Otilonium bromide exhibits novel antifungal activity against Candida albicans via regulating iron homeostasis
Li-Hang Hsu, Yuk-Ping Chou, Tang-Long Shen, Daria Wieczorek, Ying-Lien Chen

TL;DR
Otilonium bromide, a drug used for irritable bowel syndrome, shows antifungal activity against Candida albicans by affecting iron balance and cell membrane integrity.
Contribution
The novel antifungal activity of otilonium bromide against C. albicans via iron homeostasis regulation is reported.
Findings
Otilonium bromide inhibits C. albicans at 2 μg/mL by damaging cell membranes and preventing yeast-to-hyphae transition.
Iron ions negate the antifungal effect of otilonium bromide, indicating a role for iron homeostasis in its mechanism.
Otilonium bromide showed limited efficacy in reducing C. albicans in a murine infection model.
Abstract
Traditional antifungal drugs used against Candida albicans have several drawbacks, including the emergence of drug-resistant strains. In addition, developing novel antifungal agents requires long-term research and design. Drug repurposing, identifying and utilizing previously unknown functions of known drugs, such as antifungal activity, may be a quick method for mining efficient alternatives. Otilonium bromide (OB), an FDA-approved drug, is a quaternary ammonium compound used as a therapeutic drug for irritable bowel syndrome. We previously reported the inhibitory effect of OB against the spore germination of Cryptococcus neoformans. In this study, we found that the antifungal activity of OB against C. albicans was 2 μg/mL for both minimum inhibitory and fungicidal concentrations. OB could destroy the cell membrane and prevent C. albicans from undergoing yeast-to-hyphae transition,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
