Molecular genetic characterization of CASEIN KINASE 1-LIKE 12 in Arabidopsis
Adam Seluzicki, Annemarie E. Branks, Sowmya Poosapati, Joanne Chory

TL;DR
This study explores the role of the CASEIN KINASE 1-LIKE 12 gene in Arabidopsis and its connection to plant growth and hormone signaling.
Contribution
The study identifies CKL12 as a potential downstream target of brassinosteroid signaling in Arabidopsis.
Findings
CKL12 may regulate hypocotyl and root growth in Arabidopsis seedlings.
The 3′ end of the CKL12 transcript appears important for its function.
CKL12 is a transcriptional target of brassinosteroid signaling.
Abstract
The CASEIN KINASE 1 (CK1) family plays diverse roles in the development, physiology, and disease in eukaryotes. In Arabidopsis thaliana, the CASEIN KINASE 1-LIKE (CKL) family has 13 members, but to date, the roles of these kinases remain largely unclear. Here, we characterize several insertion mutants, finding that CKL12 may contribute to hypocotyl and primary root growth. Differential effects of insertions at various parts of the gene suggest that the 3′ end of the transcript may be important for CKL12 function. We provide evidence that CKL12 may be a transcriptional target of brassinosteroid (BR) signaling. The CKL12 promoter contains in vitro binding sites for BR-related transcription factors. Knock-down of these transcription factors using RNA interference reduces CKL12 transcript. Together, these data suggest that CKL12 may act downstream of BR signaling to regulate seedling growth.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
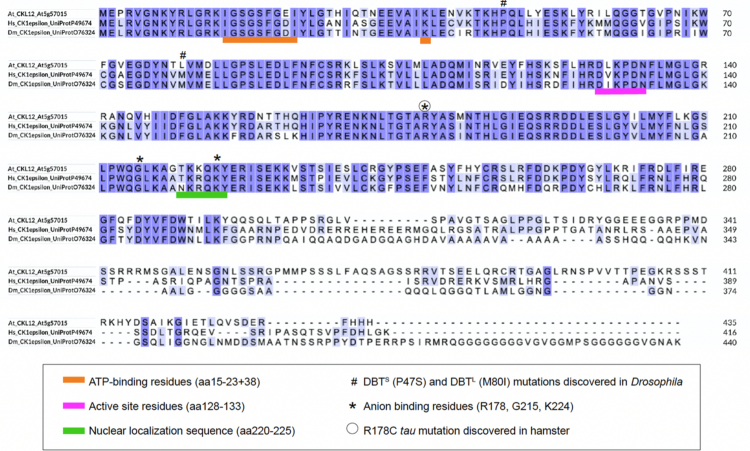 Figure 1
Figure 1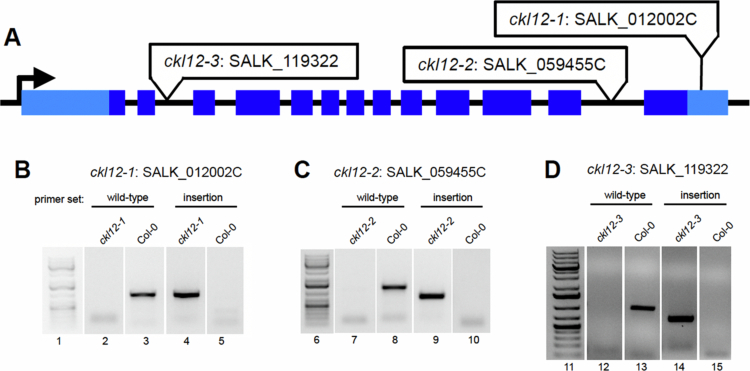 Figure 2
Figure 2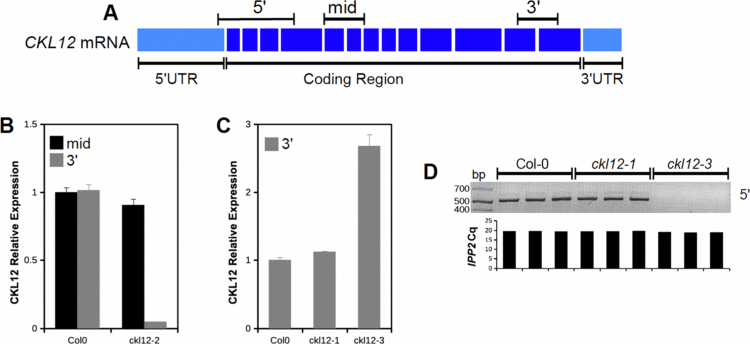 Figure 3
Figure 3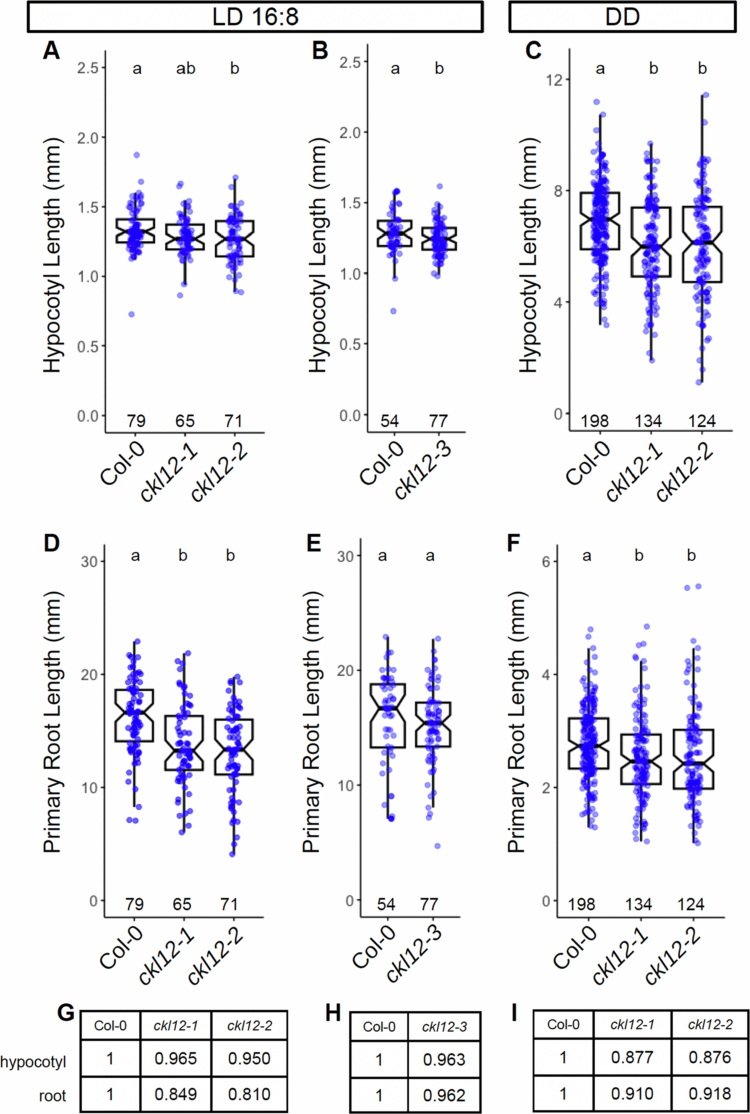 Figure 4
Figure 4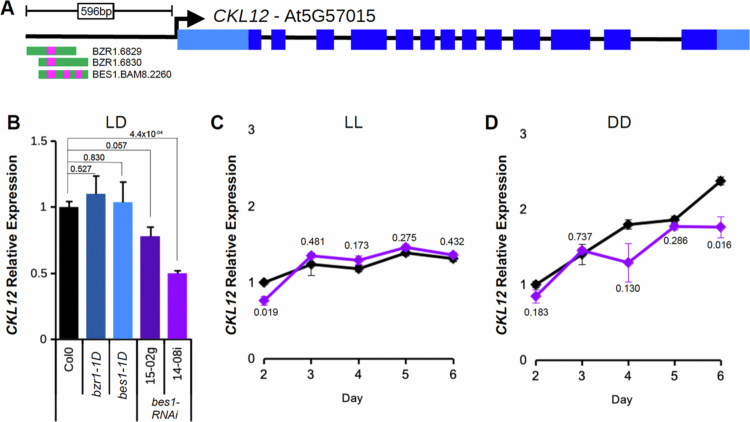 Figure 5
Figure 5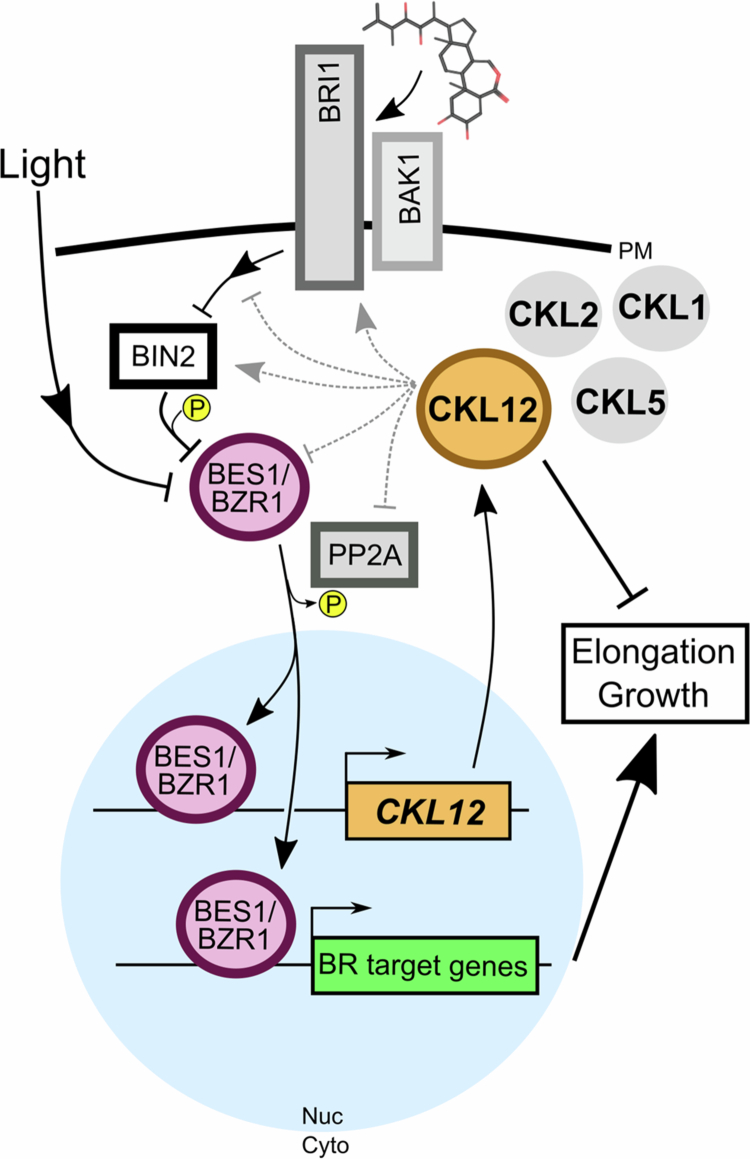 Figure 6
Figure 6- —Howard Hughes Medical Institute10.13039/100000011
- —National Institutes of Health10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant nutrient uptake and metabolism · Plant Gene Expression Analysis
