Transcriptional Regulation in Roots by Bacteria With 1‐Aminocyclopropane‐1‐Carboxylate Deaminase Enzymes for Drought Tolerance and Post‐Stress Recovery
William Errickson, Bingru Huang

TL;DR
This study explores how bacteria with ACC deaminase enzymes help plants tolerate drought and recover after stress by regulating gene expression in roots.
Contribution
The study identifies specific genes and pathways regulated by ACCd bacteria in creeping bentgrass under drought and recovery.
Findings
ACCd bacteria caused differential expression of 53 genes under drought stress, including SUMO protease OTS1 and antioxidant genes.
During post-drought recovery, 160 genes were differentially expressed, including DNA repair and root development genes.
Carotenoid biosynthesis pathways were up-regulated in inoculated plants during recovery.
Abstract
Plant growth‐promoting rhizobacteria (PGPR) that can break down 1‐aminocyclopropane‐1‐carboxylate (ACC), an ethylene precursor, by ACC deaminase enzymes (ACCd) to reduce ethylene production in plants may enhance plant tolerance to drought stress. This study aimed to identify genes in plant roots regulated by ACCd‐bacteria under drought stress and re‐watering and to determine major molecular factors and associated metabolic pathways for ACCd bacteria‐enhanced drought tolerance and post‐stress recovery in creeping bentgrass ( Agrostis stolonifera ). Transcriptomic analysis was performed in root tissues from plants inoculated with a novel strain of ACCd‐producing bacteria, Paraburkholderia aspalathi “WSF23,” under well‐watered conditions, 35 days of drought stress, and 15 days of re‐watering. ACCd bacteria inoculation resulted in differential expression of 53 genes under drought stress.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
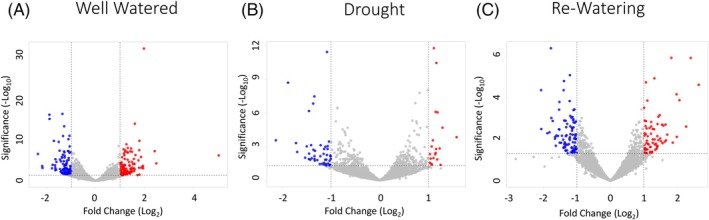 FIGURE 1
FIGURE 1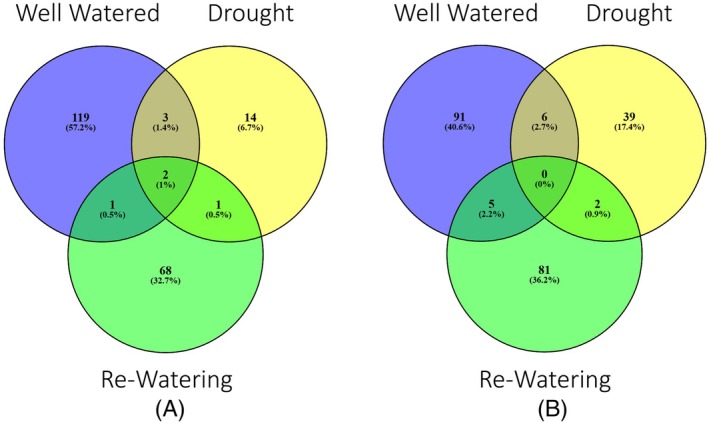 FIGURE 2
FIGURE 2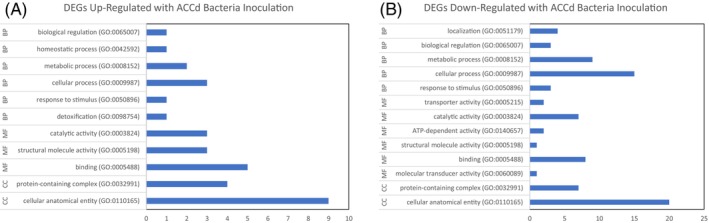 FIGURE 3
FIGURE 3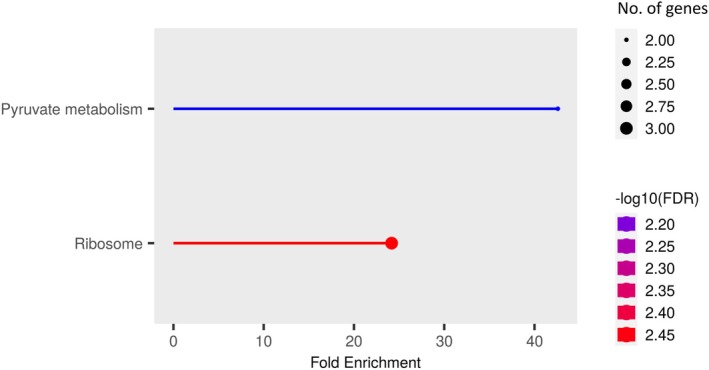 FIGURE 4
FIGURE 4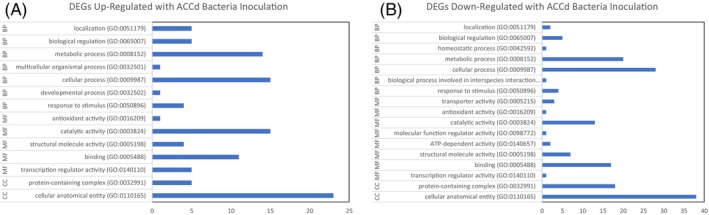 FIGURE 5
FIGURE 5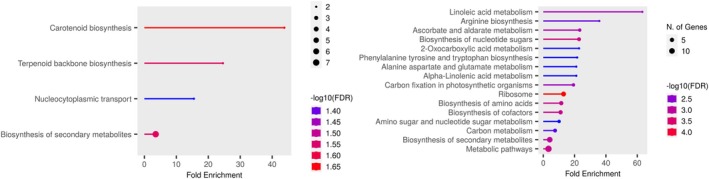 FIGURE 6
FIGURE 6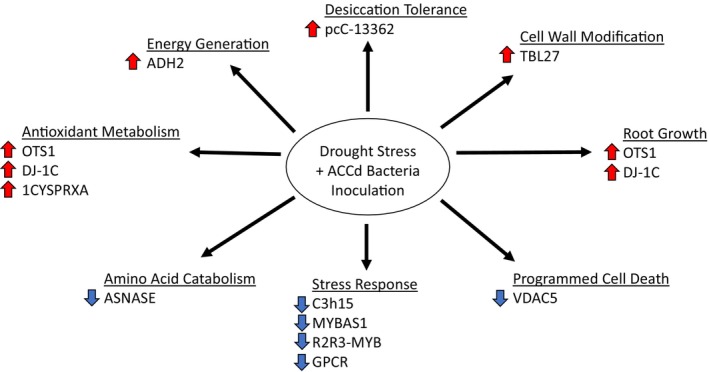 FIGURE 7
FIGURE 7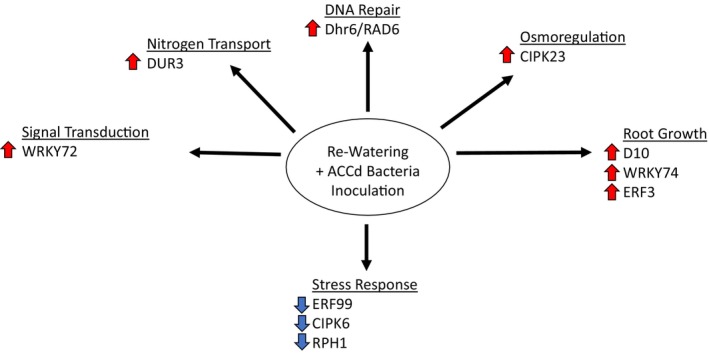 FIGURE 8
FIGURE 8| Up‐regulated | Down regulated | ||||
|---|---|---|---|---|---|
| Accession no. | FC (log2) | Gene name | Accession no. | FC (log2) | Gene name |
| Os04g0671700 | 1.18 | DJ‐1C | Os03g0202200 | −1.69 | VDAC5 |
| Os04g0404400 | 1.15 | PCC13‐62 | Os01g0363900 | −1.53 | CRD1 |
| Os05g0426300 | 1.10 | TBL27 | Os02g0301000 | −1.50 | C3H15 |
| Os01g0243200 | 1.07 | Os_F0797 | Os03g0597600 | −1.45 | ASNASE1 |
