Mechanism of anticancer action of bifidobacterium: Insights from gut microbiota
Hoang Do, Esther Asiamah, Mayanijesu Olorife, Arathi Pillai, Sakshi Patel, Ponniah Selvakumar, Sidhartha D. Ray, Ashakumary Lakshmikuttyamma

TL;DR
This review explores how bifidobacterium, a gut microbe, may help fight cancer through multiple mechanisms like boosting treatments and detoxifying harmful substances.
Contribution
The paper provides a comprehensive overview of bifidobacterium's anticancer mechanisms and its interactions with diet and therapies.
Findings
Bifidobacterium can enhance chemotherapy, immunotherapy, and radiation therapy effectiveness.
It modulates immune pathways and promotes apoptosis through regulation of signaling molecules.
Diet influences bifidobacterium's presence in the gut, affecting its anticancer potential.
Abstract
Bifidobacterium has captured major attention recently because of its health benefits and extensive research highlighting its potential in cancer treatment and prevention. Evidence suggests that bifidobacterium can actively fight against various types of cancer, including those of the colon, lungs, breast, and stomach. Research indicates that several species of bifidobacterium can potentiate the action of chemotherapy, immunotherapy and radiation therapy in battling tumors, and reducing their adverse effects. Bifidobacteria shows its multipronged effect by modulating various immunomodulatory and inflammatory signaling pathways, potentially leading to the suppression of tumor growth. Moreover, different species of bifidobacteria are known to regulate signaling molecules involved in promoting apoptosis. In addition, bifidobacteria have an impact on the regulation of diverse microRNAs. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
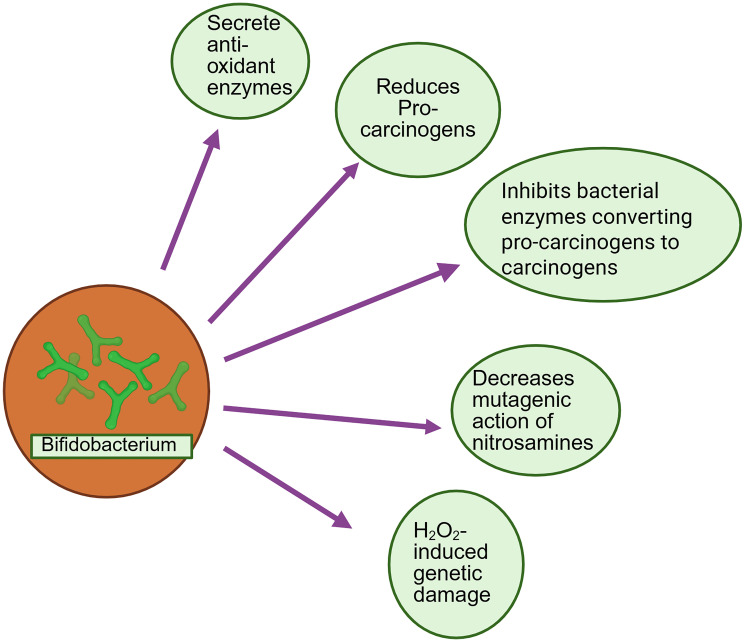 Figure 1
Figure 1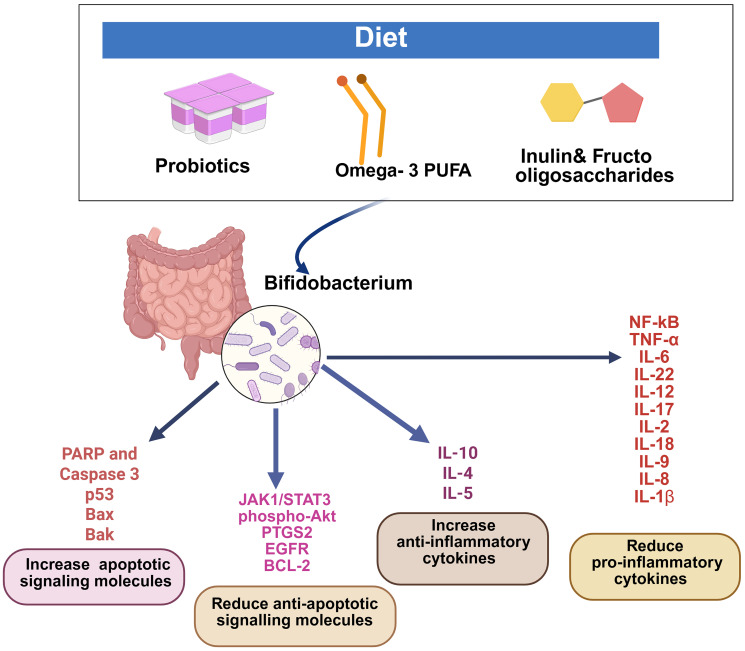 Figure 2
Figure 2| Strain | Sample | Immunological changes | References |
|---|---|---|---|
|
| Lung | ↓TNF-α, IL-6, IL-8 ↑IL-10 | Lu Y, et al., 2023 [ |
|
| Intestine | ↓TLR4 and NF-kB | Dong J, et al., 2022 [ |
|
| Intestine | ↓IFN-γ, IL-2, IL-6, IL-17, IL-22, and IL-12 | Dong J, et al., 2022 [ |
|
| Blood | ↓TNF-α | Klemenak M, et al., 2008 [ |
|
| Colon | ↑IL-10, IL-4 and IL-5 | Fan L, et al., 2021 [ |
|
| Colon | ↓TNFα, IL-6, IL- 1β, IL-18, IL-22, and IL-9 | Fan L, et al., 2021 [ |
|
| Colon | ↓IL-8, MCP-1, and IL-6 | Tomosada, et al. [ |
|
| Caco-2 cells | ↓IL-8, NF-kB | Wang Z, et al., 2011 [ |
|
| Small intestinal epithelial cell | ↓IL-1β and IL-18 | Zhu H, et al., 2021 [ |
| Strain | Cancer types | Apoptotic signaling | References |
|---|---|---|---|
|
| Colon adenocarcinoma cells | ↓PTGS-2, HER-2 and EGFR | Asadollahi P, et al., 2020 [ |
|
| Non-Small Cell Lung Cancer Cell lines | ↑PARP and caspase 3 | An J, et al., 2020 [ |
|
| Hepatocellular carcinoma | ↓IL-6/JAK1/STAT3 ↑GPCR 43 | Song Q, et al., 2023 [ |
|
| Gastric cancer xenograft model | ↓Akt Phosphorylation ↑p53 Bax and Bak, ↑PARP and caspase 3 and 9 | Kim S, et al., 2022 [ |
|
| Head and neck squamous cancer cell lines | ↑Exopolysaccharides | Wang L, et al., 2019 [ |
| Location | Dietary components | References |
|---|---|---|
|
| Pectin-derived oligosaccharides metabolite-Short Chain fatty acids | Bindels LB, et al., 2015, [ |
|
| Rye-supplemented diets (metabolite -Enterolactone) | Oikarinen S, et al., 2003 [ |
|
| Inulin/Oligofructose | Gibson, GR, et al., 1995 [ |
|
| Inulin/Oligofructose | Rao AV, 1999 [ |
|
| Short-chain fructo-oligosaccharides | Bouhnik, Y et al., 1999 [ |
|
| Fructo-oligosaccharides | Bouhnik Y, et al., 1996 [ |
|
| Omega-3 PUFAs | Fu Y, et al., 2021 [ |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Probiotics and Fermented Foods · Cancer Research and Treatments
INTRODUCTION
Bifidobacteria have been found in various environments, including the gastrointestinal tract of different mammals, the oral cavity, and the guts of insects [1, 2]. They were first isolated in 1899 from the feces of breast-fed infants [3]. Bifidobacterium species, promoted by breastfeeding, produces aromatic lactic acids in infants’ guts which is believed to support immune function early in life. A reduction in specific microbes, such as Bifidobacterium, during early life has been associated with a higher risk of developing allergies and asthma in childhood. It has been suggested that depletion of this specific bacterium weaken immune function and increase susceptibility to infectious diseases. [4–6].