Introduction
The CASEIN KINASE 1 (CK1) family of serine/threonine kinases is well described in eukaryotes, playing central signaling roles in controlling the circadian clock and regulating cellular signaling in development.1-4 CK1 proteins have highly conserved compact kinase domains, followed by highly variable C-terminal regulatory domains with autoinhibitory properties.5-7 The Arabidopsis genome contains 13 orthologs of CK1, with closest homology to the CK1δ/ε group described in animals. The Arabidopsis CKL family contains three subgroups (CKL-A, -B, and -C), and it appears that several members have undergone duplications.8 Four additional genes (MUT9-LIKE KINASES (MLKs) or PHOTOREGULATORY PROTEIN KINASES (PPKs)), show shared homology with the CK1 kinase domain but have larger C-terminal extensions.9
Despite the clear importance of CK1 kinases in many systems, there are relatively few studies examining them in plants. To date, most members of the CKL family remain uncharacterized. Pharmacological studies have shown that broad inhibition of CASEIN KINASE 1-LIKE (CKL) proteins in Arabidopsis results in altered progression of the circadian clock, possibly via through effects on PRR5 and TOC1.8-10 Similar results were obtained with broad knock-down of the CKL family using RNA interference, although no single CKL was found to be uniquely important for these effects.10 CKL3 and CKL4 were implicated in control of blue light signaling via direct phosphorylation of CRYPTOCHROME 2, although a later study mapped this activity to the PPK subgroup.11^,^12 CKL6 was shown to associate with microtubules through the C-terminal domain, directly phosphorylate TUBULIN beta 3, and localize to late endosomal vesicles.13^,^14 CKL2 was described as a positive regulator of Abscisic Acid (ABA) responses, including germination, root growth, and proline accumulation.15 CKL2 was also found to be a factor regulating crosstalk between the ABA and Brassinosteroid (BR) hormone signaling pathways, as it is activated by ABA during