery of B. pseudomallei from spiked soil samples
The recovery of ten environmental B. pseudomallei strains and the reference clinical strain K96243 from spiked soil samples was assessed using TBSS-C50, TBSS-C50-based erythritol, and ACER broth following 2, 5, and 9 days of incubation. Colony counts were determined by plating serial dilutions on Ashdown agar (Fig 3 and S2 Table). B. pseudomallei was recovered from all three types of broth at all time points, but ACER broth demonstrated a higher bacterial yield than TBSS-C50-based erythritol and TBSS-C50 broths. The median colony counts for ACER broth were significantly higher than those for TBSS-C50 broth (p = 0.001), while TBSS-C50-based erythritol broth showed the lowest recovery for all time points.
*Bacterial counts of eleven B. pseudomallei strains on TBSS-C50, TBSS-C50-based erythritol medium, and ACER broth.Eleven B. pseudomallei strains were grown statically at 37°C in TBSS-C50, TBSS-C50-based erythritol medium, and ACER broth for 2, 5, and 9 days. Each point represents one of the B. pseudomallei strains (**p < 0.001; Wilcoxon Signed-Rank Test).
Comparison of real-time PCR methods for detecting B. pseudomallei in PBS
We evaluated the performance of real-time PCR assays targeting BPSS1187 and TTS1-orf2 for detecting B. pseudomallei strain K96243 in PBS. Both targets exhibited strong linear correlations between log CFU/ml and C_t_ values. For BPSS1187, the average regression equation was y = –3.414x + 43.24 with an R^2^ of 0.9856 and a LOD of 10^2^ CFU/ml. For TTS1-orf2, the average regression equation was y = –5.447x + 56.39 with an R^2^ of 0.9734 and a LOD of 10⁴ CFU/ml (Fig 4 and S3 and S4 Tables). A paired-t test was used to compare PCR efficiencies across three replicates for *BPSS1187-*PCR (87.6%, 106.1%, and 96.9% efficiency) and TTS1-orf2-PCR (51.0%, 54.7%, and 51.5% efficiency). The analysis revealed significant differences in efficiency between the two assays, with a median efficiency of 96.9% for BPSS1187 and 51.5% for TTS1-orf2, representing a difference of 45.4% (p = 0.009).
Analytical sensitivity of real-time PCR assays for B. pseudomallei K96243.Calibration plots of bacterial concentration (log CFU/ml) versus cycle threshold (Ct) values for the BPSS1187 and TTS1-orf2 targets across nine serial dilutions (10⁸ to 10⁰ CFU/ml).
Comparison of real-time PCR methods for detecting B. pseudomallei in spiked soil samples
Next, we evaluated the performance of real-time PCR assays targeting BPSS1187 and TTS1-orf2 for detecting ten environmental B. pseudomallei strains spiked into soil samples. Calibration curves were generated for each strain and demonstrated high consistency across assays (Fig 5). For BPSS1187, the slopes ranged from –3.409 to –4.065, with R^2^ values between 0.9469 and 0.9977, and PCR efficiencies ranging from approximately 77% to 96%. For TTS1-orf2, the slopes ranged from –2.847 to –4.203, R^2^ values from 0.6819 to 0.999, and PCR efficiencies from approximately 77% to over 111%. To statistically compare the PCR efficiencies of the two targets amongst the 10 environmental strains, a paired t-test was performed. The analysis revealed no statistically significant differences in the efficiencies between the two targets with a median efficiency of 84.6% for BPSS1187 and 87.4% TTS1-orf2, which was a difference of only 2.8% (p = 0.188) (S4 and S5 Tables)
Analytical sensitivity of real-time PCR assays for 10 environmental B. pseudomallei strains.Calibration plots of bacterial concentration (log CFU/ml) versus cycle threshold (Ct) values for BPSS1187 and TTS1-orf2 across nine serial dilutions (10⁸ to 10⁰ CFU/ml).