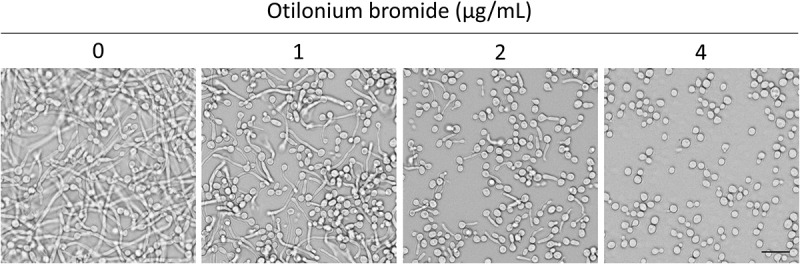 Figure 1
Figure 1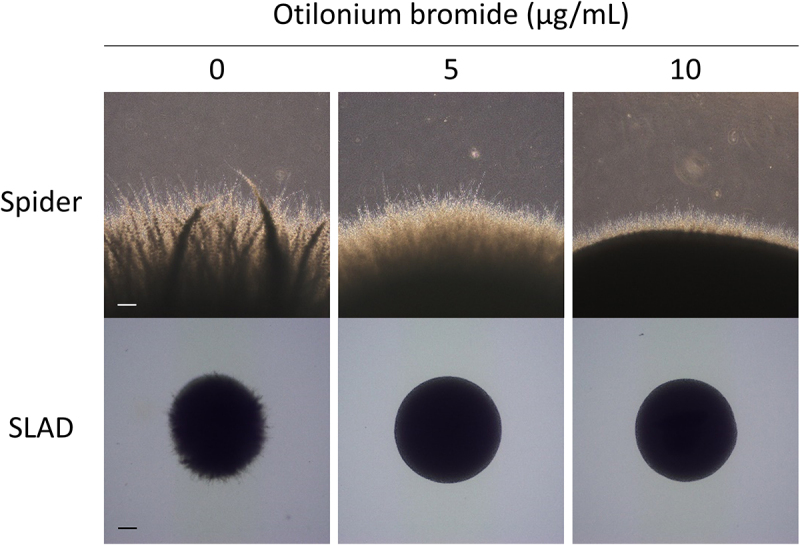 Figure 2
Figure 2 Figure 3
Figure 3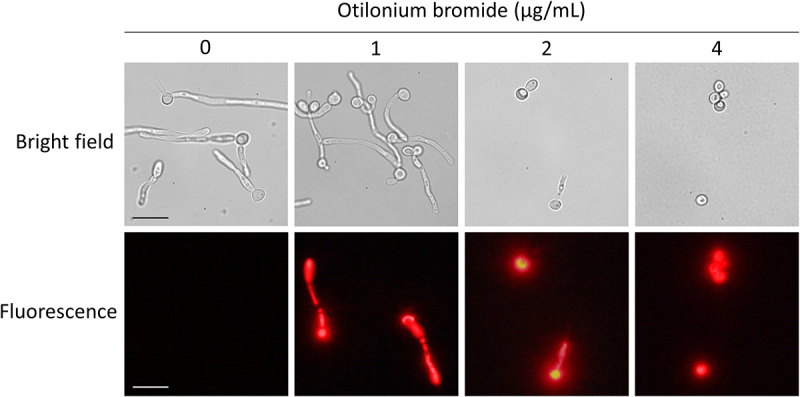 Figure 4
Figure 4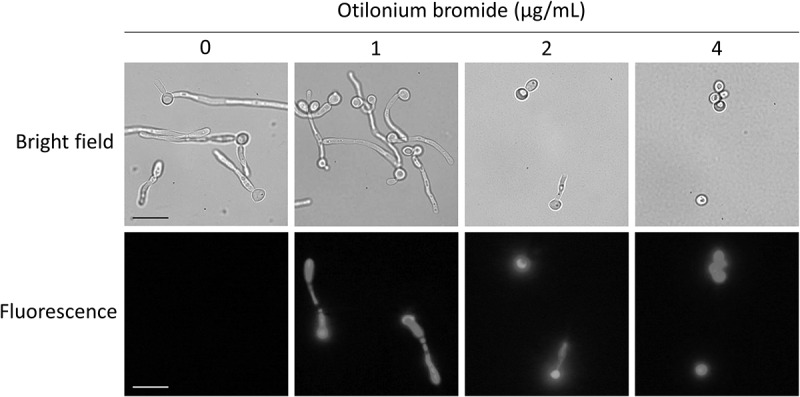 Figure 5
Figure 5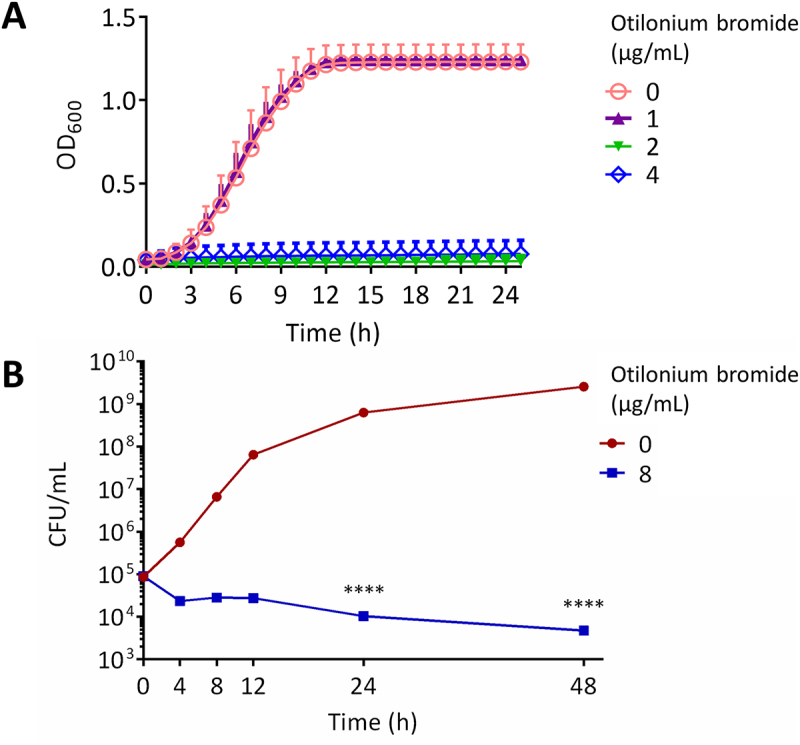 Figure 6
Figure 6 Figure 7
Figure 7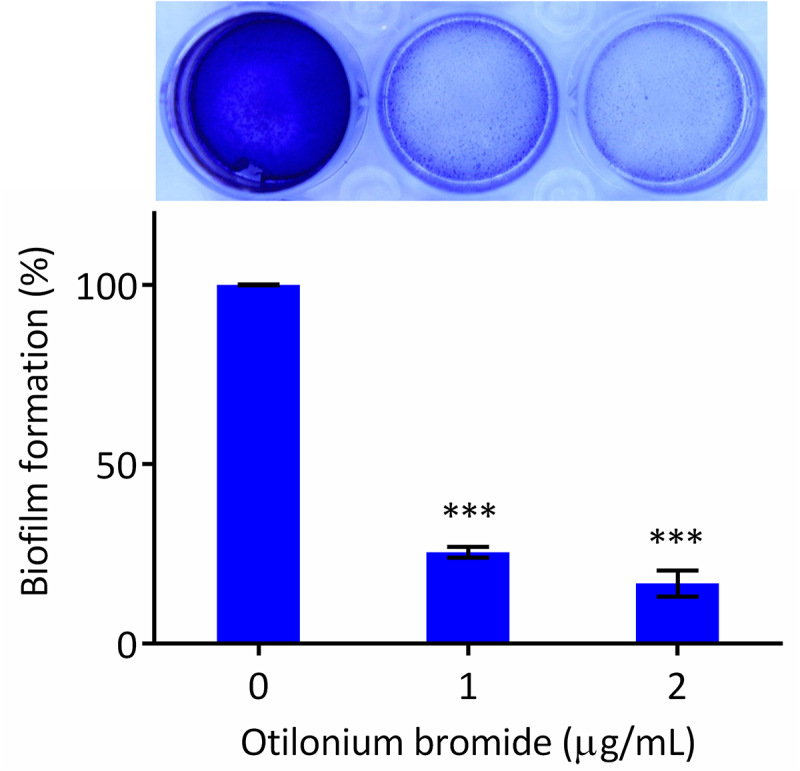 Figure 8
Figure 8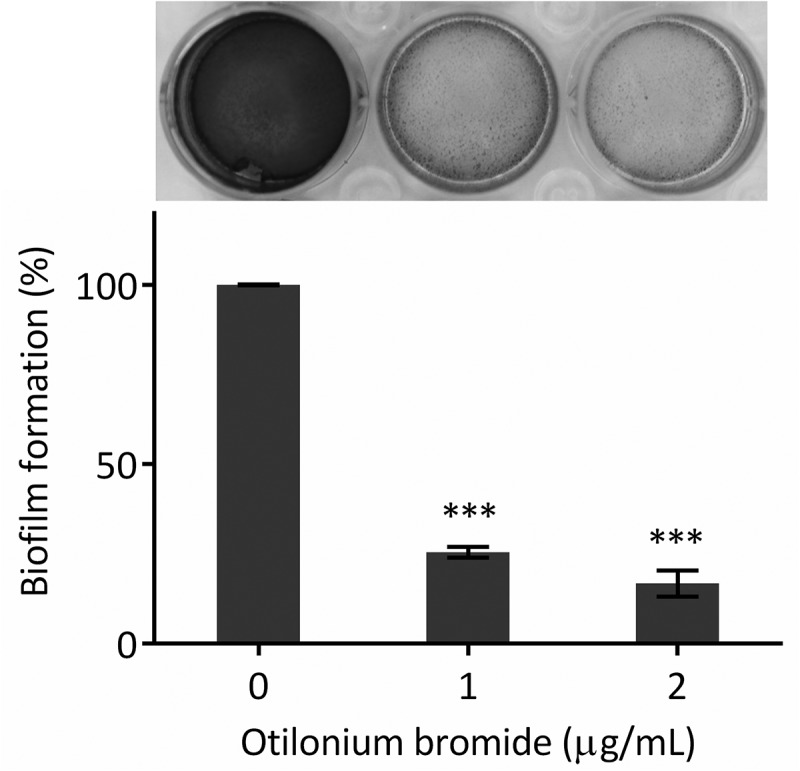 Figure 9
Figure 9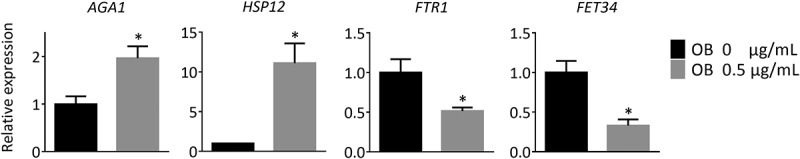 Figure 10
Figure 10 Figure 11
Figure 11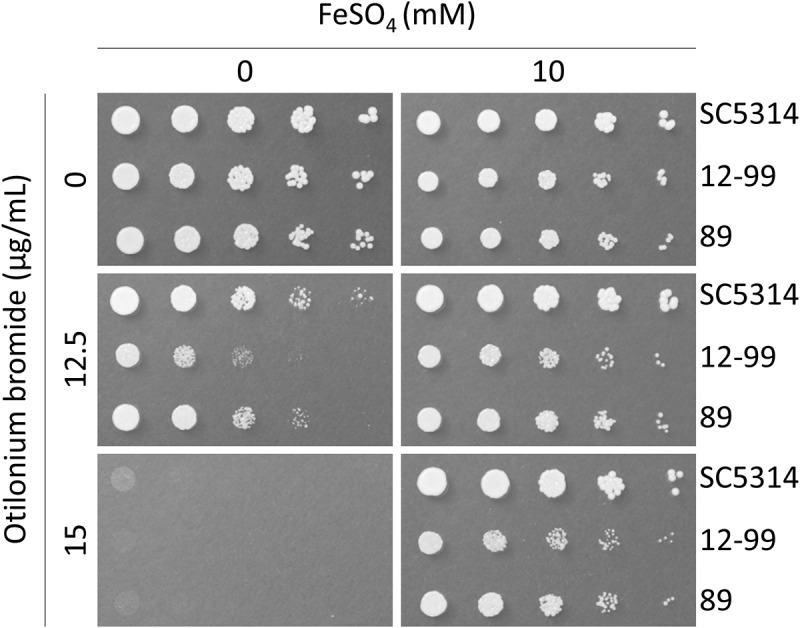 Figure 12
Figure 12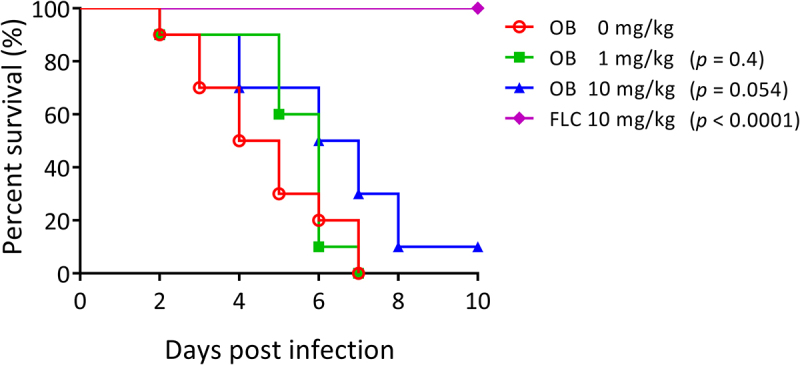 Figure 13
Figure 13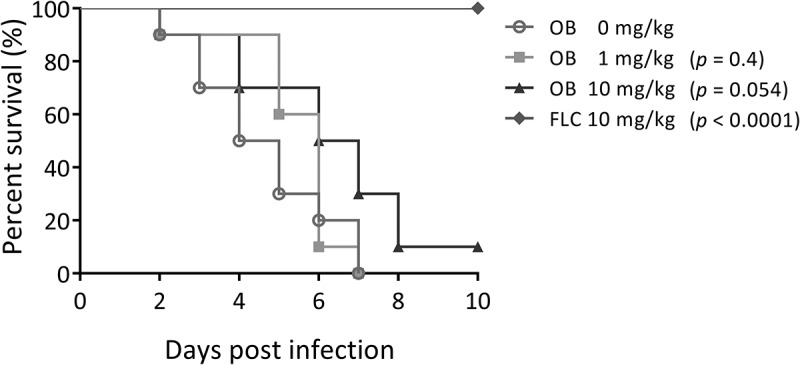 Figure 14
Figure 14- —National Science and Technology Council
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Infections and Studies · Antifungal resistance and susceptibility · Nail Diseases and Treatments
Introduction
Candida albicans is a commensal yeast found in 70% of healthy humans, especially on the skin and mucosal surfaces [1]. However, being an opportunistic pathogen, C. albicans is the dominant species that causes candidiasis in immunocompromised individuals, and it turns into candidemia if not treated appropriately [2]. Nowadays, antifungal drugs are categorized into three classes: azoles, echinocandins, and polyenes. Although the antifungal activity is significant, drug-resistant strains still evolve and survive from drug application [3]. In addition, due to high nephrotoxicity in some drugs and the increased cost of developing new antifungal drugs, alternative approaches for discovering antifungal agents are needed.