stress to directly phosphorylate the BR receptor BRI1, priming it for rapid recovery after stress has passed.16 There is little information regarding CKL12 in Arabidopsis. One study generated promoter-reporter constructs to examine the expression patterns of the CKL family in different plant tissues, finding that the CKL12 promoter could drive expression in the vasculature, trichomes, and anthers, as well as in the root cap, root tip, and primary root.17
Brassinosteroid hormone signaling is a central pathway controlling plant growth. Loss of function in this pathway results in dwarf plants that show a photomorphogenic phenotype when grown in the dark. The hypocotyl fails to elongate, and the apical hook and cotyledons open similar to those of light-grown plants.18-23 Genetic screens for suppressors of these phenotypes uncovered dominant mutations in the BR-responsive transcription factors Brassinazole Resistant 1 (BZR1) and BRI1 EMS Suppressor 1 (BES1).24^,^25 The bzr1-1D and bes1-1D mutants both mimic mild BR deficiency in long-day (LD) conditions and rescue BR deficiency in constant dark (DD). The search for additional homologs of these TFs revealed a small family of BES1/BZR1 HOMOLOG (BEH) genes that also contribute to transcription downstream of BR signaling.26 The downstream transcription factors in the BR pathway also have feedback effects on BR hormone synthesis and interact with other key signaling pathways, including auxin and gibberellin, as well as light signaling.25^,^27-31
Here, we identify genetic reagents, characterize seedling growth, and analyze transcriptional regulation of CASEIN KINASE 1-LIKE 12 (CKL12). We find that insertion mutations disrupting the 3′ end of the gene may cause mild reductions in growth in both the hypocotyl and primary root, while a possible null mutant has weaker effects. We also provide evidence that BR-related transcription factors are likely to play a role in promoting CKL12 transcription. Together, this study provides initial characterization of a member of the underexplored CKL gene family in Arabidopsis.