Bifidobacterium plays a crucial role in human health by modulating myriads of biochemical and physiological networks within the human body, of which few are known but many remain unknown [7]. Various biological functions of bifidobacteria have been documented, including immune regulation, anti-tumor activity, anti-pathogenic action, anti-inflammation, anti-aging effects, and regulation of hyperlipemia. [7–10]. Dietary polysaccharides or oligosaccharides, primarily the indigestible components, serve as the energy source for bifidobacterium growth [7, 11–12]. Dietary sources profoundly influence bifidobacteria. Some reports suggest that supplementation with pectic oligosaccharides (POS) increases specific populations of gut microbiota in vivo. Bindels et al. found that POS significantly increases Bifidobacterium spp., Roseburia spp., and Bacteroides spp. [13]. Another study suggested that all rye-supplemented diets enhanced the in vivo growth of bifidobacterium compared to non-fiber diet [14]. This study also observed higher levels of plasma enterolactone in the rye-bran group compared to the non-fiber diet group.
ANTICANCER ACTION OF BIFIDOBACTERIUM
Several provocative studies have shown bifidobacteria’s potential to act as an effective anticancer agent, in addition to reducing intestinal and liver disorders, boosting immune responses, and deescalating the rapidity of aging [7, 9, 15–20]. Due to their presence in various organs besides the intestine, their anticancer benefits have been studied in different tissues. The enrichment of Bifidobacterium breve (B. breve) has been reported to inhibit the occurrence and progression of various cancers. [21, 22]. A large volume of literature vouches for the anti-cancer action of bifidobacterium.
Bifidobacterium and lung cancer
Majority of lung cancer patients, approximately 90%, belong to non-small cell lung cancer (NSCLC) group. Healthy subjects and lung cancer patients display differences in their gut microbiota. An enhancement of Bifidobacterium. breve has been identified in healthy subjects compared to those with non-small cell lung cancer (NSCLC) patients [23]. Further studies identified that median progression-free survival was prolonged in non-small-cell lung cancer (NSCLC) patients with significant levels of Bifidobacterium. breve in their gut microbiota [24]. Consistent with these findings, additional studies demonstrated that treatment with aqueous extracts of various bifidobacterium species significantly inhibited cell proliferation and induced apoptosis in different NSCLC cell lines (A549, H1299, and HCC827) [25]. These authors suggested that mechanistically Bifidobacterium bifidum (B. bifidum) treatment reduced cancer cell invasion by downregulating MMP-9 expression. Therefore, it appears likely that use of bifidobacterium could be developed as an adjunctive anticancer treatment option [25]. Moreover, gut microbiota may likely predict the efficacy and adverse effects of immune checkpoint inhibitor therapy. The presence of Bifidobacterium, Escherichia, and Sarterella is associated with higher clinical benefits when anti-PD1 immunotherapy is combined with chemotherapy. Interestingly, presence of Bifidobacterium. breve in the gut microbiota extended the median progression-free survival of patients treated with anti-PD-1 therapy combined with chemotherapy [24]. NSCLC patients with higher levels of Parabacteroides, Clostridia bacterium UC5.1_2F7, and Bifidobacterium dentium (B. dentium) showed a better outcome to checkpoint inhibitor therapy [26]. Another study found that Bifidobacterium. bifidum was also abundant in non-small-cell lung cancer patients responsive to checkpoint inhibitor therapy. Bifidobacterium. bifidum treatment synergistically induced the anti-cancer action of PD-1 antibody or oxaliplatin therapy in a mouse lung cancer model [27]. Yang et. al. showed increased radiosensitivity of lung cancer when the NSCLC patients were treated with Bifidobacterium infantis (B. infantis) [28]. Further clinical studies are warranted to assess the beneficial effect of bifidobacterium with other therapies on the reduction of tumor growth and metastasis.