Traditional development of new antifungal drugs requires substantial time and cost investments, leading to increased interest in drug repurposing as an alternative strategy. Our previous screening of 1018 FDA-approved drugs identified otilonium bromide (OB), which demonstrated significant inhibitory effects against Cryptococcus neoformans germination [4]. This finding aligns with growing evidence that quaternary ammonium compounds (QACs) possess antimicrobial properties beyond their traditional applications. Several QACs have shown antifungal efficacy: benzalkonium chloride exhibits activity against Candida species through membrane disruption, while cetylpyridinium chloride demonstrates broad-spectrum antifungal effects [5,6]. Notably, our work with the QAC compound PMT12-BF4 revealed its antifungal mechanism involves interference with iron homeostasis [7], suggesting this pathway may be a common target for QAC-based antifungal activity.
Among the repurposed candidates, OB, an FDA-approved drug, demonstrated inhibitory activity on yeast germination in Cryptococcus neoformans, and the bacterial pathogen Acinetobacter baumannii [4,8]. OB is a quaternary ammonium compound (QAC) used to treat irritable bowel syndrome (IBS). Patients suffering from IBS have abnormal contraction of the intestinal tract, followed by cramps in the abdomen. In terms of its mechanism in mammals, OB inhibits the nerve impulse by blocking calcium ion channels and tachykinin receptors. Thus, those ligands are not accessible to the intestine, interrupting signal transduction and alleviating patient pain [9–11].
Trace elements such as calcium, iron, copper, or zinc are essential for the growth of most organisms. Among all the trace elements, iron is most closely associated with the survival and virulence of C. albicans in the host environment [12]. Iron sequestering thus plays an important role in humans to prevent C. albicans from invading, which is called nutritional immunity [12,13].
In this study, we focused on OB and its antifungal activity and found a potential pathway associated with iron homeostasis that inhibits the growth of C. albicans. Understanding this mechanism could provide valuable insights for developing more effective antifungal strategies through drug repurposing, particularly targeting iron homeostasis as a novel therapeutic approach.
Materials and methods
Strains, compounds, and media
The strains used in this study are listed in Table 1. OB (MedChemExpress, USA) as a pure compound was dissolved in dimethyl sulfoxide (DMSO, Scharlab, Spain) to make a 5 mg/mL stock solution for further experiments. Fluconazole (Pfizer, USA) in the form of 2 mg/mL and Catilon tablets containing 40 mg of OB per tablet (kindly provided by Tri-Service General Hospital, Taiwan) was used for the murine therapeutic assay.Table 1.Minimal inhibitory concentration (MIC) and minimal fungicidal concentration (MFC) of otilonium bromide against multiple human fungal pathogens.Strains (reference)DescriptionMICMFCCandida albicans SC5314 [14]Wild type22Candida albicans 12–99 [15]Fluconazole resistant22Candida albicans 89 [16]Echinocandin resistant22Candida albicans Δcna1/Δcna1 [17]Calcineurin mutant22Candida albicans Δcnb1/Δcnb1 [18]Calcineurin mutant22Candida dubliniensis CD36 [19]Wild type22Candida tropicalis MYA3404 [20]Wild type22Candida tropicalis DPL73 [21]Echinocandin resistant11Candida parapsilosis J941367 [20]Wild type11Candida glabrata CBS138 [22]Wild type88Candida glabrata DPL23 [23]Echinocandin resistant48Candida glabrata AmB-resistant isolate D [24]Amphotericin B resistant48Cryptococcus neoformans H99 [25]Wild type44Cryptococcus neoformans T1 [26]Fluconazole resistant22Cryptococcus neoformans 89–610 [26]Fluconazole resistant22Cryptococcus deuterogattii R265 [27]Wild type22Aspergillus fumigatus AF293 [28]Wild type44Aspergillus fumigatus A031 [29]Azole resistant3232
Media used in this study were YPD (1% yeast extract [Bioshop, Canada], 2% peptone [Bioshop], 2% dextrose [Bioshop]), PDB (24 g potato dextrose broth [Himedia, India] in 1 L distilled water), LB (0.5% yeast extract, 1% tryptone [Bioshop], 1% NaCl), RPMI 1640 (10.4 g RPMI 1640 powder [Sigma-Aldrich, USA], 34.5 g MOPS [3-(N-morpholino) propanesulfonic acid, Sigma-Aldrich], 2% dextrose, in 1 L distilled water, with pH adjusted to 7.0 with NaOH), Spider medium (10 g nutrient broth [Himedia], 10 g mannitol [Pancreac, Spain], 2 g K_2_HPO_4_, in 1 L distilled water, adjusted to pH 7.2 with H_3_PO_4_), SLAD (synthetic low ammonium dextrose, 0.17% yeast nitrogen base without amino acids or ammonium sulfate [Bioshop], 50 μM (NH_4_)2_SO_4, 2% dextrose), and YNB (0.17% yeast nitrogen base without amino acids or ammonium sulfate, 0.5% (NH_4_)2_SO_4, 2% dextrose). All media were solidified by adding 2% agar (Bioshop) if necessary.