Hcp1-specific antibody responses in healthy individuals in Amnat Charoen province
To investigate the environmental distribution of B. pseudomallei in relation to potential human exposures, we first determined the Hcp1-specific IgG responses in 398 plasma samples from healthy individuals living in seven different districts of Amnat Charoen province. Seropositivity was defined using two cut-off values OD_450_ = 0.418 (probable exposure to B. pseudomallei) and OD_450_ = 1.165 (probable melioidosis), established in a previous study [24]. Among the 398 samples, 34 individuals (8.5%) were identified as seropositive (OD ≥ 0.418). Of these, 28 individuals were classified as having probable exposure to B. pseudomallei, while 6 individuals were classified as having probable melioidosis (OD ≥ 1.165) (Fig 6 and Tables 1 and S6).
Table 1: Detection of Hcp1-specific IgG among healthy individuals from seven districts in Amnat Charoen Province, Thailand.
Distribution of antibody responses among 398 healthy individuals across seven districts of Amnat Charoen Province.(A) Scatter plot of antibody responses across seven districts of Amnat Charoen province. (B) Histograms showing the distribution of antibody responses within each district and across all districts combined. The solid black line represents the cut off value (0.418), while the dashed line marks the cut off value (1.165).
Seropositive individuals with probable melioidosis were identified in Chanuman (2/44, 4.5%), Mueang Amnat Charoen (3/136, 2.2%), and Pathum Ratchawongsa (1/48, 2.1%). While seropositive individuals with probable exposure to B. pseudomallei were more widely distributed and included Phana (4/31, 12.9%), Pathum Ratchawongsa (5/48, 10.4%), Senangkhanikhom (5/48, 10.4%), Lue Amnat (3/38, 7.9%), Hua Taphan (3/53, 5.6%), Chanuman (2/44, 4.5%), and Mueang Amnat Charoen (6/136, 4.4%). All 34 seropositive individuals were distributed across age groups, with a seropositivity rate of 7.5% among individuals aged 13–17 years (N = 53), 7.5% among those aged 18–59 years (N = 253), and 12.0% among those aged 60 years or older (N = 92). However, no statistically significant association was found between age group and seroprevalence (P = 0.410). By gender, seropositivity was 11.5% in males and 7.8% in females. No statistically significant association was observed between gender group and seroprevalence (P = 0.291). Differences by occupation were also observed, with seropositivity rates of 8.9% among students, 9.9% among farmers, and 6.3% among unemployed individuals. No statistically significant association was observed between occupation group and seroprevalence (*P *= 0.522) (Table 2).
Table 2: Demographic distribution of Hcp1-specific antibody positive individuals.
Soil sampling in areas surrounding the households of healthy individuals
Hcp1-ELISA results were used to guide the selection of soil sampling locations around the houses of individuals that were strongly positive (N = 6) and negative (N = 6) for Hcp1-specific antibodies. To investigate the prevalence of B. pseudomallei in soil samples, we employed two approaches: (I) direct plating on Ashdown agar and (II) enrichment culture in ACER broth prior to plating on Ashdown agar. The percentage of B. pseudomallei-positive samples varied across methods and districts. For direct plating on Ashdown agar, the highest detection rate was in Pathum Ratchawongsa, where 4/60 samples (6.7%) tested positive, followed by Mueang Amnat Charoen where 3/109 samples (2.8%) tested positive, and Chanuman where 1/69 samples (1.5%) tested positive. For ACER enrichment culture, positive results on day 5 were detected in 5/60 samples (8.3%) from Pathum Ratchawongsa, 6/109 samples (5.5%) from Mueang Amnat Charoen, and 1/69 sample (1.5%) from Chanuman. By day 9, the number of positive samples increased in Mueang Amnat Charoen (8 of 109; 7.3%), remained unchanged in Pathum Ratchawongsa and Chanuman (Fig 7 and Tables 3 and S7).
Table 3: Culture- and real-time PCR-based detection of B. pseudomallei in soil samples from three districts of Amnat Charoen Province, Thailand.
Geographical distribution of B. pseudomallei detected from soil samples in Amnat Charoen Province, Thailand.The main map highlights sampling sites within the province. Enlarged panels show culture results on Ashdown agar (top right) and ACER broth (bottom right) across three districts. Each pie chart represents the proportion of positive (red) and negative (green) samples, with size proportional to the number of samples collected (Mueang Amnat, n = 108; Pathum Ratchawongsa, n = 60; Chanuman, n = 69).
Of these 20 positive samples, 5 samples were positive in all culture methods. Two samples were positive only by direct plating on Ashdown agar, 4 samples were positive only after enrichment culture in ACER for 5 days, and 5 samples were positive only after enrichment culture in ACER for 9 days (Fig 8 and S8 Table).