Materials and methods
Plant material
Arabidopsis thaliana plants were propagated on soil with added fertilizer and fungicide. Insertion mutants for CKL12 (AT5G57015) were obtained from the Arabidopsis Biological Resource Center (ABRC – The Ohio State University). Insertion lines were confirmed by PCR from genomic DNA using primers in Supplementary Table 1. Lines carrying bzr1-1D and bes1-1D were described.24^,^25 RNAi lines targeting BES1/BZR1 were described.26
Growth conditions
Seeds were surface-sterilized using chlorine gas and plated on 1/2× Linsmaier & Skoog medium + 0.8% phytoagar (Caisson). The plates were stratified in the dark at 4 °C for 4 d. The plates were then moved to growth chambers (Percival) under appropriate conditions: LD (16 h light-8h dark, light intensity: 100 μmol m^−2^s^−1^, 20 °C), LL (constant light, 100 μmol m^−2^s^−1^, 20 °C), and DD (constant darkness, 20 °C). The light source was cool-white fluorescent bulbs (Philips). Seeds to be grown under DD were exposed to white light to trigger germination, then wrapped in foil and grown in the same chamber as LD or LL plates. The plates were scanned and measured using ImageJ software (NIH).32
qRT-PCR
RNA was extracted from seedlings collected at the indicated time points using the RNeasy Plant Mini Kit (Qiagen) and treated with DNAse according to the manufacturer's instructions. Maxima First Strand cDNA Synthesis Kit was used to make cDNA from 2 μg of RNA from each sample. Reactions were run using SYBR-Green detection in a Bio-Rad CFX Opus 384 Real-Time PCR Machine. IPP2 (AT3G02780) transcript was used as the internal control for quantification using the ΔΔCq method. Semi-quantitative RT-PCR was done using samples previously used for qRT-PCR, run for 40 PCR cycles (annealing: 56 °C for 30 s, elongation: 72 °C for 45 s) and run on 2% agarose gel with ethidium bromide visualization.
Statistics
Student's t test (two-tailed) was used for comparison between two samples. One-way ANOVA + Tukey's HSD test (α = 0.05) was used to compare three or more samples.
Results
Protein sequence alignment of CKL12 with human and fly CK1ε
Examination of the protein sequence of CKL12 in relation to the human and Drosophila CK1ε shows that the key residues in the function of the kinase domain are conserved across evolutionary time. The amino acid residues that form the ATP-binding pocket, the active site, and the location of mutations identified in mammals (human, hamster) and in Drosophila are indicated on the alignment.33^,^34 The ATP-binding pocket (orange) and active site residues (magenta) of CKL12 are identical to those of HsCK1 and DmCK1 (Figure 1). Proline 47, which is mutated to serine in the Drosophila dbt^S^ mutant, is conserved. However, methionine 80, which is mutated to isoleucine in dbt^L^, is a leucine in CKL12. This raises the possibility that the kinase activity of CKL12 may be reduced relative to that of other family members.35 All three residues that form the anion-binding motif, one of which is notable for being mutated in the tau mutant hamster (R178C), are retained in CKL12.34 The C-terminal regions of all three proteins are of similar length but contain unique sequence.
Protein sequence alignment of CKL12 with human and Drosophila CK1ε. Sequence alignment is annotated with features identified in mammalian and Drosophila CK1. The features include the ATP-binding domain (orange underscore); active site (magenta underscore); nuclear localization sequence (green underscore); amino acids altered in Drosophila DBTS and DBTL mutants (#); and anion-binding site residues (). One of the anion-binding residues, R178, is mutated in the tau mutant hamster and is indicated by a circled asterisk.*
Isolation and molecular characterization of T-DNA insertion alleles of CKL12
In the effort to expand the set of reagents available to study the CKL family in Arabidopsis, we isolated and confirmed three insertion alleles of CKL12. These three insertions are in three distinct regions of the gene, with different expected consequences. SALK_012002 (referred to from here on as ckl12-1) is inserted in the 3′ untranslated region (UTR), SALK_059455 (ckl12-2) is inserted in the 12th intron, and SALK_119322 (ckl12-3) is inserted in the second intron (Figure 2A). The insertion site and zygosity of each of these insertions were confirmed by PCR, and the lines were propagated from homozygous individuals (Figure 2B–D).