| Os03g0356414 | 1.06 | DIS1 | Os12g0207500 | −1.45 | ATPASE3 |
| Os07g0638300 | 1.03 | 1CYSPRXA | Os01g0533100 | −1.39 | GSL9 |
| Os03g0254400 | 1.02 | pPLAIIIalpha | Os01g0533000 | −1.37 | GSL9 |
| Os05g0141400 | 1.29 | Unknown | Os11g0700500 | −1.34 | MYBAS1 |
| Os03g0133875 | 1.25 | Unknown | Os06g0677000 | −1.34 | DEP3 |
| Os11g0246900 | 1.18 | Unknown | Os02g0180000 | −1.23 | PP2C11 |
| Os08g0558100 | 1.14 | Unknown | Os06g0199800 | −1.18 | GPCR |
| Os05g0477300 | 1.09 | Unknown | Os09g0563250 | −1.14 | DHHC23 |
| Os03g0807000 | 1.06 | Unknown | Os02g0639000 | −1.12 | LMS |
| Os01g0925200 | 1.04 | Unknown | Os07g0111600 | −1.11 | PAP9A |
| Os09g0346400 | −1.10 | BIP103 | |||
| Os06g0186400 | −1.10 | SCP32 | |||
| Os02g0655500 | −1.08 | OsSTA68 | |||
| Os03g0586900 | −1.07 | PLS2 | |||
| Os02g0181900 | −1.01 | CLPB‐M | |||
| Os06g0653800 | −1.01 | DWD71 | |||
| Os02g0738600 | −1.01 | GH9B8 | |||
| Os02g0800000 | −1.88 | Unknown | |||
| Os10g0574800 | −1.72 | Unknown | |||
| Os08g0562300 | −1.45 | Unknown | |||
| Os02g0580100 | −1.43 | Unknown | |||
| Os02g0778400 | −1.41 | Unknown | |||
| Os06g0106100 | −1.34 | Unknown | |||
| Os07g0191700 | −1.33 | Unknown | |||
| Os11g0195100 | −1.33 | Unknown | |||
| Os03g0645200 | −1.28 | Unknown | |||
| Os01g0858500 | −1.14 | Unknown | |||
| Os01g0760400 | −1.14 | Unknown | |||
| Os01g0812050 | −1.12 | Unknown | |||
| Os05g0306000 | −1.06 | Unknown | |||
| Os03g0769000 | −1.04 | Unknown | |||
| Os03g0170100 | −1.03 | Unknown | |||
| Os12g0615800 | −1.03 | Unknown | |||
| Os01g0793500 | −1.01 | Unknown | |||
| Os03g0388500 | −1.00 | Unknown | |||
| Up‐regulated | Down‐regulated | ||||
|---|---|---|---|---|---|
| Accession no. | FC (log2) | Gene name | Accession no. | FC (log2) | Gene name |
| Os04g0667800 | 2.41 | Dhr6 | Os01g0868000 | −2.07 | ERF99 |
| Os01g0746400 | 2.07 | D10 | Os09g0468700 | −1.89 | OsbHLH046 |
| Os03g0292100 | 1.83 | PP2C32 | Os09g0437400 | −1.82 | SAUR38 |
| Os02g0806700 | 1.67 | RING104 | Os08g0441100 | −1.73 | CIPK06 |
| Os02g0652000 | 1.65 | CYCU4 | Os01g0281600 | −1.66 | ENODL4 |
| Os11g0490900 | 1.55 | WRKY72 | Os10g0560700 | −1.57 | ERF36 |
| Os05g0494600 | 1.49 | Os_F0757 | Os02g0758800 | −1.54 | UCL6 |
| Os10g0580400 | 1.44 | DUR3 | Os05g0277000 | −1.52 | EXPA33 |
| Os09g0334500 | 1.44 | WRKY74 | Os05g0468800 | −1.45 | CRP |
| Os01g0797600 | 1.41 | ERF3 | Os04g0581100 | −1.39 | S3H |
| Os01g0802100 | 1.41 | GRY340 | Os03g0826500 | −1.38 | ASA1 |
| Os07g0659700 | 1.38 | FBX477 | Os07g0105000 | −1.37 | UCL20 |
| Os04g0655300 | 1.36 | RLCK168 | Os06g0682900 | −1.36 | HSA32 |
| Os02g0681200 | 1.33 | RING327 | Os05g0230600 | −1.34 | RFC3 |
| Os02g0115700 | 1.33 | CATA | Os01g0875300 | −1.34 | USP7 |