Determination of antifungal activity
The minimal inhibitory concentrations (MICs) were obtained following the CLSI protocol M27-A3 and M38-A2. For yeasts, strains were cultured in 3 mL YPD broth medium overnight at 30°C, washed twice, and resuspended with ddH_2_O, and the concentration was determined using a hemocytometer. For Aspergillus fumigatus, strains were streaked out on a PDA agar plate and incubated for 5 days at 37°C. Spores were collected with sterile water, filtered through Miracloth (Millipore, USA) to remove hyphae, and the concentration was determined by hemocytometer. All inoculums were then diluted to the recommended concentrations with RPMI 1640 medium. OB was serially diluted 2-fold by RPMI 1640 medium, and 100 μL of the compound was added to a 96-well plate, followed by 100 μL of the inoculums. The final concentration of OB ranged from 0.125 - 64 μg/mL, and the final concentrations of the inoculums were 1.25 × 10^3^ cells/mL for yeasts and 2.5 × 10^4^ for A. fumigatus. Plates were incubated at 35°C for 48 h, and the MIC was defined as the lowest concentration at which no visible colony grew. Minimal fungicidal concentration (MFC) was determined after the MIC test. For each strain, 3 μL of wells containing OB at concentrations from 0.5 MIC (positive control) to the highest concentration (64 μg/mL) were thoroughly pipetted and subcultured onto drug-free YPD or PDA agar plates. The plates were incubated at an optimal temperature for growth of each fungal pathogen for 48 h and the MFC was determined as the drug concentration at which no colonies formed.
Time-kill kinetic assay
Overnight cultures of C. albicans SC5314 were washed twice with ddH_2_O and inoculated into YPD broth to a volume of 3 mL (10^5^ CFU/mL) with or without 8 μg/mL OB at 30°C. A 100 μL aliquot was removed from each culture at the indicated time points (0, 4, 8, 12, 24, and 48 h), appropriately diluted with ddH_2_O, plated on a fresh YPD agar plate and incubated at 30°C for 48 h before colony count. Two-way ANOVA was performed using GraphPad Prism 6, and a p value of less than 0.05 represented a significant difference.
Biofilm formation assay
Overnight cultures of C. albicans SC5314 were washed twice with PBS and diluted with 2 mL Spider medium to 0.5 OD_600_. OB was added at the indicated concentrations after 1 h incubation at 37°C for cell adhesion. After incubation for 48 h, the medium and suspended cells were removed by pipetting; the biofilm was washed with PBS twice, stained with 0.4% crystal violet and destained with 99% EtOH. The OD_595_ was measured after the destaining step, and wells without OB were defined as having 100% biofilm formation. Unpaired t-tests were performed using GraphPad Prism 6, and a p value of less than 0.05 represented a significant difference.
Hyphae induction and membrane penetration assay
For hyphal induction assay, C. albicans SC5314 cells cultured in YPD broth overnight at 30°C were washed twice with ddH_2_O and diluted with 2 mL RPMI 1640 medium to OD_600_ 0.25 in a 12-well plate (Nest Biotechnology, China). OB was added into wells at the concentrations of 0, 1, 2, or 4 μg/mL, with three replicates. The plate was incubated at 37°C for 3 h for germ tube induction and observed with an inverted microscope (Olympus, Japan) at 400X magnification. Cells were photographed using a camera connected to Olympus cellSens Entry 2.1 software. In addition, cells were also adjusted to 10^3^ cells/mL with ddH_2_O and 50 μL of cells were plated on solid Spider or SLAD medium with 0, 5, or 10 μg/mL OB. After incubation for 24 h at 37°C, colonies were observed using a microscope at 40X.
For membrane penetration assay, the induced germ tube was washed with ddH_2_O, stained with 1 μg/mL propidium iodide, and observed with a fluorescence microscope (Olympus BX53) at 1000X magnification.
RNA-sequencing experiments
RNA extraction was performed following the previous study [7]. In brief, overnight cultures of C. albicans SC5314 cells were washed twice with sterile water, and adjusted to OD_600_ 0.25 with 5 mL fresh YPD in the presence or absence of 0.5 μg/mL OB. The cultures were incubated at 30°C for 3 h with shaking at 200 rpm before RNA extraction. TRIzol reagent (Invitrogen, USA) was used in total RNA extraction following the manufacturer’s protocol. The 1 mL TRIzol was added to collected cells and alternately vortexed for 30 s with 2 mm beads and put on ice for 30 s five times. After being centrifuged at 4°C/12,000 g for 10 min, the supernatant was transferred to a new tube and incubated for 5 min at room temperature. The 200 μL of chloroform was then added and mixed thoroughly, and tubes were incubated at room temperature for 3 min, centrifuged at 4°C/12,000 g for 15 min. The upper layer of supernatant was separated to another tube, and 400 μL isopropanol was added for RNA precipitation. After incubated at room temperature for 10 min, the tubes were centrifuged at 4°C/12,000 g for 10 min, and the supernatant was discarded. The RNA pellet was washed twice with 75% ethanol and resuspended with RNase-free water.
The Next Generation Sequencing (NGS) library construction using RNA was as described in a previous study [30]. The mRNA was amplified with oligo(dT) magnetic beads and shortened into approximately 200-base fragments. The double-strand cDNAs were synthesized using a random hexamer, dNTPs, buffer, and RNase H (for the first strand), and DNA polymerase I (for the second strand). The double-strand cDNAs were purified, and end preparation and 3′ end single nucleotide adenine addition were performed. Sequencing adaptors were then ligated to the fragments and amplified by PCR. Sample library was qualified and quantified with an Agilent 2100 bioanalyzer and ABI StepOne Plus real-time PCR system. Sequencing of the library was done by an Illumina H-Seq 2000 instrument.
Real-time qRT-PCR
A Turbo DNA-free kit (Invitrogen, USA) was used in removing genomic DNA, and 1 μg of total RNAs were reverse transcribed to cDNA using a high-capacity cDNA reverse transcription kit (Applied Biosystems, USA). Real-time PCR was then performed in mixtures including 1 μL of cDNA (1 ng), 5 μL of Fast SYBR green master mix (Applied Biosystems), 1 μL of 5 μM forward primer, and 1 μL of 5 μM reverse primer. Primer pairs used in real-time PCR are listed in Supplementary Table S1 (Table S1). Quantitative PCR conditions were set as follows: 95°C/7 min denaturation, 95°C/10 s, and 60°C/30 s (40 cycles) amplification; 95°C/15 s, and then 60 s each at 0.3°C increments between 60°C and 95°C (for melting curve). Cycle threshold (C_T_) values were obtained using a StepOnePlus system and StepOne software (v.2.3), and the relative gene expressions (2^-ΔΔCT^) were calculated based on ACT1-calibrated values. The relative expression levels of C. albicans genes in the presence of 0.5 μg/mL OB were normalized to those in the absence of OB, and the diagrams were depicted by GraphPad Prism 6.0 software. Significant differences were analyzed using an unpaired t test (p < 0.05).
Spot plating assay
C. albicans SC5314, and two drug-resistant C. albicans 12–99 and 89 isolates were incubated in 30°C overnight, washed twice and adjusted to 0.2 OD_600_ with ddH_2_O. Three microliters of cell suspensions in 5-fold serial dilution were transferred to YNB agar plates containing OB (0, 12.5, or 15 μg/mL) or FeSO_4_ (0 or 10 μg/mL). Plates were incubated at 30°C for 48 h and then photographed.