Isolation and confirmation of TDNA insertion mutants in CKL12. (A) Structure of the CKL12 gene locus. 5′ and 3′ untranslated regions are light blue rectangles. Exons are in dark blue rectangles. The insertion sites for each allele are indicated in white boxes. (B)–(D) PCR-amplified products from genomic DNA isolated from wild-type or mutant plants. Primer sets targeting wild-type genomic DNA flank the TDNA insertion site. Primers targeting the TDNA alleles use one primer within the TDNA and one of the primers used for wild-type amplification. The numbers below the gel images indicate the region of the unspliced images marked in the Supplemental Figure 1.
To examine the molecular consequences of each insertion on the production of CKL12 transcript, we carried out quantitative reverse-transcription polymerase chain reaction (qRT-PCR) assays. The primer sets used are diagrammed in Figure 3A. Analysis of ckl12-2, which was inserted in intron 12, showed wild-type transcript levels when assayed with primers across exons 5–6 but failed to amplify across exons 12–13, suggesting that the T-DNA insertion disrupts transcription or splicing of the 3′ end of the coding region (Figure 3B). CKL12 transcript level assayed with the “mid” primer set was similar to wild-type in ckl12-1 but was significantly elevated in ckl12-3 (Figure 3C). Analysis of the same cDNA from Figure 3C using the 5′ primer set, which surrounds the ckl12-3 insertion site, amplified from the Col-0 and ckl12-1 samples but did not amplify from ckl12-3 samples (Figure 3D). Thus, ckl12-3 could potentially result in the overexpression of a CKL12 fragment that lacks the first 25 amino acids encoding the ATP-binding pocket (Figure 1). Together, these data indicate that ckl12-3 is a likely loss-of-function allele, while ckl12-1 and ckl12-2 may modify CKL12 protein function in more complex ways.
Analysis of CKL12 transcript in TDNA mutant plants. (A) Model of mature CKL12 mRNA. The target amplicons are indicated above the model. (B, C) qRT-PCR of CKL12 transcript. Transcript abundance was quantified using the ΔΔCq method, with IPP2as the internal control. The experiments were done in biological triplicate. (B) CKL12 transcript in ckl12-2 using the mid (black bars) and 3′ (gray bars) amplicons sampled on day 6. (C) qRT-PCR of ckl12-1 and ckl12-3 mutants using the 3′ (gray bars) amplicon sampled on day 4. (D) Semi-quantitative RT-PCR of CKL12 transcript using the 5′ amplicon in the same cDNA samples as (C). Cq values for IPP2 in each replicate from (C) are aligned to each sample beneath the gel image.
CKL12 may promote growth in the hypocotyl and root
To begin to understand the function of CKL12, we tested seedling growth in ckl12 mutants. Under a long-day photoperiod (LD16:8) at 20 °C, ckl12-2 and ckl12-3 showed slight but statistically shorter hypocotyls than control, while ckl12-1 was comparable to control, all of which were ~95% of Col-0 (Figure 4A,B,G,H). Dark (DD) grown ckl12-1 and ckl12-2 seedling hypocotyls were shorter than those of the control, showing ~10% reductions in both hypocotyl and primary root lengths (Figure 4C,I). Primary roots were shorter than control in ckl12-1 and ckl12-2 (Figure 4D,G), but comparable to control in ckl12-3 (Figure 4E,H). The short-root phenotypes of ckl12-1 and ckl12-2 was maintained in DD (Figure 4F,I). Together, these data suggest that disruption, but not elimination, of CKL12 may disrupt growth in both hypocotyl and root.
Hypocotyl and root growth in ckl12 mutants. (A)–(C) Hypocotyl length. (D)–(F) primary root length. (A,B,D,E) seedlings grown under LD16:8, measured on day 6. (C, F) Seedlings grown in constant dark (DD), measured on day 3. Box plots show pooled data from independent replicate experiments [(A, D) 3, (B, E) 2, (C, F) 5]. One experiment included ckl12-1, ckl12-2, and ckl12-3. Col-0 data from this experiment are included in both (A, B) and (D, E) for hypocotyl and root measurements, respectively. Mean relative hypocotyl and primary root lengths in LD (G, H) and DD (I), normalized to Col-0 = 1, are noted in the tables. Box plots indicate the median and 25th and 75th percentiles. Whiskers extend to 1.5 * interquartile range. Notches approximate the 95% confidence interval of the median. N (individual plants scored) is shown near the x-axis for each genotype. Different letters at the top indicate statistically significant difference between groups (α = 0.05) by one-way ANOVA + Tukey HSD.