| Os03g0820300 | 1.32 | ZFP182 | Os02g0699700 | −1.33 | TOP2 |
| Os04g0556000 | 1.31 | HMA5 | Os08g0253800 | −1.32 | CSLC3 |
| Os04g0578400 | 1.27 | BCH2, HYD1 | Os08g0327400 | −1.31 | ENR1 |
| Os10g0399200 | 1.27 | CGS1 | Os05g0572000 | −1.30 | RPH1 |
| Os01g0147200 | 1.25 | ASD1 | Os03g0682100 | −1.29 | MFS1 |
| Os07g0190000 | 1.22 | DXS3 | Os01g0644200 | −1.28 | SALP1 |
| Os03g0838800 | 1.22 | HINGE3 | Os04g0473400 | −1.28 | RPL6 |
| Os02g0681700 | 1.22 | RLCK83 | Os03g0297100 | −1.24 | RPS7A |
| Os02g0282900 | 1.20 | ABCE1 | Os01g0273100 | −1.22 | UBC27 |
| Os03g0242900 | 1.19 | SUB28 | Os10g0565150 | −1.22 | BMY6 |
| Os05g0556100 | 1.15 | DRP1A | Os11g0591100 | −1.21 | GME2 |
| Os03g0744650 | 1.12 | WD40‐90 | Os04g0531100 | −1.21 | RPP16 |
| Os07g0225300 | 1.11 | NAC3 | Os03g0822700 | −1.20 | FES1A |
| Os05g0530400 | 1.09 | SPL7 | Os05g0399300 | −1.20 | CHT2 |
| Os01g0667600 | 1.09 | RAB11C1 | Os10g0417600 | −1.17 | GME1 |
| Os08g0118000 | 1.08 | AHL1 | Os12g0582800 | −1.16 | OSCA2.4 |
| Os03g0349200 | 1.08 | OsCDK2 | Os06g0561000 | −1.15 | MIOX |
| Os04g0645100 | 1.07 | FLO2 | Os01g0675100 | −1.14 | PRXIIC |
| Os03g0741100 | 1.07 | BHLH148 | Os01g0801500 | −1.13 | GNS7 |
| Os07g0150700 | 1.06 | CIPK23 | Os02g0797400 | −1.13 | MCM5 |
| Os02g0114000 | 1.05 | BRM | Os01g0905700 | −1.13 | RING204 |
| Os05g0354400 | 1.04 | XOAT5 | Os01g0152900 | −1.10 | H2B.7 |
| Os03g0197800 | 1.04 | IDD1 | Os08g0509100 | −1.09 | LOX8 |
| Os04g0444200 | 1.04 | CAM | Os01g0622300 | −1.08 | HEME |
| Os09g0566550 | 1.03 | CTR1 | Os05g0437100 | −1.07 | ERF105 |
| Os06g0192800 | 1.03 | ATL69 | Os05g0274200 | −1.07 | MSH2 |
| Os01g0202500 | 1.02 | BBX1 | Os01g0153300 | −1.06 | H2B.5 |
| Os05g0426200 | 1.02 | NAC36 | Os03g0606200 | −1.06 | RMtATP6 |
| Os01g0194000 | 2.27 | Unknown | Os12g0133050 | −1.06 | OsP0C |
| Os04g0667850 | 1.99 | Unknown | Os01g0668100 | −1.05 | FLA7 |
| Os07g0164800 | 1.87 | Unknown | Os10g0390500 | −1.04 | ALAAT |
| Os08g0427900 | 1.70 | Unknown | Os04g0441800 | −1.02 | PTR |
| Os04g0421800 | 1.63 | Unknown | Os08g0508800 | −1.02 | LOX2 |
| Os03g0263900 | 1.59 | Unknown | Os05g0418100 | −1.01 | MLO_ |
| Os01g0570500 | 1.56 | Unknown | Os03g0721900 | −2.07 | Unknown |
| Os08g0540300 | 1.52 | Unknown | Os01g0205500 | −1.78 | Unknown |
| Os02g0177800 | 1.41 | Unknown | Os08g0112566 | −1.77 | Unknown |
| Os08g0109000 | 1.40 | Unknown | Os03g0162200 | −1.65 | Unknown |
| Os07g0569166 | 1.39 | Unknown | Os03g0685500 | −1.63 | Unknown |
| Os03g0349000 | 1.34 | Unknown | Os01g0839300 | −1.61 | Unknown |
| Os01g0148050 | 1.27 | Unknown | Os04g0635500 | −1.53 | Unknown |
| Os04g0408600 | 1.25 | Unknown | Os01g0969100 | −1.47 | Unknown |