Therapeutic assay
For inoculum preparation, cells of C. albicans SC5314 were grown overnight in liquid YPD medium, washed twice with phosphate-buffered saline (PBS), and adjusted to a concentration of 5 × 10^6^ cells/mL with PBS. Five- to six-week-old male ICR mice (BioLasco Taiwan, Taiwan) were inoculated with 10^6^ C. albicans cells in 200 μL by tail-vein injection. For oral administration, OB was dissolved in a solvent containing 50% Kolliphor (Sigma-Aldrich, USA) in absolute ethanol (Bioman Scientific, Taiwan), and 10-fold diluted with PBS. Then, OB or fluconazole (FLC) was administered orally to mice at the indicated concentrations at 4, 24, 48, 72 h after the inoculation. Ten mice per group were used, and all mice were monitored for 10 days. The mice which lost 20% weight or more after inoculation were euthanized and counted deaths. For euthanasia, the mice were placed in a sealed chamber, and carbon dioxide was introduced (flow rate: 10% − 30% volume/min; flowmeter reading between 1 and 2 liters per minute). Once the animal exhibited no apparent signs of life, the chamber was opened to allow carbon dioxide to dissipate for approximately 2 minutes, after which cervical dislocation was performed as a secondary method to ensure euthanasia. The euthanasia procedures were carried out in accordance with the American Veterinary Medical Association. All mice were included in the experiment, and data were statistically analyzed using a log-rank test using GraphPad Prism 6. All animal experiments were conducted in the Animal Resource Center at National Taiwan University (NTU), and adhered to the ARRIVE guidelines.
Results
Otilonium bromide possessed broad-spectrum antifungal activity
In our previous study, a collection of 1018 FDA-approved drugs was screened and assessed for anticryptococcal activity [4]. Among these drugs, OB was able to inhibit Cryptococcus neoformans and Cryptococcus deuterogattii, and we found that it also inhibited Candida species. C. albicans strains, including strain SC5314 and drug-resistant isolates 12–99 and 89 were inhibited by OB with MIC at 2 μg/mL. To further investigate whether the antifungal activity of OB involves the calcineurin signaling pathway, we tested its effects on calcineurin-deficient C. albicans mutants Δcna1/Δcna1 and Δcnb1/Δcnb1. These mutants are known to exhibit defects in stress responses and cell wall integrity, often resulting in hypersensitivity to various antifungal agents. OB retained antifungal activity against both mutants, suggesting that its mode of action does not require a functional calcineurin pathway. This finding implies that OB likely acts through a mechanism independent of calcineurin signaling. Although similar results were found in Candida tropicalis, Candida glabrata was more tolerant to OB as the MIC was higher at 8 μg/mL (Table 1). The fungicidal activity of OB was then investigated after the susceptibility test. All tested strains treated with OB at respective MICs were unable to grow when transferred to fresh YPD medium, indicating that OB possessed fungicidal activity and effectively eliminated these pathogens.
Otilonium bromide inhibited yeast-to-hyphae transition in C. albicans
Yeast-to-hyphae transition is an important virulence factor for C. albicans to infect its host; thus we observed whether OB could prevent yeast cells from germination. With exposure to OB for 3 h, few cells were still showing hyphal growth at the concentration of 2 μg/mL OB and lost the ability to germinate and maintain yeast form at 4 μg/mL (Figure 1), suggesting that C. albicans were inhibited from germination as the concentration of OB increased. In addition, C. albicans was incubated on solid agar medium in hyphal-inducing conditions to evaluate hyphal growth. As the amount of OB increased, the hyphae growing on the edge of the colony became short, suggesting the inhibition of yeast-to-hyphal transition was concentration-dependent (Figure 2). Figure 1.Otilonium bromide inhibited C. albicans from germination.C. albicans SC5314 overnight culture was washed twice with ddH_2_O, diluted with 2 mL RPMI 1640 medium to 0.25 OD_600_, and otilonium bromide was added at indicated concentrations. After 3 h incubation at 37°C, cells were observed with an optical microscope at 400X. Scale bar = 20 μm. Figure 2.Otilonium bromide inhibited hyphal growth in C. albicans.C. albicans SC 5314 overnight culture was washed twice and diluted with ddH_2_O to 100 cells/mL, and 50 μL was spread on Spider or SLAD agar plates with 0, 5, or 10 μg/mL otilonium bromide. After 24 h incubation at 37°C, colonies were observed using a microscope at 40X. Scale bar = 200 μm.
Otilonium bromide caused membrane damage in C. albicans
C. albicans exhibited reduced growth while being treated with OB; therefore, we further tested the viability of these cells through propidium iodide (PI) staining. Consistently, PI as a membrane impermeable stain, which can distinguish live cells, could diffuse into cells treated with OB at 1 μg/mL and showed partial staining compared to untreated cells (Figure 3). On the other hand, at both 2 and 4 μg/mL of OB, C. albicans cells were fully stained, and remained at low optical density when measured with a spectrophotometer (Figure 4(A)). Time-kill assay revealed that C. albicans cells were significantly decreased after incubation for 24 h with OB, and >90% cells were eliminated after 48 h (Figure 4(B)). In brief, C. albicans was killed through the membrane damage induced by OB, which functions as a fungicidal compound. Figure 3.Otilonium bromide caused membrane damage in C. albicans.C. albicans SC5314 overnight culture was washed twice with ddH_2_O, diluted with 2 mL RPMI 1640 medium to 0.25 OD_600_, and otilonium bromide was added at indicated concentrations. After 3 h incubation at 37°C, cells were washed twice with ddH_2_O, stained with 1 μg/mL propidium iodide (PI), and observed with a fluorescence microscope at 1000X. Scale bar = 5 μm. Figure 4.Cells of C. albicans were prevented from growth and were eliminated while treated with otilonium bromide.(A). C. albicans overnight culture was washed twice with ddH_2_O, diluted to OD_600_ = 0.02 using 200 μL YPD broth medium with otilonium bromide at 0, 1, 2, or 4 μg/mL, and incubated at 30°C. The OD_600_ values were measured every hour. (B). C. albicans SC5314 overnight culture was washed twice with ddH_2_O, diluted to 10^5^ cells/mL using 3 mL YPD broth medium with otilonium bromide at 0 or 8 μg/mL, and incubated at 30°C/200 rpm. The 100 μL of culture at each time point (0, 4, 8, 12, 24, or 48 h) were properly diluted, plated on YPD agar medium, and incubated at 30°C for 48 h before colony count. **** represented p-value < 0.0001.
Biofilm formation was reduced in C. albicans treated with otilonium bromide
Biofilm formation is also a virulence factor in C. albicans after its invasion and filamentation and is sometimes related to drug resistance [31]. Therefore, crystal violet was used to stain the biofilm structure to investigate how effective OB was in reducing biofilm formation in C. albicans. Cells adhered to the bottom of wells were allowed to form biofilm in the absence of OB. The percentage of biofilm formation decreased more than 90% and more than 95% in the presence of 2 μg/mL and 4 μg/mL OB, respectively (Figure 5). As a result, OB could significantly reduce the biofilm formation through a potential mechanism that destroyed the cell membrane and interrupted filamentation in C. albicans. Figure 5.Biofilm formation was reduced in C. albicans treated with otilonium bromide.Overnight culture of C. albicans SC5314 was diluted with Spider medium to 0.5 OD_600_, and otilonium bromide was added after 1 h incubation at 37°C for cell adhesion at indicated concentrations. After 48 h incubation, biofilm was stained with crystal violet and destained with EtOH. The OD_595_ was measured after destaining, and wells without otilonium bromide were set as 100% biofilm formation. The asterisk represents significant difference (unpaired t-test, p < 0.05).