Bifidobacterium and breast cancer
Breast cancer is one of the most common malignancies affecting women worldwide. Approximately 80% of early-detected, non-metastatic breast cancers are curable. Assessing ER (Estrogen Receptor), PR (Progesterone Receptor), and HER2 (Human Epidermal Growth Factor Receptor 2) are the hallmarks for determining whether a patient is eligible for hormonal or anti-HER2-targeted therapies. The health benefits of probiotics, including bifidobacterium, have been investigated in breast cancer patients (Stage I–III). When these patients received Bifidobacterium. longum (B. longum), Lactobacillus acidophilus, and Enterococcus faecalis in combination with docetaxel-based chemotherapy, considerable reduction in plasma LDL and body weight was observed in probiotics treated patients compared to control group [29]. Another study demonstrated that administering probiotics (Bifidobacterium. longum BB536, Lactobacillus rhamnosus HN001) along with a mediterranean diet led to reduction in body weight, glucose levels, and insulin resistance in breast cancer survivors [30]. Furthermore, a study using breast cancer survivors found that the combination of bifidobacterium and perilla oil decreased the fear of cancer recurrence [31]. Another study showed that combination of Bifidobacterium. longum and ononis hirta methanol-extract treatment was effective in enhancing the immune response with a simultaneous reduction in mammary gland tumors in a mouse model [32].
Dynamic changes of gut microbiota are common in premenopausal and postmenopausal women. A recent study identified that bifidobacterium spp. was specifically decreased in premenopausal breast cancer patients compared to postmenopausal breast cancer patients [33]. Triple negative breast cancer (TNBC) is a subtype where ER, PR, and HER2 are negative. Hormonal and anti-Her2 therapies remain ineffective for TNBC patients. Epidemiological data indicate that TNBC occurs most often in young women in the premenopausal stage [34]. Young TNBC patients frequently exhibit BRCA1 and BRCA2 mutations. Coincidentally, TP53 and RAD50 mutations are also associated with TNBC in young patients [33]. Studies on the impact of gut microbiota in premenopausal breast cancer are limited. Bifidobacterium. infantis 35624 reduced tumor growth in TNBC animal model. Furthermore, Bifidobacterium. infantis 35624 and doxorubicin co-administration enhanced the anticancer potential of doxorubicin [35]. Another study using a BALB/c mouse model also found that sonication-killed Bifidobacterium. bifidum reduced tumor incidence and progression of TNBC. This bacterium also increased p53 levels and, at the same time, reduced Ki-67 expression [36]. Another study reported the physiological properties of gut bacterial membrane vesicles (B-MVs) and their combined action with cancer immunotherapy in a TNBC mouse model [37]. In a mouse study, bifidobacterium-derived B-MVs inhibited the growth of MDA-MB-231-induced triple-negative breast tumors. The antitumor effect of B-MVs is achieved by inducing cancer cell apoptosis through the upregulation of Bax and downregulation of Bcl-2 [37]. Treatment with Bifidobacterium. longum-C-CPE-PE23 (a genetically modified strain of B. longum secreting claudin-4 (CL-4) targeting protein C-CPE-PE23) to a TNBC mouse exhibited higher induction of apoptotic cells and tumor growth inhibition compared to control groups. Systemic toxicity was minimal in experimental animals with Bifidobacterium. longum-C-CPE-PE23 treatment [38].
Currently, options for TNBC treatment are extremely limited, and more carefully designed studies are needed to explore the action of bifidobacterium in enhancing the anticancer action of chemotherapy, radiation therapy and immunotherapy.
Bifidobacterium and colorectal cancer
Colorectal cancer (CRC) is the third leading cause of cancer-associated death in men and fourth cause in women. There is ample evidence available on the correlation between CRC and gut microbiota [21, 39]. The ameliorative role of probiotics, including lactobacillus and Bifidobacterium spp. on CRC has been reported both in vitro and in vivo [40, 41]. One study investigated the presence of a specific fecal bacteria in colorectal cancer patients and healthy subjects. Bifidobacterium adolescentis (B. adolescentis) was found to be significantly lower in CRC patients compared to healthy controls [42]. This suggests the benefit of this bacterial population against CRC. The apoptosis-inducing ability of bifidobacterium has been demonstrated in the colon cancer cell line LS174T compared to IEC-18 normal cells, although actual mechanisms remain unclear. One study used individual and mixed probiotics, including (1) single strain of Lactobacillus. reuteri, (2) B. Breve, (3) mixture of 5 strains of Lactobacilli (LC), (4) 5 strains of bifidobacteria (BC), (5) 10 strains of Lactobacillus and Bifidobacterium (L+B). Among these probiotics, a cocktail of 5 strains of bifidobacterium showed higher apoptosis in LS174T cells. Bifidobacterium decreased the expression levels of EGFR, HER-2, and COX-2. The colon cancer incidence and progression of tumor stage were significantly inhibited by Bifidobacterium in an animal study [21, 39]. In order to establish a correlation between gut microbiota and CRC, another study isolated bifidobacterium from dairy products, infant feces, and probiotic capsules. The cell-free supernatant of isolated bifidobacterium was tested for its cytotoxicity in colon cancer cell lines. Bifidobacterium. bifidum inhibited the growth of colon cancer cell line SW742 cell-line in particular [43]. Bifidobacterium also exhibited regulation of miRNAs and their target genes in an azoxymethane-induced colon cancer mouse