| Os02g0802400 | 1.19 | Unknown | Os08g0151400 | −1.41 | Unknown |
| Os02g0626532 | 1.19 | Unknown | Os08g0490800 | −1.41 | Unknown |
| Os05g0183900 | 1.17 | Unknown | Os01g0502900 | −1.38 | Unknown |
| Os02g0177900 | 1.12 | Unknown | Os05g0555800 | −1.34 | Unknown |
| Os04g0565200 | 1.11 | Unknown | Os02g0189800 | −1.34 | Unknown |
| Os01g0764900 | 1.09 | Unknown | Os05g0394200 | −1.33 | Unknown |
| Os01g0708600 | 1.08 | Unknown | Os08g0130550 | −1.22 | Unknown |
| Os12g0163700 | 1.07 | Unknown | Os02g0236000 | −1.20 | Unknown |
| Os01g0558850 | 1.06 | Unknown | Os07g0656800 | −1.19 | Unknown |
| Os03g0219300 | 1.03 | Unknown | Os12g0481400 | −1.16 | Unknown |
| Os06g0591600 | 1.03 | Unknown | Os10g0418000 | −1.15 | Unknown |
| Os01g0152700 | −1.05 | Unknown | |||
| Os05g0466600 | −1.03 | Unknown | |||
| Os03g0628900 | −1.00 | Unknown | |||
- —Center for Turfgrass Science New Jersey Agricultural Experiment Station Rutgers, The State University of New Jersey 59 Dudley Rd. New Brunswick, NJ 08901‐8520
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant responses to water stress · Polymer-Based Agricultural Enhancements
Introduction
1
Abiotic stress, such as drought, causes an increase in ethylene production in plant tissues, which has been associated with the inhibition of root growth and induction of premature leaf senescence (Singh et al. 2015; Dubois et al. 2018; Naing et al. 2021; Chen et al. 2022). Strategies that can suppress ethylene production have shown evidence of improving drought tolerance. Inoculation of plants with different species of plant growth‐promoting rhizobacteria (PGPR), including Paraburkholderia and Bacillus , has resulted in the promotion of drought tolerance through diverse biological functions or modes of action (Glick 2014; Errickson and Huang 2019; Naing et al. 2021). PGPR that produce deaminase enzymes have demonstrated efficacy in improving drought stress tolerance, as ACC deaminase enzymes (ACCd) function to break down 1‐aminocyclopropane‐1‐carboxylate (ACC), a precursor of ethylene, into ammonia and α‐keto butyrate before ACC is converted into ethylene (Glick 2005). This can mitigate the impacts of stress‐induced ethylene production, including premature leaf senescence and reduced root growth. For example, ACCd bacteria, Paraburkholderia aspalathi , enhanced tiller production, root growth, whole‐plant tolerance to drought stress, and post‐stress recovery by suppressing drought‐induced ethylene production in creeping bentgrass ( Agrostis stolonifera L.) (Errickson et al. 2023).