Detection of B. pseudomallei in soil samples by culture and PCR methods.Euler diagram illustrating the overlap of results among culture-based and PCR-based detection methods for 20 B. pseudomallei-positive soil samples collected in Amnat Charoen Province, Thailand.
A total of 20 soil samples confirmed to be culture positive for B. pseudomallei were further analyzed using real-time PCR assays targeting the BPSS1187 and TTS1-orf2 genes. The results showed that 2 out of 11 samples from Mueang Amnat Charoen district, and none of the samples from Pathum Ratchawongsa (0/8) and Chanuman (0/1) districts, tested positive for both genes (Table 3).
The base map was generated using the open-source R packages rgdal and rworldmap, with administrative boundary shapefiles obtained from UN-OCHA (https://data.humdata.org/dataset/cod-ab-tha). The shapefile is freely available for scientific, educational, and commercial use under the Creative Commons Attribution 4.0 International License (CC BY 4.0) (https://creativecommons.org/licenses/by/4.0/).
The association between the presence of B. pseudomallei in the soil and seroprevalence
To evaluate the association between environmental detection of B. pseudomallei and seroprevalence, Fisher’s exact test was performed using data from 12 household soil samples, consisting of individuals that were strongly positive (N = 6) and negative (N = 6) for Hcp1-specific antibodies, as summarized in Table 4. Based on combined culture-based methods, B. pseudomallei was detected in three households associated with strongly seropositive individuals and in two households associated with seronegative individuals. The analysis revealed no significant association across the three districts (P = 0.165), indicating no statistically significant association between the presence of B. pseudomallei in the household environment and seroprevalence.
Table 4: Seroprevalence and culture-based detection of B. pseudomallei in household soil samples across three districts in Amnat Charoen Province, Thailand.
Discussion
Our evaluation of B. pseudomallei detection methods provides critical insights for optimizing environmental surveillance strategies in an endemic area of Northeast Thailand, through a comprehensive comparative analysis of culture- and molecular-based techniques on soil samples and the association between prevalence of B. pseudomallei and seroprevalence. Comparison of the growth of B. pseudomallei on Ashdown and ACER agar showed that Ashdown agar performed better than ACER agar in terms of growth and provided a more distinct colony morphology of B. pseudomallei. These findings are consistent with previous reports, which emphasized using Ashdown agar for presumptive identification, based on the distinct wrinkled and purple colony appearance characteristic of B. pseudomallei [29]. This particular colony appearance is attributed to components of Ashdown agar, including crystal violet, which contributes to the purple coloration of B. pseudomallei colonies, and neutral red, which enhances their distinctive wrinkled appearance [20]. Although ACER agar supported comparable bacterial counts, differentiation was challenging due to the less distinct colony form, necessitating confirmation tests.
Further comparison of the efficacy of TBSS-C50 broth, TBSS-C50-based erythritol broth, and ACER broth in culturing B. pseudomallei demonstrated that ACER broth provided superior enrichment performance, yielding a higher recovery rate of B. pseudomallei compared to both TBSS-C50-based erythritol broth and TBSS-C50 broth. This is consistent with previous studies reporting that ACER broth enhances the recovery of B. pseudomallei, likely due to its acidic condition (pH 6.3), which shortens the lag phase of bacterial growth [12]. While B. pseudomallei grows optimally at neutral pH, acidic conditions (approximately pH 5.0–5.5) have been shown to enhance recovery from environmental samples by suppressing competing organisms and reducing the lag phase [20,30]. Additionally, erythritol, used as a carbon source in enrichment media, has been demonstrated to promote B. pseudomallei proliferation and improve recovery rates from environmental samples [12,31].
Our study evaluated the efficiency of real-time PCR assays targeting BPSS1187 and TTS1-orf2 for the detection of B. pseudomallei. The PCR assay targeting BPSS1187 demonstrated significantly higher efficiency than the TTS1-orf2 assay when performed on PBS samples spiked with B. pseudomallei strain K96243. This result is consistent with a previous study that reported high sensitivity and specificity for B. pseudomallei detection in plasma samples using real-time PCR targeting BPSS1187 [18]. However, no statistically significant difference in PCR efficiency was observed between the two targets across multiple environmental strains. These findings suggest that both assays exhibited comparable performance for the detection of diverse B. pseudomallei strains in environmental samples.