CKL12 expression is regulated by brassinosteroid-related transcription factors
We hypothesized that transcriptional regulation of CKL12 may provide clues as to its function. We searched the DAP-seq database for transcription factor binding sites in the promoter region of CKL12, comprised of ~600 bp upstream of the transcription start site to the nearest neighboring gene. We found annotated binding sites for BZR1 and BES1, two key transcription factors of the brassinosteroid (BR) signaling pathway (Figure 5A).36 We hypothesized that the BR pathway may regulate CKL12 expression. We assayed CKL12 transcript in several lines that have modified BR TFs in 10-day-old seedlings grown in LD conditions. The bzr1-1D and bes1-1D mutants both expressed CKL12 at levels comparable to those of the control. Two bes1-RNAi lines, which knock down both BES1 and BZR1, reduce CKL12 transcript (Figure 5B).26 The stronger of the two lines, 14-08i, reduces CKL12 expression by approximately half.
CKL12 transcription may be promoted by brassinosteroid-related transcription factors in darkness. (A) Diagram derived from the DAP-seq browser track for the CKL12 locus. CKL12 gene structure is shown at the top. The CKL12 promoter (from the CKL12 transcription start site up the 3′ end of the upstream gene transcript) is 596bp. Transcription factor binding sites are shown below the gene model, with BES1 and BZR1 binding sites noted with magenta bars. (B) qRT-PCR of CKL12 in 10-day-old seedlings grown in LD16:8. Genotypes: bzr1-1D (dark blue), bes1-1D (light blue), and bes1-RNAi lines 15-02g (dark purple) and 14-08i (light purple). Transcript abundance was determined using the ΔΔCq method, using IPP2as the internal control in biological triplicate. Error bars are SEM. Two-tailed t test p values for each mutant vs. Col-0 are indicated above the bars. (C, D) Time course expression of CKL12 in Col-0 (black) and bes1-RNAi 14-08i (light purple) on days 2–6 grown in LL (C) and DD (D). Expression level was determined using the ΔΔCq method using IPP2 as the internal control, which was normalized to that of Col-0 on day 2, within each light condition. The samples in (B)–(D) were assayed using the 3′ primer set diagramed in Figure 3. Samples are in biological triplicate with the exception of bes1-RNAi in DD on day 4, which is in duplicate. The error bars are SEM. Two-tailed t test p values for bes1-RNAi vs. Col-0 are included near the traces.
BR signaling is required for hypocotyl growth in the dark. We observed reduced hypocotyl elongation in the dark in ckl12 mutants. Thus, we examined CKL12 transcript in constant light (LL) and constant dark (DD) conditions in Col-0 and bes1-RNAi plants across days 2–6. In LL, CKL12 transcript shows a very mild increase from day 2 to day 6, and expression is similar in Col-0 and bes1-RNAi (14-08i) plants, with the exception of lower expression on day 2 in the RNAi line (Figure 5C). In DD, CKL12 transcript increases more rapidly, more than doubling from day 2 to day 6 in Col0 (Figure 5D). The bes1-RNAi plants show attenuated CKL12 transcript accumulation, with the strongest effect on day six. Together, these data support the hypothesis that BR-related transcription factors may be important transcriptional regulators of CKL12-expression.
Discussion
CK1 proteins are important regulators of many signaling processes in eukaryotes, but are little studied in plants. In this study, we have provided genetic and molecular characterization of CKL12, a rarely studied member of the CASEIN KINASE 1-LIKE family in Arabidopsis. We have isolated and confirmed three insertion mutant alleles, and characterized the consequences of these insertions on gene expression (Figures 2 and 3). There are other insertions in the CKL12 genomic locus, with most mapping to the last intron near the site of ckl12-2. It is unclear why this is such a “hot spot” for insertions, but it is possible that there are sequence-intrinsic properties that enhance integration. Indeed, T-DNA insertions are known to preferentially integrate in regions of low GC content, and the last intron of CKL12 is only 29% GC.37 We isolated ckl12-3, which is inserted in the second intron and strongly disrupts the CKL12 transcript (Figures 2 and 3). Interestingly, this allele shows the strongest disruption, and the weakest phenotypes of the alleles we tested (Figure 4G–I). The pROK2 vector, which comprises the insertion vector in ckl12-3, contains a 35S promoter, which is likely the cause of the high expression detected 3′ of the insertion.38 However, the lack of contiguous transcript in the 5′ end of the gene, connecting the regions encoding the ATP-binding pocket and the catalytic site, suggests that this allele would be unable to produce functional protein (Figure 3D). These results support the hypothesis that CKL12 is not an essential gene and that the uneven distribution of insertion sites is more likely due to sequence preference than to negative consequences of disrupting the gene.