RNA sequencing analysis
To investigate the potential targets of OB in C. albicans, we conducted RNA sequencing. We found 143 genes with significantly differential gene expressions (DGEs), defined as equal to or more than a 4-fold change in gene expression. Among these genes, 108 were upregulated, and 35 were downregulated in C. albicans treated with 0.5 μg/mL OB (Table 2). The main functions of these genes included oxidoreductase activity, heme binding, and tetrapyrrole binding, and all the functions analyzed from gene ontology indicated metal ions might play a critical role in OB-mediated gene regulation. For instance, Fet34 and Ftr1 controlled iron homeostasis in C. albicans, and heme binding proteins such as Rbt5 were involved in iron acquisition in the host environment; tetrapyrrole functioned as a metal ion chelator. Furthermore, heat shock proteins and cell wall-related genes were also regulated. To confirm the relative gene expression obtained from RNA sequencing, qRT-PCR experiments were conducted, and 2 genes with significant DGE were selected relatively from up- and down-regulated genes (Figure 6). All of the sequencing data were submitted to NCBI Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE189386, token: mfczcciyfxodteb). Figure 6.Relative gene expressions in C. albicans treated with otilonium bromide.Relative gene expressions were calculated through real-time qRT-PCR based on ACT1-calibrated and 2^−ΔΔCT^ values. The relative expression levels of C. albicans genes in the presence of 0.5 μg/mL otilonium bromide were normalized to those in the absence of otilonium bromide. The asterisk represents significant difference (unpaired t-test, p < 0.05).Table 2.Otilonium bromide-mediated gene regulation.CGDIDGeneDescriptionlog_2_FCGenes upregulated in C. albicans treated with otilonium bromide (108/143)CAL0000180646AGA1Protein with similarity to agglutinin subunit8.99CAL0000197297WH11White-phase yeast transcript; expression in opaques7.82CAL0000177418C7_00870WPutative guanine deaminase7.09CAL0000198254C1_02800WProtein of unknown function6.85CAL0000177213PLB1Phospholipase B6.44CAL0000198834RBR1Glycosylphosphatidylinositol (GPI)-anchored cell wall protein6.44CAL0000188028CR_03320Chypothetical protein6.18CAL0000191795ADH3Putative NAD-dependent (R,R)-butanediol dehydrogenase5.99CAL0000179373CPD2Protein with homology to NADH dehydrogenase5.98CAL0000182697HSP12Heat-shock protein5.66CAL0000180850C5_02110WPutative heat shock protein5.66CAL0000176228C2_07630CPossible stress protein5.63CAL0000187233ASR1Heat shock protein5.33CAL0000189971RTA37-transmembrane receptor protein5.21CAL0000175449CSH1Aldo-keto reductase5.09CAL0000179752C1_10170WPutative adhesin-like protein4.84CAL0000189325INO1Inositol-1-phosphate synthase4.61CAL0000175827C1_04010CProtein with a NADP-dependent oxidoreductase domain4.49CAL0000182669CSP37Hyphal cell wall protein4.47CAL0000190475ASR2Adenylyl cyclase and stress-responsive protein4.46CAL0000199237C2_02220Chypothetical protein4.26CAL0000180798TNA1Putative nicotinic acid transporter4.01CAL0000186516CDR1Multidrug transporter of ABC superfamily3.76CAL0000191119PDR16Phosphatidylinositol transfer protein3.64CAL0000189321C4_03000CPlanktonic growth-induced gene3.38CAL0000194637C1_14060WGlucan 1,3-beta-glucosidase3.34CAL0000180820C2_02910Whypothetical protein3.34CAL0000197271C2_08890Whypothetical protein3.31CAL0000190633SOU1Enzyme involved in utilization of L-sorbose3.31CAL0000197522C3_03460CProtein of unknown function3.27CAL0000177800YDC1Alkaline dihydroceramidase; involved in sphingolipid metabolism3.25CAL0000176590DDR48Immunogenic stress-associated protein3.22CAL0000187177SOU2Protein similar to Sou13.22CAL0000175241UGA6Putative GABA-specific permease3.11CAL0000198101C5_00810CHas domain(s) with predicted heme binding and peroxidase activity3.10CAL0000191057C1_11200WPredicted mucin-like protein3.04CAL0000197958ROA1Putative PDR-subfamily ABC transporter involved in azole sensitivity3.04CAL0000197123C7_00770Whypothetical protein3.03CAL0000191729ECM331GPI-anchored protein3.00CAL0000176060AQY1Aquaporin water channel2.94CAL0000190248ARG1Argininosuccinate synthase2.93CAL0000197450IFU5Predicted membrane protein involved in cell wall maintenance2.90CAL0000178211AHP1Alkyl hydroperoxide reductase2.90CAL0000190001C2_07440COrtholog(s) have sterol esterase activity2.86CAL0000199842C6_03280Whypothetical protein2.83CAL0000184548CR_09140CProtein in directing meiotic recombination events2.83CAL0000174577OYE32NAD(P)H oxidoreductase family protein2.81CAL0000199973C2_06630Chypothetical protein2.78CAL0000178196PGA13GPI-anchored cell wall protein involved in cell wall synthesis2.78CAL0000186885C2_03110WProtein of unknown function2.74CAL0000180384YMX6Putative NADH dehydrogenase2.72CAL0000193774RME1Zinc finger protein2.72CAL0000189501TAC1Zn(2)-Cys(6) transcriptional activator of drug-responsive genes2.71CAL0000197194PST1Flavodoxin-like protein involved in oxidative stress protection2.70CAL0000190905MNN41Ortholog(s) have enzyme activator activity2.69CAL0000200522C7_04090CPredicted mitochondrial cardiolipin-specific phospholipase2.59CAL0000196854ECM4Cytoplasmic glutathione S-transferase2.59CAL0000187831MOH1Ortholog of S. cerevisiae Moh1, essential for stationary phase growth2.59CAL0000188193AAP1Putative amino acid permease2.58CAL0000198578C5_04470CPredicted RNA binding protein2.56CAL0000189861GRP2NAD(H)-linked methylglyoxal oxidoreductase2.45CAL0000193318ARE2Acyl CoA: sterol acyltransferase2.43CAL0000198473C1_10280CPutative protein of unknown function2.40CAL0000178438TRY6Helix-loop-helix transcription factor2.40CAL0000200650C1_05440CProtein of unknown function2.39CAL0000195719GRX1Putative glutaredoxin2.39CAL0000185031C2_10650WProtein of unknown function2.37CAL0000194072GLX3Glutathione-independent glyoxalase2.36CAL0000190671C2_06890CSimilar to oxidoreductases2.36CAL0000180176C1_10240CProtein of unknown function2.34CAL0000183255C1_03870CPredicted heme-binding stress-related protein2.34CAL0000194858SAP4Secreted aspartyl proteinase2.33CAL0000182133MRF1Putative mitochondrial respiratory protein2.29CAL0000200153CR_08670CProtein with an enoyl-CoA hydratase-related domain2.29CAL0000191751ASR3Adenylyl cyclase and stress-responsive protein2.28CAL0000180926C3_04310Chypothetical protein2.27CAL0000201827PGA7GPI-linked hyphal surface antigen2.27CAL0000198851C7_02140Whypothetical protein2.27CAL0000190381C1_11950WProtein of unknown function2.26CAL0000195131RBT5GPI-linked cell wall protein2.25CAL0000193825C3_02140Chypothetical protein2.24CAL0000191735XKS1Putative