We also investigated Hcp1-specific antibody responses to B. pseudomallei among individuals in Amnat Charoen province. Our study revealed an overall seropositivity rate of 8.5% among seropositive individuals that included individuals with probable exposure to B. pseudomallei (7.0%) and individuals with probable melioidosis (1.5%).This finding contrasts with a previous study conducted in Ubon Ratchathani, which reported a high seroprevalence rate of 38% among healthy individuals, based on indirect hemagglutination assay (IHA) at a cutoff titer of 1:80 [32]. However, the IHA has been shown in several studies to have low sensitivity and specificity [33,34], which may account for its higher background antibodies to non-specific antigens. In contrast, the Hcp1-ELISA has demonstrated greater sensitivity and specificity for detecting exposure to B. pseudomallei [24,34]. In terms of age distribution, our study found no significant differences in seropositivity among age groups. This contrasts with previous findings from northeast Thailand, where high antibody titers were detected in approximately 20% of 2,214 children aged <15 years in Udon Thani and 11% of children in Ubon Ratchathani, as determined using the IHA [35], potentially reflecting increased environmental exposure through outdoor school activities or contact with contaminated soil and water. Regarding gender, our findings also showed no significant difference in seropositivity. This contrasts with prior studies that have reported higher seroprevalence among males [36], attributed to differences in testing methods (IHA versus Hcp1-ELISA), behavioral factors such as outdoor occupations, greater exposure to soil and water, and alcohol consumption. Additionally, our results demonstrated no significant differences in seropositivity across occupational groups. This contrasts with previous studies that have reported significantly higher seroprevalence among farmers compared to non-farmers [32].
In addition, we collected soil from areas surrounding the households of individuals that were strongly positive and negative based on Hcp1-specific IgG levels to determine the prevalence of B. pseudomallei in the environment. Our study showed that enrichment culture in ACER broth generally outperformed direct plating which is consistent with previous studies demonstrating improved sensitivity of enrichment-based methods for isolation of B. pseudomallei in the environment [10,27,37]. Among the surveyed districts, Pathum Ratchawongsa exhibited the highest detection rate using both culture-based approaches. Notably, prolonged incubation in ACER broth (up to nine days) increased the number of positive samples in Mueang Amnat Charoen, highlighting the importance of extended enrichment, especially in samples with low bacterial load [12]. In contrast, Chanuman district showed the lowest detection rate across both methods. These findings suggest that the environmental prevalence of B. pseudomallei may be influenced by soil type, consistent with previous reports indicating that silt soils harbor a lower proportion of the bacterium compared to sandy or clay soils [38]. This aligns with our observation in Chanuman, where silt-dominant soils were predominant and relatively compact. In contrast, soil samples from Mueang Amnat Charoen and Pathum Ratchawongsa districts were primarily collected from agricultural areas such as rice paddies, which provide moist and nutrient-depleted conditions favorable for B. pseudomallei survival and proliferation [38,39]. Furthermore, seasonal variations influence the prevalence of B. pseudomallei. Previous studies have reported that the monsoon season is associated with the highest rates of B. pseudomallei detection compared to other seasons [40–43]. Since our study was conducted in November, during the dry season in Thailand, this timing may explain the relatively low detection rates observed, with B. pseudomallei identified in only 3.36% of samples by direct plating and 5.04%–5.46% using enrichment methods. Moreover, human activities might also play a role, with previous reports identifying agricultural work, particularly rice farming and gardening, as major risk factors due to the increased disturbance of soil environment [44–46].
We further tested the culture-positive soil samples using real-time PCR targeting the TTS1-orf2 and BPSS1187 genes. Of the 20 culture-positive soil samples, B. pseudomallei DNA was detected in two samples. This low detection rate may be attributed to the presence of PCR inhibitors commonly found in soil extracts, as well as low DNA yield, since PCR was performed directly on soil without prior enrichment or the use of soil pellet for DNA extraction. In contrast, previous studies have successfully detected B. pseudomallei DNA from soil samples by conducting PCR after enrichment using TBSS-C50 based erythritol broth, or ACER broth [10,12] or by extracting DNA from direct soil samples [47,48]. This discrepancy may reflect differences in DNA extraction efficiency, the effectiveness of inhibitor removal, soil composition, and the smaller quantities of soil used for DNA extraction, which may reduce the likelihood of detecting B. pseudomallei given its heterogeneous distribution in soil.