Two of the three alleles that we characterized have insertions in the 3′ end of the gene: ckl12-1 in the 3′ UTR, and ckl12-2 in intron 12 (Figure 2A‒C). Both insertions had minimal effects on transcript levels (Figure 3B and C). However, ckl12-1 is an insertion in the 3′ UTR, potentially altering the translation, localization, or binding of accessory proteins.39 Additionally, ckl12-2 disrupted the 3′ end of the transcript, potentially removing the 72 C-terminal amino acids (Figures 1 and 3). These two alleles showed similar hypocotyl and root phenotypes under the conditions tested. The C-terminal tails of CK1 proteins are often autoinhibitory. It is possible then, that removing a portion of this tail, as in ckl12-2, may reduce autoinhibition and lead to a more active kinase. Similarly, modification of the 3′ UTR, as in ckl12-1, may lead to enhanced, or unregulated, translation. Thus, we may be observing the phenotypic consequences of hyperactivity or overproduction of CKL12 in these mutants. This interpretation leads to the hypothesis that ckl12-1 and ckl12-2 act as hypermorphic alleles and that CKL12 might act as an inhibitor of growth in the wild-type conditions. Given that the C-terminal extensions of CK1 family proteins often act in an autoinhibitory or autoregulatory capacity, it is possible that these mutants represent the consequences of removing this regulatory mechanism. However, more detailed experiments are necessary to determine the contribution of the C-terminal tail to the regulation of CKL12 activity.
The CKL family in Arabidopsis contains 13 members, and sequence analysis has suggested that this family has expanded via the whole genome or other duplications in the recent past.17 It is likely that these genes retain redundant functions. We find that the three alleles we tested show mild phenotypes, with the strongest two alleles showing ~10% reductions in hypocotyl and root length (Figure 4). If, as discussed above, the strongest phenotypes arise from overactive CKL12, with minimal consequences in the expected loss-of-function mutant, deeper analysis of CKL functions will likely require both overexpression constructs, including truncations removing the C-terminal tails, and higher-order mutants. CKL12 fits into a clade with CKL1, 2, and 5, with CKL1 being its closest paralog.17 It may therefore be necessary to generate double, triple, or quadruple loss-of-function mutants covering all members of this clade to fully examine the roles of CKL genes in Arabidopsis.
Environmental light is a critical regulator of plant growth, particularly in early seedlings. In Arabidopsis, seedlings grown under constant light show short hypocotyls and long roots, while seedlings grown in constant dark show long hypocotyls and shorter roots.40 The length of the photoperiod in light‒dark cycles determines the growth pattern, with longer and shorter days leading to shorter and longer hypocotyls, respectively.41 These growth responses are dependent on both auxin and brassinosteroid hormones.42^,^43 Indeed, genetic screens looking for mutants that resembled light-grown plants when grown in the dark were key to the discovery of many critical components in BR synthesis, signaling, and transcriptional control.18-20^,^22-25 Light inhibits BR signaling by destabilizing BZR1 and BES1 through multiple mechanisms and linking active BR signaling to low light, shade, and dark conditions.44 We find that ckl12-1 and ckl12-2 show slight defects in hypocotyl elongation in DD, but not LD light conditions (Figure 4). Additionally, we observe little difference in CKL12 transcript levels between Col0 and BES1-RNAi in LL, but BES1-RNAi plants show reduced CKL12 transcript compared to control when grown in LD and DD conditions (Figure 5B–D). Additionally, CKL12 transcript increases more rapidly in DD-grown than in LL-grown wild-type plants (Figure 5C and D). It therefore appears that the BR-dependent increase in CKL12 transcript depends on a dark phase, and this increase occurs in concert with elongation.
Three independent lines of evidence indicate possible brassinosteroid involvement in CKL12 function. 1) We observed reduced hypocotyl and root growth, both of which are regulated by BR, in ckl12 mutants (Figure 4). 2) Published data show that the CKL12 promoter is bound in vitro by the BR-related transcription factors BZR1 and BES1.36 3) RNAi knockdown of BZR1 and BES1 results in reduced CKL12 transcript (Figures 5B–D). The BR signaling pathway bears similarity to the WNT/β-catenin pathway in animals, with both being activated by extracellular signals through transmembrane receptors in concert with coreceptor proteins.24 Both have GSK3β-type kinases as central signal transduction factors. In animals, CK1 also plays important roles in the WNT signaling pathway, acting as a priming kinase to generate binding sites and promote the phosphorylation of target proteins by GSK3β.45 Genetic or biochemical studies of potential BIN2-CKL12 interactions may provide insight into the regulation of BR-related TFs or other parts of the pathway. The specific functions of CKL12 in the canonical BR signaling pathway are currently unknown, although CKL2 has been implicated in the cross-regulation of BR signaling by ABA-activated direct phosphorylation of the BR receptor BRI1.16 BR biosynthesis is feedback-regulated, with the expression of BR biosynthetic enzymes and the production of BR metabolic intermediates being downregulated by active BR signaling and in the stabilized bzr1-1D mutant.25 CKL-family kinases, possibly including CKL12 and CKL2, may also be regulated by a feedback mechanism, fine-tuning BR signal strength via the transcriptional output of BZR1/BES1 transcription factors (Figure 6).