model. Treatment of both Lactobacillus acidophilus and Bifidobacterium. bifidum increased the miR-26b and miR-18a expressions, which were decreased in azoxymethane-induced colon tumors. Further, both the bacterial populations elevated the expression of miR-155. KRAS is one of the targets for miR-155, and the probiotics treatment significantly increased KRAS expression level in the colon tumor. Regulation of these miRNAs and target genes provided potential tumor suppressive action in the colon cancer cells by these probiotics [44]. In contrast, a study found that Bifidobacterium. longum (BL) decreased oncomiRs miR-21a and miR-155 but enhanced the tumor suppressor miRNAs (miR-145 and miR-15a) in colorectal cancer mice model [45]. Induction of tumor suppressing miRNAs and inhibition of oncomirs resulted in decreased CRC cell proliferation and invasion [45]. Increased levels of cholesterol, triacylglycerol, and low-density lipoprotein are associated with colorectal cancer. Administration of Lactobacillus acidophilus and Bifidobacterium. bifidum significantly reduced these lipid molecules in azoxymethane-induced colon tumors. Furthermore, these probiotics decreased vitamin D receptor (VDR) and leptin receptor (LPR) gene expression levels in colon tumors. Regulation of VDR, LPR, and lipid parameters play a key role in the development of colon cancer [46]. Another study also demonstrated the effect of bifidobacterium in reducing colorectal cancer tumorigenesis. Administration of Bifidobacterium. bifidum CGMCC 15068 regulated metabolites involved in different pathways such as glycolysis, citrate cycle, fatty acid biosynthesis, butyrate metabolism, and galactose metabolism in a colorectal cancer mouse model. Polysaccharide fractions comprising chiro-inositol, rhamnose, glucose, galactose, and ribose isolated from Bifidobacterium. bifidum BGN4 reduced the proliferation of colon cancer HT-29 and HCT-116 cells [47]. Another bifidobacterium species, Bifidobacterium. adolescentis SPM0212 reduced the growth of colon cancer cell lines, such as, Caco-2, HT-29, and SW480 [48]. Further studies are warranted to explore bifidobacterium’s role in supporting chemotherapy, immunotherapy and radiotherapy.
Gastric cancer and bifidobacterium
Gastric cancer is classified into two subtypes such as diffuse and intestinal. The overall survival rate is better for intestinal gastric cancer subtype compared to diffuse type. A study was carried out to analyze the intestinal microflora closely associated with gastric cancer and found that the gastric cancer patients had significantly lower levels of faecalibacterium, bifidobacterium, and subdoligranulum species in their intestinal tract compared to healthy individuals [49]. Another study showed that probiotics such as bifidobacterium, lactobacillus, and streptococcus species reduced the gastric cancer associated inflammation [50]. Helicobacter pylori infection is one of the prime causes for the development of gastric cancer. There is evidence suggesting that the presence of gut bifidobacterium protected from Helicobacter pylori associated gastric diseases [51]. Nada et al. found that Lactobacillus acidophilus and Bifidobacterium. longum exhibited antiproliferative and anti-angiogenic action against gastric and bladder cancers [52]. More detailed study is required to unravel the beneficiary action of bifidobacterium against gastric cancer.
Bifidobacterium and oral squamous carcinoma
Oral carcinoma can occur in mouth, lips, tongue, and gums. Oral squamous cell carcinoma is a tumor that originates from the oral cavity squamous epithelial cells lining. Very few studies are available associated with bifidobacterium and oral squamous carcinoma. Li et al. reported that Bifidobacterium. Breve, another species of bifidobacterium, inhibited the proliferation of oral squamous carcinoma tumor transplanted in C3H/HeN mice. The antitumor action of bifidobacterium is due to the recruitment of T cells to the tumor microenvironment through the secretion of higher levels of IL-12 from dendritic cells [53]. An in vitro study using head and neck squamous cell carcinoma cell line observed that exopolysaccharides composed of rhamnose (Rha), arabinose (Ara), galactose (Gal), glucose (Glc), and mannose (Man) derived from Bifidobacterium. breve lw01 induced cell cycle arrest and cell apoptosis [54]. Although all these observations significantly testify a positive role for bifidobacteria, carefully designed clinical studies are required to explore the beneficial effect of bifidobacterium against oral squamous carcinoma.
MECHANISM OF ANTI-CANCER ACTION OF BIFIDOBACTERIUM
The anticancer action of bifidobacterium follows a multitude of pathways and mechanisms such as biotransformation, activation of immune function, fermentation of undigested complex polysaccharides, and regulation of various oncogenic and tumor suppressor molecules.
Immune modulation by bifidobacterium
Bifidobacterium exhibits tumor-suppressing effects by enhancing immune function by impacting immune cells such as B lymphocytes, NK cells, and macrophages via a variety of pathways and mechanisms [18, 55, 56]. Bifidobacterium. animalis F1-7 decreased the proliferation of melanoma B16-F10 cells by inhibiting the pro-inflammatory molecules such as IL-6, IL-8 and TNF-α, and activating anti-inflammatory molecule IL-10 [56]. Butanol extract of Bifidobacterium. adolescentis SPM0212 reduced the growth of colon cancer cell lines (Caco-2, HT-29, and SW480) and enhanced the TNF-α release [48]. Another study found that supplementation of Bifidobacterium. adolescentis reduced colorectal tumor formation by increasing CD143+ cancer-associated fibroblasts, increased GAS1 expression and Wnt/β-catenin pathway [42].