xylulokinase2.24CAL0000201116CAN1Basic amino acid permease2.22CAL0000201829IFD6Aldo-keto reductase2.21CAL0000174172LCB4Putative sphingosine kinase2.21CAL0000191559C6_00930Chypothetical protein2.18CAL0000177288ARG8Putative acetylornithine aminotransferase2.18CAL0000195767ARA1D-Arabinose dehydrogenase2.17CAL0000179259C7_02920WHas domain(s) with predicted carbon-nitrogen ligase activity2.16CAL0000182975C4_05250WPutative ubiquitin-protein ligase2.16CAL0000181678C1_12140WHas domain(s) with predicted cofactor binding activity2.10CAL0000193984C4_02620CCarbohydrate kinase domain-containing protein2.09CAL0000201755TFS1Putative carboxypeptidase y inhibitor2.08CAL0000191430C6_03050CProtein of unknown function2.08CAL0000192412UGA2Predicted succinate semialdehyde dehydrogenase2.08CAL0000174960CIP1Possible oxidoreductase2.08CAL0000181317ECI1Protein similar to S. cerevisiae Eci1p2.08CAL0000189087C1_10360CPutative protein of unknown function2.07CAL0000193502C4_04140WProtein with a predicted endonuclease/exonuclease/phosphatase2.06CAL0000188954C4_02340WPutative protease B inhibitor2.06CAL0000185914C4_02740WProtein of unknown function2.05CAL0000196224C1_03150Chypothetical protein2.05CAL0000177093ERG2C-8 sterol isomerase2.04CAL0000192083OSM2Putative mitochondrial fumarate reductase2.04CAL0000178766C3_00210CPredicted integral membrane protein2.03CAL0000179816AMS1Putative alpha-mannosidase2.02CAL0000175340DLD1Putative D-lactate dehydrogenase2.02CAL0000177748C2_09590COrtholog(s) have copper ion transmembrane transporter activity2.01Genes down-regulated in C. albicans treated with otilonium bromide (35/143)CAL0000198669JEN2Dicarboxylic acid transporter−6.66CAL0000188676SOD5Cu-containing superoxide dismutase−5.31CAL0000193948C1_09500Whypothetical protein−5.02CAL0000194292IHD1GPI-anchored protein−4.92CAL0000187820C6_02330WDescribed as a Gag-related protein−4.65CAL0000196552C5_02130WProtein of unknown function−4.50CAL0000180602TEF4Putative translation elongation factor−4.08CAL0000177231C1_07160CProtein conserved among the CTG-clade−3.97CAL0000181510NAG3Putative MFS transporter−3.96CAL0000185056FTR1High-affinity iron permease−3.83CAL0000192466FET34Multicopper ferroxidase−3.75CAL0000198258C3_02790Whypothetical protein−3.41CAL0000181671PGA26GPI-anchored adhesin-like protein of the cell wall−2.99CAL0000174751C1_11320CProtein of unknown function−2.96CAL0000186869MRV2Protein of unknown function−2.95CAL0000188660HAK1Putative potassium transporter−2.88CAL0000182549C6_01780CPredicted chloride transporter−2.82CAL0000197736CR_09350CProtein with a predicted heme oxygenase domain−2.80CAL0000174928C1_08900WProtein of unknown function−2.79CAL0000177400C5_03670CHas domain(s) with predicted metal ion binding activity−2.59CAL0000197551FCY2Purine-cytosine permease of pyrimidine salvage−2.57CAL0000193702C1_09210CPutative transporter−2.50CAL0000191596C5_03770CProtein similar to Candida boidinii formate dehydrogenase−2.42CAL0000180074FDH1Formate dehydrogenase−2.42CAL0000185898CFL4C-terminus similar to ferric reductases−2.42CAL0000186897SLP3Plasma membrane protein implicated in stress response−2.40CAL0000200616HGT1High-affinity MFS glucose transporter−2.35CAL0000200315DPP3Protein similar to S. cerevisiae pyrophosphate phosphatase Dpp1−2.29CAL0000198089GAP4High-affinity S-adenosylmethionine permease−2.18CAL0000192266C3_01540WPlasma-membrane-localized protein−2.12CAL0000182573C3_06950Whypothetical protein−2.07CAL0000190637FRP6Putative ammonia transport protein−2.06CAL0000198189C2_00760CProtein of unknown function−2.06CAL0000177585SEO1Protein with similarity to permeases−2.05CAL0000179202YHB1Nitric oxide dioxygenase−2.01
Iron acquisition was interrupted by otilonium bromide in C. albicans
According to our present research, quaternary ammonium compounds (QACs) could inhibit C. albicans through a pathway like nutrient immunity in humans [7]. Based on this result, combined with the RNA sequencing data, we inferred that OB, also a QAC, would affect iron homeostasis in C. albicans. Spot plating assay on YNB agar medium showed that colonies of C. albicans hardly grew, but recovered after the addition of iron, suggesting that sufficient iron was required for C. albicans to conquer the fungicidal activity of OB (Figure 7). Figure 7.The inhibition of otilonium bromide against C. albicans was interrupted by the addition of FeSO_4_.Overnight culture of C. albicans SC5314, and two drug-resistant C. albicans 12–99 and 89 isolates were washed twice and adjusted to OD_600_ = 0.2 with ddH_2_O. Three microliters of 5-fold serial diluted cell suspensions were dropped onto YNB agar plates with or without otilonium bromide or FeSO_4_ at the indicated concentrations. Plates were incubated at 30°C for 48 h before being photographed.
Otilonium bromide exhibited marginal therapeutic efficacy in the murine model of systemic infection
A therapeutic assay in a murine model was conducted to investigate the efficacy of OB. Mice treated with 0 or 1 mg/kg OB had all died by the 7th day after C. albicans inoculation, while 10% those treated with 10 mg/kg still survived at the endpoint of the experiment (p = 0.054, Figure 8). This result represented a marginal therapeutic effect for OB in curing C. albicans infected mice, and this effect was likely dose-dependent. Figure 8.Otilonium bromide exhibited marginal therapeutic efficacy in murine model of systemic candidiasis.Cells of C. albicans SC5314 were grown overnight in YPD medium, washed twice with PBS buffer, and adjusted to a concentration of 5 × 10^6^ cells/mL with PBS buffer. Mice were inoculated with 10^6^ C. albicans cells in 200 μL by tail-vein injection, and the oral administration of otilonium bromide (OB) or fluconazole (FLC) at indicated concentrations were conducted at 4, 24, 48, 72 h after the inoculation. Ten mice per group were used, and all mice were monitored for 10 days, and data were statistically analyzed by log-rank test.
Discussion
It was recently reported that OB can inhibit the germination of C. neoformans, and also possesses an inhibitory effect against A. fumigatus and C. albicans [4,32]. Based on this, we found that OB not only prevented C. albicans from yeast-to-hyphae transition but also killed yeast or early germinated cells, indicating the novel antifungal activity of this drug. OB has been used to treat irritable bowel syndrome (IBS). Interestingly, an increased amount of C. albicans could be found in IBS patients [33,34], suggesting fungal dysbiosis may lead to gastrointestinal dysfunctions, and the disease might be cured with OB after future development.