A hypothetical model of CKL12 regulation by and of brassinosteroid signaling. Brassinosteroids are sensed by the receptor BRI1 and it's co-receptor BAK1 at the plasma membrane (PM). The activated receptor complex, through several steps, inhibits the GSK3βb-related kinase BIN2. BIN2 phosphorylation of the transcription factors (TFs) BES1 and BZR1 promotes nuclear export and degradation (the nucleus is indicated by the light blue circle), and inhibition of BIN2 allows the TFs to be dephosphorylated and enter the nucleus. Light also negatively regulates these TFs, consistent with reduced CKL12 transcript accumulation in LL conditions. BES1 family TFs regulate the transcription of many target genes, including CKL12. CKL12 transcription is promoted by BES1/BZR1 transcription factors and requires darkness. Alterations at the 3′ end of the CKL12 transcript, which may be an autoinhibitory domain, result in reduced elongation growth, particularly in the root, suggesting that these phenotypes reflect the activation of CKL12. The wild-type-like phenotype of the loss-of-function allele suggests functional redundancy, possibly involving CKL1, 2, and/or 5 (in gray). The targets of CKL12 kinase are not known. It may phosphorylate BRI1, as occurs with CKL2, or may regulate multiple parts of the BR pathway or other growth-related proteins (grey dashed lines). Ultimately, activation of the BR pathway leads to increased elongation growth, while CKL12 is produced downstream of BR signaling and may act to reduce growth.
Supplementary Material
Supplementary materialSeluzicki_CKL12_SupplementaryFigure1_20251025.docx
Supplementary materialSeluzicki_CKL12_SupplementaryTable1_20251025.xlsx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Peters JM, Mc Kay RM, Mc Kay JP, Graff JM. Casein kinase I transduces Wnt signals. Nature. 1999;401:345–350. doi: 10.1038/43830.10517632 · doi ↗ · pubmed ↗
- 2Vielhaber E, Eide E, Rivers A, Gao Z-H, Virshup DM. Nuclear entry of the circadian regulator m PER 1 is controlled by mammalian casein kinase I ɛ. Mol Cell Biol. 2000;20:4888–4899. doi: 10.1128/MCB.20.13.4888-4899.2000.10848614 PMC 85940 · doi ↗ · pubmed ↗
- 3Cheong JK, Virshup DM. Casein kinase 1: complexity in the family. Int J Biochem Cell Biol. 2011;43:465–469. doi: 10.1016/j.biocel.2010.12.004.21145983 · doi ↗ · pubmed ↗
- 4Knippschild U, Krüger M, Richter J, Xu P, García-Reyes B, Peifer C, Halekotte J, Bakulev V, Bischof J. The CK 1 family: contribution to cellular stress response and its role in carcinogenesis. Front Oncol. 2014;4:96. doi: 10.3389/fonc.2014.00096.24904820 PMC 4032983 · doi ↗ · pubmed ↗
- 5Graves PR, Roach PJ. Role of COOH-terminal phosphorylation in the regulation of casein kinase Iδ. J Biol Chem. 1995;270:21689–21694. doi: 10.1074/jbc.270.37.21689.7665585 · doi ↗ · pubmed ↗
- 6Cegielska A, Gietzen KF, Rivers A, Virshup DM. Autoinhibition of casein kinase I ε (CKIε) is relieved by protein phosphatases and limited proteolysis. J Biol Chem. 1998;273:1357–1364. doi: 10.1074/jbc.273.3.1357.9430669 · doi ↗ · pubmed ↗
- 7Harold RL, Tulsian NK, Narasimamurthy R, Yaitanes N, Ayala Hernandez MG, Lee H, Crosby P, Tripathi SM, Virshup DM, Partch CL. Isoform-specific C-terminal phosphorylation drives autoinhibition of Casein kinase 1. Proc Natl Acad Sci U S A. 2024;121:e 2415567121. doi: 10.1073/pnas.2415567121.39356670 PMC 11474029 · doi ↗ · pubmed ↗
- 8Ono A, Sato A, Fujimoto KJ, Matsuo H, Yanai T, Kinoshita T, akamichi N N. 3,4-dibromo-7-azaindole modulates Arabidopsis circadian clock by inhibiting casein kinase 1 activity. Plant Cell Physiol. 2019;60(11):2360–2368. doi: 10.1093/pcp/pcz 183.31529098 PMC 6839374 · doi ↗ · pubmed ↗