Drought‐tolerant and drought‐susceptible genotypes of a given plant species often exhibit differential gene expression related to drought tolerance, suggesting mechanisms by which the drought‐tolerant genotypes achieve this level of improved tolerance (Tan et al. 2022). Novel approaches to stimulate stress resistance in turf have included exogenous applications of compounds, such as proline, gamma‐aminobutyric acid (GABA), and nitric oxide (NO), to stimulate stress‐defensive gene expression and antioxidant activity (Jiang et al. 2023; Jiang 2023). PGPR inoculation may also induce changes in gene expression that contribute to improved drought tolerance, as PGPR can exhibit other plant growth‐promoting traits in addition to ACCd activity, including stimulation of other hormones, antioxidant activity, and nutrient mobilization (Lata et al. 2024). The promotion of drought tolerance in cool‐season turfgrass addresses a primary objective of sustainable turfgrass management through the reduction of irrigation requirements and subsequent water use (Wang, Olsen, et al. 2023). Symbiotic interactions between plants and endophytic bacteria may represent novel strategies to improve plant growth while reducing water use (White et al. 2021; Chang et al. 2023). While the influence of beneficial microbial endophytes on improving abiotic stress tolerance in cool‐season grass species has been documented, there remains limited understanding of the mechanisms involved, especially considering the multitude of species of beneficial endophytes that can colonize cool‐season grasses (Hewitt et al. 2022; Roberts 2022).
Recent studies in genetic and metabolic changes in turfgrass species in response to abiotic stress have started to provide insights into the molecular basis of stress tolerance (Brown et al. 2023). Metabolomic analysis of leaves and roots of creeping bentgrass colonized by P. aspalathi demonstrated that improved drought tolerance and post‐stress recovery were associated with several mechanisms. Leaf tissue of inoculated plants demonstrated up‐regulation of carbohydrate, pyrimidine, and cytokinin hormone metabolism. Inoculation enhanced the accumulation of metabolites in pathways related to carbohydrate metabolism, organic acid metabolism during respiration, DNA and protein synthesis, and cellular metabolism in roots (Errickson and Huang 2023). While several physiological and metabolic studies have demonstrated ACCd bacteria's abilities to confer abiotic stress resistance, including drought in various plant species (Stearns et al. 2012; Saikia et al. 2018; Naing et al. 2021; Shahid et al. 2024), the mechanisms underlying ACCd PGPR‐enhanced drought tolerance, particularly post‐stress recovery at the molecular level, are poorly understood.