Understanding the association between the environmental prevalence of B. pseudomallei and human seroprevalence is crucial for informing surveillance strategies in endemic areas. In this study, we investigated whether the presence of B. pseudomallei in household soil samples was associated with the individual seroprevalence. However, the results revealed no statistically significant association between environmental detection of B. pseudomallei and seroprevalence, and several factors may contribute to the absence of a strong correlation. Seroprevalence reflects cumulative or past exposure, whereas culture-based methods detect the current and often localized presence of B. pseudomallei in the environment. The intermittent distribution of B. pseudomallei [49], combined with individual variation in exposure behavior, such as farming or gardening activities that involve contact with contaminated soil and water [50,51], may contribute to this discrepancy. Furthermore, host immune responses to B. pseudomallei are highly variable and can influence the detectability of antibody responses, with some individuals mounting strong cell-mediated immunity in the absence of measurable antibodies [52]. In addition, environmental sampling limitations, such as low sampling density or non-representative site selection may lead to underestimation of B. pseudomallei prevalence in soil [53]. These factors may contribute to the discordance between the environmental prevalence of B. pseudomallei and human seroprevalence.
This study has several limitations. First, our evaluation of culture-based methods was conducted using only a limited number of B. pseudomallei strains. Future studies should include a broader range of soil-dwelling bacteria to better assess the selectivity and overall microbial diversity influenced by each culture condition. In addition, the media optimization experiments were performed with sterile soil, which may not accurately reflect the microbial competition present in natural environments. Future studies should utilize non-sterile soils during media optimization to better reflect natural ecological conditions. Second, the real-time PCR assays in our study were performed using direct soil samples, which may have reduced detection sensitivity due to the presence of PCR inhibitors or low DNA yield. In addition, the relatively small amount of soil used for DNA extraction (approximately 250 mg) may have further limited the sensitivity of real-time PCR, as B. pseudomallei is unevenly distributed in soil. Consequently, randomly selected sub-samples may not have contained sufficient bacterial cells for detection, thereby reducing assay sensitivity. This limitation could have affected the ability to detect B. pseudomallei in actual soil samples. Future studies should consider using prior enrichment or soil pellet for DNA extraction and incorporate validated internal controls to enhance the accuracy and reliability of real-time PCR assays. Finally, at some sampling locations and during certain periods, the soil was compacted or difficult to dig, which posed challenges for sample collection and may have limited the number of samples obtained. Therefore, sampling should ideally be conducted during the monsoon season, when soil conditions are more favorable. Addressing these limitations in future studies will improve the accuracy and applicability of environmental surveillance for B. pseudomallei.
Overall, this study highlights the effectiveness of culture-based and molecular methods for detecting B. pseudomallei from environmental soil samples. Our findings revealed a low prevalence of B. pseudomallei in Amnat Charoen Province. Moreover, this study provides additional insight into the environmental distribution and detection challenges of B. pseudomallei, and supports the integration of combined culture-based, molecular, and serological approaches for effective surveillance and development of public health responses for melioidosis.