Yeast-to-hyphae transition is an important virulence factor in C. albicans, and it could colonize and form biofilm in the kidney, spleen or liver to defend against foreign compounds, including antifungal drugs during infection [35]. This study found that OB inhibited C. albicans from such transition in a dose-dependent manner and considerably reduced biofilm formation, indicating that the virulence and defensive ability of C. albicans against antifungal drugs may decrease. Nevertheless, in our therapeutic assay, OB showed marginal therapeutic efficacy in a murine model of systemic infection. Since OB suppresses IBS by blocking Ca^2+^ channels or tachykinin receptors on nerve cells of the intestinal wall, it could be inferred that there might be a lack of sufficient OB to inhibit pathogens in the kidneys, spleens or liver, in which C. albicans usually colonized [9]. Thus, we speculate that providing a higher dose of OB may increase the therapeutic efficacy. However, there was no significant therapeutic effect while mice were treated with a higher dosage (20 mg/kg) as well as intermediate doses (2 and 5 mg/kg) (Fig. S1). Meanwhile, further chemical modifications of the OB molecule may enhance therapeutic activity against C. albicans. Indeed, our recent study showed that quaternization with N-butylsulfonate may improve antifungal properties of the OB derivatives, and the incorporation of a 3-aminobenzoic acid moiety had a favorable impact on the compounds, further enhancing their antifungal efficacy. However, these derivatives did not display an MIC against C. albicans lower than that in OB with original structure [36].
QACs were found to have broad-spectrum antimicrobial activity, due to their activity on the bacterial cell membrane [37,38]. In our previous study, we found that another QAC, PMT12-BF4, also inhibited fungal pathogens such as C. albicans, non-albicans Candida species and A. fumigatus [7]. Similarly, the inhibitory effect of OB was abolished by replenishing iron ions. RNA sequencing data showed that many transcripts associated with iron homeostasis were expressed differentially in C. albicans treated with OB, indicating that OB might inhibit C. albicans through destabilizing iron homeostasis. Iron deficiency also mediated cell wall composition by, for example, decreasing β-1, 3-glucan in C. albicans. Under high iron conditions, C. albicans was able to resist cell wall-perturbing antifungals [39]. Compared to our spot plating assay, the non-sufficient iron conditions caused by OB might indirectly decrease the cell wall integrity of C. albicans, resulting in growth reduction.
The relationship between calcium signaling and iron homeostasis in fungal pathogens has recently emerged as an important area in antifungal research. While OB is known to affect calcium channels in mammalian cells [9], our study revealed its impact on iron homeostasis in C. albicans. Recent research has shown that calcium and iron homeostasis are intricately linked in fungal pathogens. In brief, iron deficiency may lead to calcium overload, while calcium supplementation could suppress iron uptake-related gene expression [40]. Specifically, under iron-deficient conditions, cells may mistakenly absorb calcium as a substitute, leading to disturbed ion homeostasis and growth inhibition. This mechanism was particularly relevant to our findings with OB, where we observed that iron supplementation could rescue the growth inhibition effect. The interplay between calcium and iron might contribute to OB’s antifungal mechanism in several ways. The iron deficiency caused by OB might lead to calcium overload and increased intracellular reactive oxygen species (ROS) production, further contributing to its antifungal activity. This dual effect on both calcium and iron homeostasis could explain efficacy of OB against C. albicans and suggested potential advantages in combating drug resistance.
In recent study, iron chelator was reported to synergize with antifungal drugs. The fluconazole-resistant C. albicans strain was found enhancing iron uptake to maintain its resistance; however, it was more sensitive to fluconazole while in hosts treated with iron chelators, suggesting that iron availability played an important role in azole resistance [41]. Compared to our results, OB may cause iron depletion, which was similar to the function of iron chelators, and thus disrupted iron availability of C. albicans, resulting in growth defect. In fact, iron chelators could also be antifungal agents that inhibit C. albicans from growth [42–44].
Calcineurin is a calcium and calmodulin-dependent serine/threonine protein phosphatase that plays a crucial role in immune responses [45]. Although OB and calcineurin both play roles in modulating calcium signaling pathways, they affect different tissues and systems. While calcineurin activity is dependent on calcium signaling in immune cells, OB acts by inhibiting calcium channels in smooth muscle cells. Results showed that OB could also inhibit C. albicans calcineurin mutants Δcna1/Δcna1 and Δcnb1/Δcnb1 (Table 1), suggesting specific pathways regulated by calcineurin might not participate in the antifungal mechanism of OB. However, whether there could be pharmacological or physiological interactions if OB and calcineurin inhibitors are used in a clinical setting is not clear, and how calcium dynamics simultaneously affect both smooth muscle contraction and immune responses also needs to be clarified. If OB acts independently, it suggests broader potential for combination therapy or effectiveness.
In conclusion, this study demonstrated that OB, an FDA-approved drug for IBS treatment, exhibited promising antifungal activity against C. albicans in vitro through interference with iron homeostasis. OB effectively inhibited both yeast-to-hyphae transition and biofilm formation, which were two critical virulence factors of C. albicans. While its in vivo efficacy was limited, possibly due to distribution challenges and complex host environments, our findings provided valuable insights for future antifungal drug development. The established safety profile of OB, combined with its novel antifungal mechanism, suggesting that chemical modifications or alternative delivery strategies could potentially enhance its therapeutic efficacy against fungal infections. Further investigation of iron homeostasis as a target for antifungal drug development may also lead to new therapeutic approaches.
Supplementary Material
ARRIVE guidelines checklist 20250902.pdf
Hsu et al_Supplementary_20250609.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Noble SM, Gianetti BA, Witchley JN. Candida albicans cell-type switching and functional plasticity in the mammalian host. Nat Rev Microbiol. 2017;15(2):96–14. doi: 10.1038/nrmicro.2016.15727867199 PMC 5957277 · doi ↗ · pubmed ↗
- 2Horn DL, Neofytos D, Anaissie EJ, et al. Epidemiology and outcomes of candidemia in 2019 patients: data from the prospective antifungal therapy alliance registry. Clin Infect Dis. 2009;48(12):1695–1703. doi: 10.1086/599039 Epub 2009/05/16. Pub Med PMID: 19441981.19441981 · doi ↗ · pubmed ↗
- 3Chang YL, Yu SJ, Heitman J, et al. New facets of antifungal therapy. Virulence. 2017;8(2):222–236. doi: 10.1080/21505594.2016.125745727820668 PMC 5354158 · doi ↗ · pubmed ↗
- 4Ko HT, Hsu LH, Yang SY, et al. Repurposing the thrombopoietin receptor agonist eltrombopag as an anticryptococcal agent. Med Mycol. 2020;58(4):493–504. doi: 10.1093/mmy/myz 077 Epub 2019/07/13. Pub Med PMID: 31297540.31297540 · doi ↗ · pubmed ↗
- 5Mao X, Auer DL, Buchalla W, et al. Cetylpyridinium chloride: mechanism of action, antimicrobial efficacy in biofilms, and potential risks of resistance. Antimicrob Agents Chemother. 2020;64(8):576–520. doi: 10.1128/aac.00576-20PMC 752681032513792 · doi ↗ · pubmed ↗
- 6Dire O, Ahmad A, Duze S, et al. Survival of Candida auris on environmental surface materials and low-level resistance to disinfectant. J Hosp Infect. 2023;137:17–23. doi: 10.1016/j.jhin.2023.04.007 Epub 2023/04/29. Pub Med PMID: 37116661.37116661 · doi ↗ · pubmed ↗
- 7Hsu LH, Kwaśniewska D, Cheng W, et al. Gemini quaternary ammonium compound PMT 12-BF 4 inhibits Candida albicans via regulating iron homeostasis. Sci Rep. 2020;10(1):2911. doi: 10.1038/s 41598-020-59750-532076050 PMC 7031538 · doi ↗ · pubmed ↗
- 8Knauf Gregory A, Cunningham Ashley L, Kazi Misha I, et al. Exploring the antimicrobial action of quaternary amines against Acinetobacter baumannii. m Bio. 2020;9(1):e 02394–17. doi: 10.1128/m Bio.02394-17PMC 580147129437928 · doi ↗ · pubmed ↗