Creeping bentgrass is a cool‐season turfgrass species that is widely used on golf course putting greens and fairways. Strategies for improving abiotic stress tolerance in creeping bentgrass can help to maintain turf quality while reducing irrigation requirements, thus conserving natural resources and improving efficiency and economics for turfgrass managers. Creeping bentgrass is a stoloniferous perennial grass species that can tolerate varying degrees of drought stress and re‐grows upon re‐watering, making it a suitable model turfgrass species for conducting this investigation.
The objectives of this study were to understand which key genes and associated metabolic pathways to promote drought stress tolerance and post‐stress recovery in creeping bentgrass are regulated by ACCd bacteria P. aspalathi “WSF23,” a novel PGPR strain that was isolated from the roots of native grasses in the New Jersey Pine Barrens and demonstrated high levels of ACCd activity. This is a further research advancement from our understanding of physiological and metabolic effects of P. aspalathi on creeping bentgrass tolerance to drought stress and post‐stress recovery that have been reported in previous publications (Errickson et al. 2023; Errickson and Huang 2023). A comprehensive understanding of the mechanisms involved in ACCd bacteria‐mediated stress tolerance is essential for the development of PGPR as broadly applicable biostimulants and their widespread adoption for crop production, especially as it relates to reducing water use while maintaining crop quality.
Materials and Methods
2
Plant Growing Conditions
2.1
Creeping bentgrass (cv. “Penncross”) plants were established from vegetative sod plugs collected at the Rutgers Horticultural Research Farm II in North Brunswick, NJ, USA (40.46918734608342, −74.42450570308384). The sod plugs were divided into individual tillers, and the shoots and roots were trimmed. The tillers were soaked in 1% sodium hypochlorite solution for surface sterilization and then rinsed in autoclaved deionized water. Bunches of 10 tillers were planted into plastic containers (20 cm wide × 30 cm long × 20 cm deep) filled with calcined clay (Profile Products, Buffalo Grove, IL) that had been autoclaved for a 20‐min wet cycle to minimize contamination from other microorganisms. The containers were dispersed across four different controlled environment growth chambers (Environmental Growth Chambers, Chagrin Falls, OH) after a 30‐day establishment period in the greenhouse. The growth chamber conditions were controlled at 23/16°C (day/night temperature), 650 μmol m^−2^ s^−1^ photosynthetically active radiation of 12‐h photoperiod and 50% relative humidity. The plants were watered every 2 days and fertilized every 7 days with half‐strength Hoagland's nutrient solution (Hoagland and Arnon 1950) and were allowed to acclimate to these conditions for 7 days before the imposition of drought stress treatments.
Culture of Bacterial Inoculum and Inoculation of Plants
2.2
A novel PGPR strain of P. aspalathi was used in this study due to its high levels of ACCd activity and previous evidence of improving drought tolerance in creeping bentgrass (Errickson et al. 2023; Errickson and Huang 2023). Bacterial cultures of P. aspalathi “WSF23” were revived from frozen stock vials that had been stored at −80°C. The bacteria were cultured in Luria Broth (Miller) (Sigma–Aldrich) on a shaker set to 120 rpm at 23°C for 4 days. The bacterial suspensions were then centrifuged at 10,000 g for 5 min at 23°C, then resuspended in deionized water. The samples were centrifuged and resuspended twice to remove all Luria Broth from the preparations. The bacterial suspension in deionized water was then adjusted to an OD_600_ value of 1.0.
The bacterial inoculum was watered into the root zone of the plants at a rate of 75 mL per plant. A second dose of the inoculum was applied 24 h later. The inoculation treatments were applied to eight containers that each held six plants that were comprised of 10 individual tillers. A second set of eight containers that held six plants each was treated as a non‐inoculated control group and received only deionized water that did not contain the ACCd bacteria.
Drought Stress and Re‐Watering
2.3
Drought stress began 7 days after plants were first inoculated. Drought stress was applied by withholding all water for 35 days. After 35 days of drought stress, the plants were then re‐watered for 15 days for a post‐drought recovery. While the drought‐stressed plants all showed signs of desiccation after the drought stress period, all plants in every treatment group survived and resumed growth during the re‐watering period. Four containers, each of inoculated and non‐inoculated treatments, were subjected to drought stress and re‐watering treatments. Four containers of inoculated plants and four containers of non‐inoculated plants also received full irrigation and served as a non‐stress control group. The experiment was conducted as a completely randomized design with ACCd bacteria inoculation and drought stress and re‐watering treatments, each repeated in four containers.