Supporting information
S1 TableColony counts of eleven B. pseudomallei strains on Ashdown and ACER agar.The colony forming unit (CFU/ml) of eleven B. pseudomallei was measured at 3 and 7 days of incubation at 37 °C. The experiments were performed in triplicate in three independent assays.(DOCX)
S1 FigColony morphology of eleven B. pseudomallei strains on Ashdown and ACER agar.The growth of B. pseudomallei was measured at 3 and 7 days of incubation at 37 °C in air. The experiments were performed in triplicate in three independent assays.(PDF)
S2 TableColony counts of eleven B. pseudomallei strains on TBSS-C50, TBSS-C50 based erythritol and ACER enrichment broth.The log colony forming unit (log CFU/ml) of eleven B. pseudomallei was measured at 2, 5 and 9 days of incubation at 37 °C. The experiments were performed in triplicate in three independent assays.(DOCX)
S3 TableLimit of detection (LOD) of real-time PCR targeting BPSS1187 and TTS1-orf2 genes in B. pseudomallei K96243-spiked PBS.The table shows the limit of detection at nine serial dilutions (10^8^-10^0^ CFU/ml).(DOCX)
S4 TableLinear regression analysis for real-time PCR targeting BPSS1187 and TTS1-orf2 genes.The table shows the regression equations, correlation coefficients (R^2^ values), and amplification efficiencies for each gene target.(DOCX)
S5 TableLimit of detection (LOD) of real-time PCR targeting BPSS1187 and TTS1-orf2 genes in spiked soil with ten environmental B. pseudomallei strains.The table shows the limit of detection at nine serial dilutions (10^8^-10^0^ CFU/ml).(DOCX)
S6 TableSeroprevalence data of individuals in Amnat Charoen Province.The table presents seroprevalence data, with the first cut-off value (0.418) highlighted in bold purple and the second cut-off value (1.165) highlighted in bold red.(DOCX)
S7 TableDetection of B. pseudomallei in soil samples using culture methods.The table presents the detection results obtained from direct culture on Ashdown agar and enrichment culture in ACER broth at 5 and 9 days of incubation.(DOCX)
S8 TableSummary of B. pseudomallei-positive soil samples detected by culture and PCR methods in Amnat Charoen province.Red color indicates samples that tested positive by each corresponding method.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Oslan SNH, Yusoff AH, Mazlan M, Lim SJ, Khoo JJ, Oslan SN, et al. Comprehensive approaches for the detection of Burkholderia pseudomallei and diagnosis of melioidosis in human and environmental samples. Microb Pathog. 2022;169:105637. doi: 10.1016/j.micpath.2022.105637 35710088 · doi ↗ · pubmed ↗
- 2Vuddhakul V, Tharavichitkul P, Na-Ngam N, Jitsurong S, Kunthawa B, Noimay P, et al. Epidemiology of Burkholderia pseudomallei in Thailand. Am J Trop Med Hyg. 1999;60(3):458–61. doi: 10.4269/ajtmh.1999.60.458 10466977 · doi ↗ · pubmed ↗
- 3Wiersinga WJ, Currie BJ, Peacock SJ. Melioidosis. New England Journal of Medicine. 2012;367(11):1035–44.10.1056/NEJ Mra 120469922970946 · doi ↗ · pubmed ↗
- 4Limmathurotsakul D, Golding N, Dance DAB, Messina JP, Pigott DM, Moyes CL, et al. Predicted global distribution of Burkholderia pseudomallei and burden of melioidosis. Nat Microbiol. 2016;1:15008. doi: 10.1038/nmicrobiol.2015.8 27571754 · doi ↗ · pubmed ↗
- 5Chantratita N, Phunpang R, Yarasai A, Dulsuk A, Yimthin T, Onofrey LA, et al. Characteristics and One Year Outcomes of Melioidosis Patients in Northeastern Thailand: A Prospective, Multicenter Cohort Study. Lancet Reg Health Southeast Asia. 2023;9:100118. doi: 10.1016/j.lansea.2022.100118 36570973 PMC 9788505 · doi ↗ · pubmed ↗
- 6Stewart T, Engelthaler DM, Blaney DD, Tuanyok A, Wangsness E, Smith TL, et al. Epidemiology and investigation of melioidosis, Southern Arizona. Emerg Infect Dis. 2011;17(7):1286–8. doi: 10.3201/eid 1707.100661 21762589 PMC 3381374 · doi ↗ · pubmed ↗
- 7Wiersinga WJ, van der Poll T, White NJ, Day NP, Peacock SJ. Melioidosis: insights into the pathogenicity of Burkholderia pseudomallei. Nat Rev Microbiol. 2006;4(4):272–82. doi: 10.1038/nrmicro 1385 16541135 · doi ↗ · pubmed ↗
- 8Limmathurotsakul D, Dance DAB, Wuthiekanun V, Kaestli M, Mayo M, Warner J, et al. Systematic review and consensus guidelines for environmental sampling of Burkholderia pseudomallei. P Lo S Negl Trop Dis. 2013;7(3):e 2105. doi: 10.1371/journal.pntd.0002105 23556010 PMC 3605150 · doi ↗ · pubmed ↗