Most of the cancer cells have higher NF-kB activity, which produce higher amounts of pro-inflammatory factors and induces cancer cell proliferation. A recent study found that lipopolysaccharide (LPS) induced protein expression of TLR4 and NF-kB p65 was decreased by Bifidobacterium. longum BL-10 group treatment [57]. Furthermore, this study showed that BL-10 treatment balanced the Th1 and Th2 function and reduced the expression levels of different pro-inflammatory cytokines (IFN-γ, IL-2, IL-6, IL-17, IL-22, and IL-12). Conversely, BL-10 enhanced anti-inflammatory cytokine IL-4 levels, which was decreased by LPS treatment [57]. Consumption of probiotic Bifidobacterium. breve BR03 and B632 strains decreased TNF-α levels in children with celiac disease [58]. An in vitro study using Caco-2 cells found that Bifidobacterium. animalis subsp. lactis strain BB12 reduced the TNF-induced production of IL-8 via transcriptional inhibition of NF-kB pathway [59]. Pro-inflammatory cytokines such as TNFα, IL-6, IL- 1β, IL-18, IL-22, and IL-9 were significantly reduced in the colon homogenates of mice treated with Bifidobacterium. adolescents compared to controls, whereas the anti-inflammatory cytokines IL-10, IL-4 and IL-5 were higher [60]. Bifidobacterium. breve also has been shown to enhance IL-10-production (Type 1 regulatory cells) in Tr1 cells in the large intestine [61]. One study using porcine intestinal epithelial cells challenged with heat-killed enterotoxigenic Escherichia coli displayed that Bifidobacterium. longum BB536 and B. breve M-16V strains significantly reduced interleukin (IL)-8, monocyte chemotactic protein (MCP)-1 and IL-6 expression levels [62]. One of our earlier reviews also discussed the action of bifidobacterium in downregulating all proinflammatory cytokines through the downregulation of NF-kB activity [63]. A selenium enriched Bifidobacterium. longum DD98 effectively reduced the intestinal and hepatic toxicity of irinotecan (CPT-11), by reducing the pro-inflammatory cytokines IL-1β and IL-18 [64]. Treatment with Bifidobacterium. breve NCIMB 702258 decreased proinflammatory cytokines TNF-alpha and IFN-gamma levels [65]. Together, these studies demonstrated that different strains of bifidobacterium have the potential to decrease different proinflammatory cytokines and at the same time increase anti-inflammatory cytokines. Summary of the influence of different strains of bifidobacterium on inflammatory cytokines is depicted in Table 1.
Bifidobacterium enhances anti-tumor action of immunotherapy
Although the immune modulating bifidobacterium strains that directly attack cancers are very limited, there are several evidences available on the combination of immunotherapy and bifidobacterium’s action in enhancing the anti-cancer potential of immunotherapy. Immune checkpoint inhibitors act on immune checkpoints such as CTLA-4 and PD-1 to enhance the T cell response towards cancer cells. Recent research observed that immune checkpoint inhibitors anti-tumor action is significantly influenced by gut probiotics [34]. Mager et al. reported that Bifidobacterium. pseudolongum could not exhibit anti-tumor immunity against colon cancer, bladder cancer and melanoma, whereas its combination with anti-CTLA-4 treatment significantly decreases tumor growth compared to anti-CTLA-4 treatment alone by increasing IFN-γ production mediated via higher spleen TH1 cell activation [66]. Furthermore, this study found that the concentration of a microbial metabolite inosine was significantly higher in Bifidobacterium. pseudolongum monocolonized colorectal cancer mice model compared with colidextribacter species monocolonized mice [66]. The authors stated that the enhancement of anti-tumor action of CTLA-4 antibody by Bifidobacterium. pseudolongum was mediated by inosine-A2AR signaling in T cells [66]. A milestone study by Sivan et. al. showed that bifidobacterium spp. enhanced the anticancer action of PD-L1 against melanoma [67]. Another study showed that Bifidobacterium. bifidum significantly increased the anti-tumor immunity by enhancing the anti-cancer action of PD-L1 treatment in a melanoma mouse model [68]. A study by Lee et al. observed that Bifidobacterium. bifidum treatment together with PD-1 antibody significantly inhibited the tumor growth compared to the monotherapy of PD-1 antibody [27]. Bifidobacterium. longum RAPO enhances the anti-tumor immune response of anti-PD-1 therapy in triple negative breast cancer [69].
A very recent study found that Bifidobacterium-derived extracellular vesicles (Bif.BEVs) enhanced anti-cancer action of anti-PD-1 therapy in NSCLC. Both in vivo and in vitro study found that lung cancer cells uptake Bif.BEVs through dynamin-dependent endocytosis pathway. Further, this study demonstrated that Bif.BEVs treatment significantly increased PD-L1 expression in lung cancer cell lines through TLR4-NF-κB pathway [70]. A recent study in liver cancer observed that the combination of bifidobacterium or isobutyrate with anti-PD-1 significantly reduced the tumor size. Further, this study demonstrated that the abundance of bifidobacterium or levels of isobutyrate in the gut microbiota may assist as predictive markers for immunotherapy response in liver cancer patients [71].
Carcinogen detoxification by bifidobacterium
Available reports suggested that bifidobacterium decreases the production of carcinogens by regulating metabolism in the intestinal flora [72]. Bifidobacterium intake decreased the levels of different bacterial enzymes such as nitroreductase, β-glucuronidase, and azoreductase [73, 74]. These bacterial enzymes are responsible for the conversion of procarcinogens to carcinogens [75]. Anti-carcinogenic effects of bifidobacterium may also be due to the elimination of procarcinogens from the intestinal flora [1]. Common carcinogenic compounds that exist in food and the atmosphere are polycyclic aromatic hydrocarbons (PAHs), heterocyclic amines (HCAs) pthalic acid esters (PAEs), and mycotoxins [76]. It has been reported that HCAs were bound with Bifidobacterium. pseudocatenulatum and Bifidobacterium. longum [77]. This may lead to decreases in the absorption of HCA in the intestine and consequently increase the elimination via feces [77]. Bifidobacterium can decrease the mutagenic action of nitrosamines. It has been reported that bifidobacteria can physically bind to nitrosamines such as HCAs [77]. Bifidobacterium. animalis var. lactis Bb12 inhibited the formation of aberrant crypt foci (ACF) by dimethylhydrazine (DMH) in mice. This indicates bifidobacterium protected from carcinogen-induced colonic preneoplastic lesions in mice [78]. Another study found that H_2_O_2_-induced DNA strand breaks and overall genetic damage were protected in colorectal cancer cells (HT29clone19A) by human fecal water, which is collected from probiotics (Lactobacillus acidophilus 145 and Bifidobacterium. longum 913) treated healthy human volunteers [79]. Figure 1 represents the schematic illustration of carcinogen detoxification by bifidobacterium.