Transcriptome Analysis
2.4
Root tissue was destructively sampled for RNA extraction from inoculated and non‐inoculated plants from the three different stress treatment groups (well‐watered, 35 days drought stress, 15 days re‐watering). All plants remained in the vegetative stage during the study, and all samples were vegetative. Three replicates were used for each treatment group. The roots were rinsed free of calcined clay with deionized water, and 0.25 g of fresh tissue was flash‐frozen with liquid nitrogen and ground in a sterile mortar and pestle before being transferred to a new 2‐mL microcentrifuge tube. RNA was isolated from the ground samples following the manufacturer's protocol for the QIAGEN RNEasy kit (Qiagen). Samples were sent to GeneWiz (Genewiz) for transcriptome analysis using RNA‐Seq (Wang et al. 2009). The rice ( Oryza sativa L.) genome was used as a reference because it is similar to the creeping bentgrass genome; however, it is much more complete due to significantly more genomic research being conducted on rice compared to creeping bentgrass.
Statistical Analysis, Gene Classification, and Metabolic Pathway Analysis
2.5
Differentially expressed genes (DEGs) were analyzed using DESeq2 (Love et al. 2014), with a threshold of 1.0 log2 fold change and p < 0.05. Individual genes were researched using the Rice Annotation Project Database (RAP‐DB) (Sakai et al. 2013) and Oryzabase (Kurata and Yamazaki 2006). Volcano plots for the different treatment groups were created using VolcaNoseR (Goedhart and Luijsterburg 2020). Functional classification of gene ontology was conducted using Panther 19.0 (Mi et al. 2019), and KEGG pathway analysis (Kanehisa et al. 2016) was conducted for up‐regulated and down‐regulated DEGs in each treatment group. Venny (Oliveros 2007) was used to create Venn diagrams and compare DEGs for each treatment group.
Results
3
DEG Regulated by ACCd Bacteria in Roots of Plants Exposed to Drought Stress
3.1
ACCd bacteria resulted in up‐regulation or down‐regulation of a total of 53 (DEGs) in roots of plants inoculated with the bacteria relative to the non‐inoculated plants under drought stress (Figure 1). Among the up‐regulated differentially‐expressed genes (DEGs) by ACCd bacteria inoculation, 14 genes were found only under drought stress. Inoculation also resulted in three DEGs that were up‐regulated in both drought and well‐watered conditions, two DEGs that were up‐regulated in drought, well‐watered, and re‐watering conditions, and one DEG that was up‐regulated in all three watering treatments (Figure 2A, Table 1, and Table S1). ACCd bacteria caused the down‐regulation of 39 unique DEGs under drought stress. Six DEGs were down‐regulated under both drought and well‐watered conditions, and two DEGs were down‐regulated during both drought and re‐watering (Figure 2B, Table 1, and Table S1).
Differentially expressed genes in P. aspalathi “WSF23” inoculated creeping bentgrass roots under well‐watered (A), drought (B), and post‐drought recovery (C) conditions, relative to non‐inoculated creeping bentgrass roots (p ≤ 0.05).
Up‐regulated (A) and down‐regulated (B) genes in P. aspalathi “WSF23” inoculated creeping bentgrass roots under well‐watered, drought, and post‐drought recovery conditions, relative to non‐inoculated creeping bentgrass roots (p ≤ 0.05).
The GO Enrichment Analysis demonstrated that up‐regulated DEGs by ACCd bacteria under drought stress included biological processes (biological regulation, homeostatic process, metabolic process, and detoxification), molecular functions (catalytic activity, structural molecule activity, and binding), and cellular components (protein containing complex and cellular anatomical entity) (Figure 3A). Down‐regulated DEGs influenced biological processes (localization, biological regulation, metabolic and cellular processes, and response to stimulus), molecular functions (transporter activity, catalytic activity, ATP‐dependent activity, structural molecular activity, and binding), and cellular components (protein containing complex and cellular anatomical entity) (Figure 3B).
Gene ontology of up‐regulated (A) and down‐regulated (B) genes in P. aspalathi “WSF23” inoculated creeping bentgrass roots under drought stress conditions, relative to non‐inoculated creeping bentgrass roots (p ≤ 0.05).