Schematic illustration of carcinogen detoxification by bifidobacterium.
A study analyzed the effects of dietary probiotic bacteria Bifidobacterium. lactis (B. lactis), resistant starch (RS), and their interaction on colorectal cancer apoptosis. This study revealed that a combination of dietary RS and Bifidobacterium. lactis significantly enhanced the cancer cell’s apoptotic response to a carcinogen in the colorectal cancer animal model [80]. Bifidobacterium strains were found to decrease cancer cell growth by exhibiting anti-proliferative, pro-apoptotic, and antioxidant attributes. Bifidobacteria was shown to accomplish pro-apoptotic action by increasing pro-caspases and Bax proteins and downregulating the anti-apoptotic Bcl-2 proteins [81].
Bifidobacterium regulates cancer cell apoptosis and signaling
Different reports suggest that bifidobacterium spp. can regulate the genes involved in cell proliferation and apoptosis [81]. Treatment of a cocktail containing five strains of bifidobacterium induced cell death in colon adenocarcinoma cells [82]. Further, bifidobacterium treatment decreased the expression of PTGS-2 (20 folds), HER-2 (6.7 folds), and EGFR (4.4 folds) [21]. Another study suggested that aqueous extracts from Bifidobacterium. bifidum and Bifidobacterium. lactis significantly decreased the proliferation of NSCL cancer cell lines (A549 and H1299). Further, bifidobacterium increased apoptosis, enhanced the levels of cleaved poly ADP-ribose polymerase (PARP) and caspase 3. Bifidobacterium also decreased expression of MMP-9, which led to decreased invasiveness of lung cancer cells [25]. A recent study suggested that Bifidobacterium. pseudolongum reduced non-alcoholic fatty liver disease-associated hepatocellular carcinoma (NAFLD-HCC) in an animal model. Bifidobacterium. pseudolongum significantly reduced cell growth via the reduction in the G1/S transition of cell cycle. Furthermore, this study found that Bifidobacterium. pseudolongum reduced IL-6/JAK1/STAT3 signaling pathway by the activation of G coupled-protein receptor 43 [83]. Bifidobacterium. bifidum reduced tumor growth in the gastric cancer xenograft model. This study further demonstrated that the decreased tumor growth is due to decreased Akt phosphorylation, increased expression of tumor suppressor p53, and apoptotic regulators Bax and Bak proteins. Higher apoptosis in gastric tumor was confirmed by higher expression of cleaved caspase-3 and 9, and PARP [84]. A study provided evidence on the anti-cancer action of exopolysaccharides derived from Bifidobacterium. breve lw01. The authors identified that genes responsible for the exopolysaccharide’s biosynthesis are present in this bacterium’s genome as cluster regions with 14 predicted genes. The exopolysaccharides identified in Bifidobacterium. breve lw01 are rhamnose, arabinose, galactose, glucose, and mannose. Experiments using head and neck squamous cancer cell lines identified that exopolysaccharides inhibited cancer cell proliferation in a concentration dependent manner by powerfully impacting cell cycle arrest and apoptosis regulating machinery [54]. Exopolysaccharides derived from Bifidobacteria enhance the production of cytokines such as TNF-α and IL-12, thereby promoting T cell-mediated apoptosis in colon cancer cells [85]. Summary of different apoptotic signaling molecules regulated by different strains of bifidobacterium are illustrated in Table 2.
Moreover, the metabolites produced by bifidobacterium exhibit anti-cancer action. Supplementation of bifidobacterium via probiotics or prebiotics may increase the levels of blood omega-3 PUFAs. B. breve NCIMB 702258 and linoleic acid-supplemented diet enhanced the omega-3 (n-3) fatty acids, such as, eicosapentaenoic acid and docosahexaenoic acid levels in mice adipose tissue [65]. Studies suggested that PUFAs can induce anti-carcinogenesis action [86]. Furthermore, bifidobacteria produces short chain fatty acids including acetate, butyrate, and propionate [87, 88]. These metabolites produce anti-cancer action against colon cancer [89, 90]. Butyrate is an important metabolite that exhibits both immunomodulatory and anti-inflammatory properties. Furthermore, butyrate acts as a tumor suppressor which induces anti-tumor action in various cancers [91–93].
Bifidobacterium influences biotransformation of dietary components
Anticancer action of bifidobacteria may also be due to the biotransformation of dietary components into anti-cancer molecules. A study using a leukemic mice model showed that dietary pectic oligosaccharides (POS) enhanced the growth of bifidobacteria compared to inulin containing diet. It was also shown that (POS) increased the bifidobacterium species and increased the concentration of short-chain fatty acids (SCFA) such as acetate, propionate, and butyrate [13]. A study analyzed the effect of two types of rye-bran fractions on the population of bifidobacrterium/enterolactone production in intestinal neoplasia animal models. This study demonstrated that rye-bran diet increased the bifidobacterium population, whereas the non-fiber diet significantly lowered intestinal bifidobacterium level [14]. In addition, the same study found that higher levels of intestinal bifidobacterium might be associated with more enterolactone production [14], which likely provides an anti-tumor safeguard against intestinal neoplasia in mice [9]. Several other studies indicated that intake of inulin, a prebiotic fiber increased gut bifidobacterium [94]. A human study showed that higher production of bifidobacteria through the supplementation of oligofructose and inulin-containing diet [95], and another human study showed that the growth of most bifidobacteria was higher with inulin and oligofructose compared to glucose [96]. A recent randomized controlled trial using type 2 diabetes patients showed that, compared to maltodextrin, six weeks of supplementation of a 50/50 mixture of inulin and oligofructose increased fecal bifidobacteria and short-chain fatty acids, acetic acid, and propionic acid [97]. Butyrate has been shown as an effective anti-cancer agent by reducing the level of inflammatory cytokines, and act as an inhibitor to histone deacetylase [98].
It has been established by various studies that fructose oligosaccharides and inulin are effective prebiotics. Short-chain fructo-oligosaccharides consist of polysaccharides containing glucose connected to fructose units (Gfn; n = </= 4). The oligosaccharides are fermented only in the colon and enhance the bifidobacteria growth. The optimal level of fructo-oligosaccharides required for healthy human volunteers to increase fecal bifidobacteria is 10 g/d. These non-digestible dietary components specifically enhance the growth of certain gut microbiota such as bifidobacteria [99]. One other report suggested that intake of 12.5 g/d dietary fructo-oligosaccharides increased colonic bifidobacteria in human volunteers [100]. Fructo-oligosaccharides (FOS) consist of fructose connected by β-(2→1)-glycosidic bonds with a glucose unit. FOS and inulin are present in many dietary sources such as asparagus, onions, garlic, Jerusalem artichokes, and leeks [101]. Inulin, comprise of fructose moieties connected through (2-1)-d-frutosyl fructose bonds, which is mainly isolated from chicory roots [102]. Bifidobacteria spp. can catabolize different kinds of mono-and oligosaccharides. Bifidobacterium can ferment dietary fructo-oligosaccharides and inulin [101, 103]. Bifidobacterium. adolescentis and B. thermophilum fermented both FOS and inulin, Whereas Bifidobacterium. infantis and Bifidobacterium. bifidum fermented only FOS not inulin. β-fructofuranosidase is the common enzyme found in most of the bifidobacterium strains, however, strains that ferment inulin have more hydrolytic activity against fructans. Various gut microbiota showed differences in fermenting carbohydrates by using different pathways. Bifidobacteria in particular use a pathway named “bifid shunt” to degrade hexose sugar. The key enzyme in this pathway is fructose-6-phosphoketolase (EC 4.1.2) [104].
Polyunsaturated fatty acids (PUFA) have a direct impact on the level of different gut microbiota. Similarly, gut microbiota influences the bioavailability, and biotransformation of PUFAs [104]. Different studies suggest that omega-3 PUFAs regulate gut microbiota population, increasing the growth of bifidobacteria [104–106] and decreasing the growth of Enterobacteria [105]. Correspondingly, there is a correlation that exists between probiotic supplementation and fatty acid levels in serum and tissues. Omega-3 polyunsaturated fatty acids (PUFAs) are essential nutrients that display various health benefits. Bifidobacterium has been shown to have a positive association with the level of different PUFAs such as the level of DHA and omega-3 PUFAs in breast cancer survivors [107]. Summary of the different dietary constituents which influence the intestinal/colon/fecal bifidobacterium population is described in Table 3.
Bifidobacteria may also metabolize certain drugs into active anti-cancer molecules [108]. Gut microbiota, such as ruminococcus spp., bacteroides spp., and bifidobacterium spp. are involved in the metabolism of ginseng to ginsenoside Rb1, which has potent anti-cancer action [109]. Another study reported that bifidobacterium sp. metabolized ginsenoside Rb2 to 20-O-beta-D-glucopyranosyl-20(S)-protopanaxadiol (compound K) [110]. Anticancer action of compound K has been observed in different cancers such as acute myeloid leukemia, liver, lung, and nasopharyngeal carcinoma [111]. Lapachol (1,4-naphthoquinone) is naturally present in the bark of the lapacho tree. Bifidobacterium sp. and Lactobacillus acidophilus metabolized lapachol to dehydro-α-lapachone, which showed higher anticancer activity compared to lapachol [112]. Antitumor bioactive molecules formed in soymilk during fermentation by Streptococcus thermophilus 14085 and Bifidobacterium. infantis 14603 displayed higher anti-tumor activity against colorectal cell lines compared to normal soymilk [113].
SUMMARY AND FUTURE PERSPECTIVE
Different health benefits of bifidobacterium have been identified, such as anti-cancer, immune regulation, and anti-inflammatory effects (Figure 2). Anti-cancer action of bifidobacterium has been documented in different tumors and cancer cell types. Bifidobacterium species reduced the growth and metastasis of NSCLC [23, 25]. Presence of Bifidobacterium. breve in gut microbiota extended the median progression-free survival of NSCLC patients. Different studies reported the levels of different species of bifidobacterium provide a prediction on the efficacy and adverse effects of immune checkpoint inhibitor therapy for NSCLC patients [24]. Premenopausal breast cancer patients displayed lower levels of bifidobacterium spp. compared to postmenopausal patients [33] yielding to a possibility that should be investigated. TNBC occurs mostly in premenopausal women, and there are few reports suggesting the anti-cancer action of bifidobacterium against TNBC [35–38]. Additionally, studies demonstrated that Bifidobacterium. infantis increased the anticancer action of doxorubicin for TNBC [35], which should be extrapolated whether this type of combination that can be used to synergize the action of chemotherapeutics yielding to the possibility of minimizing the side effects of the drug that are incurred by increasing the dose of the drug. TNBC is one of the hard-to-treat cancer compared to other types of breast cancers. Chemotherapy is one of the major treatment options for TNBC, and majority of patients develop resistance, and it leads to tumor metastasis. Hence it will be worthwhile to explore the efficacy of bifidobacterium in supporting the chemotherapy action in TNBC patients.
Bifidobacterium regulates inflammatory cytokines and apoptotic signals.
The pro-apoptotic action of bifidobacterium was established in colon cancer cell lines and in animal models [40–42]. Bifidobacterium reduced the level of different colorectal cancer, oncogenic signaling molecules such as EGFR, COX-2, miR-21a and miR-155 [21], and simultaneously enhancing the tumor suppressor miR-145 and miR-15a [45]. A fascinating mechanism of anti-cancer action of bifidobacterium is mediated through the immune regulation primarily targeting proinflammatory cytokines [57, 60, 64]. Available studies have demonstrated that the presence of bifidobacterium considerably decreased pro-inflammatory cytokines such as TNF- α and IL-6, IL-18, and IL- 1β, whereas the anti-inflammatory cytokines such as IL-10, IL-4, and IL-5 increased appreciably (Figure 2). Most of these changes are detected in intestinal and/or/colonic tissues. Moreover, studies available in various cancer models are limited; therefore, results of these studies should be interpreted with caution. Hence, further studies are required to evaluate the effect of bifidobacterium species on specific immune regulators in different types of cancer. Different stains of bifidobacterium decrease the levels of different bacterial enzymes, which are responsible for the conversion of dietary procarcinogens to carcinogens [73, 75]. It has also been suggested that different signaling molecules involved in cancer cell proliferation decrease [25, 81]. One of the other mechanisms is the biotransformation of different dietary components into anti-cancer molecules. Various studies demonstrated bifidobacterium growth is influenced by specific diets such as inulin, oligofructose [94–96], and PUFA [105–108] (Figure 2). Initiating studies on cancer prevention through a high fiber diet mediated by enriched gut bifidobacterium is crucial.
With context to complement the benefit of bifidobacterium in enhancing the action of immunotherapy, radiation therapy and chemotherapy, more studies are needed to identify anti-cancer specificity of different bifidobacterium species. Recent studies suggest that bifidobacterium increases the anti-cancer action of immune check point inhibitors (anti-PD1 and anti-CTLA4) for various cancers [66–71]. It is well documented that chemotherapy and radiation treatment decrease the intestinal microflora including bifidobacterium [113–119] and increases pathogenic family members of enterobacteriaceae [120, 121]. When designing in vivo studies related to the combination of bifidobacterium and chemotherapy, it’s imperative to evaluate the presence of intestinal bifidobacterium.
In summary, the presence of bifidobacterium in the intestinal flora can be heavily influenced by the intake of probiotics as well as a diet rich in fibers like inulin, pectin, oligofructose, and other components such as PUFA. Bifidobacterium’s anticancer effects are multifaceted, involving the modulation of immune responses through the regulation of both pro-inflammatory and anti-inflammatory cytokines, influencing apoptotic pathways, and neutralizing carcinogens introduced into the body via diet and miscellaneous other environmental exposures. Consequently, bifidobacterium has the potential to complement conventional chemotherapeutic regimens, immunotherapy, and radiotherapy. Nonetheless, further research is necessary to fully understand the specific roles of individual bifidobacterium strains and their impact on different types of cancer, particularly in terms of cancer prevention, enhancing standard treatments and reducing tumor proliferation and metastasis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1O’Callaghan A , van Sinderen D . Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front Microbiol. 2016; 7:925. 10.3389/fmicb.2016.00925. 27379055 PMC 4908950 · doi ↗ · pubmed ↗
- 2Klijn A , Mercenier A , Arigoni F . Lessons from the genomes of bifidobacteria. FEMS Microbiol Rev. 2005; 29:491–509. 10.1016/j.femsre.2005.04.010. 15939502 · doi ↗ · pubmed ↗
- 3Laursen MF , Sakanaka M , von Burg N , Mörbe U , Andersen D , Moll JM , Pekmez CT , Rivollier A , Michaelsen KF , Mølgaard C , Lind MV , Dragsted LO , Katayama T , et al. Bifidobacterium species associated with breastfeeding produce aromatic lactic acids in the infant gut. Nat Microbiol. 2021; 6:1367–82. 10.1038/s 41564-021-00970-4. 34675385 PMC 8556157 · doi ↗ · pubmed ↗
- 4Hevia A , Milani C , López P , Donado CD , Cuervo A , González S , Suárez A , Turroni F , Gueimonde M , Ventura M , Sánchez B , Margolles A . Allergic Patients with Long-Term Asthma Display Low Levels of Bifidobacterium adolescentis. P Lo S One. 2016; 11:e 0147809. 10.1371/journal.pone.0147809. 26840903 PMC 4739579 · doi ↗ · pubmed ↗
- 5Wang W , Luo X , Zhang Q , He X , Zhang Z , Wang X . Bifidobacterium infantis Relieves Allergic Asthma in Mice by Regulating Th 1/Th 2. Med Sci Monit. 2020; 26:e 920583. 10.12659/MSM.920583. 32249275 PMC 7160606 · doi ↗ · pubmed ↗
- 6Arboleya S , Watkins C , Stanton C , Ross RP . Gut Bifidobacteria Populations in Human Health and Aging. Front Microbiol. 2016; 7:1204. 10.3389/fmicb.2016.01204. 27594848 PMC 4990546 · doi ↗ · pubmed ↗
- 7Luo J , Li Y , Xie J , Gao L , Liu L , Ou S , Chen L , Peng X . The primary biological network of Bifidobacterium in the gut. FEMS Microbiol Lett. 2018; 365. 10.1093/femsle/fny 057. 29546277 · doi ↗ · pubmed ↗
- 8Lim HJ , Shin HS . Antimicrobial and Immunomodulatory Effects of Bifidobacterium Strains: A Review. J Microbiol Biotechnol. 2020; 30:1793–800. 10.4014/jmb.2007.07046. 33144551 PMC 9728261 · doi ↗ · pubmed ↗