KEGG Pathway Analysis of the network of gene products, mostly proteins, indicated that the ACCd bacteria up‐regulated genes in drought‐stressed plants that were mainly involved in pyruvate metabolism and ribosome biogenesis (Figure 4).
KEGG Pathways associated with up‐regulated DEGs in P. aspalathi “WSF23” inoculated creeping bentgrass roots under drought stress conditions, relative to non‐inoculated creeping bentgrass roots (p ≤ 0.05).
Differentially Expressed Genes (DEG) Regulated by ACCd Bacteria in Roots of Plants Upon Re‐Watering
3.2
Creeping bentgrass roots inoculated with ACCd bacteria demonstrated 160 DEGs after 15 days of re‐watering compared to non‐inoculated plants. There was a total of 72 up‐regulated DEGs and 88 down‐regulated DEGs (Figure 1). Sixty‐eight DEGs were uniquely up‐regulated in inoculated plants upon re‐watering, while one DEG was up‐regulated in both well‐watered and re‐watered treatment groups, in addition to the up‐regulated DEGs in common with drought stress treatments listed above (Figure 2A, Table 2, and Table S2). Eighty‐one DEGs were uniquely down‐regulated in inoculated plants after re‐watering, while five down‐regulated DEGs were common to both re‐watering and well‐watered conditions, in addition to the two down‐regulated DEGs shared between re‐watering and drought stress conditions (Figure 2B, Table 2, and Table S2).
The GO Enrichment Analysis indicated DEGs up‐regulated by ACCd bacteria during re‐watering influenced the regulation of biological processes including localization, biological regulation, metabolic process, multicellular organismal process, cellular process, developmental process, and response to stimulus, as well as molecular functions including antioxidant activity, catalytic activity, structural molecule activity, binding, and transcription regulator activity, and the cellular components for protein‐containing complex and cellular anatomical entity (Figure 5A). DEGs down‐regulated by ACCd bacteria inoculation during re‐watering influenced the biological processes of localization, biological regulation, homeostatic process, and metabolic and cellular processes. Biological processes involved in interspecies interactions between organisms, and response to stimulus, as well as molecular functions of transporter activity, antioxidant activity, catalytic activity, molecular function regulator activity, ATP‐dependent activity, structural molecular activity, binding, and transcription regulator activity, and cellular components of protein‐containing complex, and cellular anatomical entity were also affected (Figure 5B).
Gene ontology of up‐regulated (A) and down‐regulated (B) genes in P. aspalathi “WSF23” inoculated creeping bentgrass roots upon re‐watering, relative to non‐inoculated creeping bentgrass roots (p ≤ 0.05).
The KEGG Pathway Analysis revealed that DEGs up‐regulated by ACCd bacteria during re‐watering mainly influenced carotenoid biosynthesis, terpenoid backbone biosynthesis, nucleocytoplasmic transport, and biosynthesis of secondary metabolites. DEGs down‐regulated by inoculation affected linoleic acid metabolism, arginine biosynthesis, ascorbate and aldarate metabolism, and biosynthesis of nucleotide sugars (Figure 6).
KEGG Pathways associated with up‐regulated (A) and down‐regulated (B) DEGs in P. aspalathi “WSF23” inoculated creeping bentgrass roots upon re‐watering, relative to non‐inoculated creeping bentgrass roots (p ≤ 0.05).
Discussion
4
Improved drought tolerance by ACCd bacteria has been associated with improvement in physiological and metabolic activities (Saikia et al. 2018; Naing et al. 2021; Errickson et al. 2023; Errickson and Huang 2023; Shahid et al. 2024). This study found that improved plant tolerance to drought stress and post‐stress recovery by the ACCd bacteria, P. aspalathi “WSF23,” was associated with differential transcriptional regulation in the roots of creeping bentgrass under drought stress and upon re‐watering. Specific genes regulated by P. aspalathi “WSF23” under drought stress and re‐watering conditions are summarized in the pathway maps (Figures 7 and 8). Selected genes with known functions regulated by P. aspalathi “WSF23” under drought stress or during re‐watering that may be related to stress tolerance and recovery are discussed below.
Summary of genes and associated metabolic pathways regulated by P. aspalathi “WSF23” contributing to improved drought tolerance. The upward arrows indicate up‐regulated genes by the inoculation. The downward arrows indicate down‐regulated genes by the inoculation.
Summary of genes and associated metabolic pathways regulated by P. aspalathi “WSF23” contributing to improved post‐stress recovery. The upward arrows indicate up‐regulated genes by the inoculation. The downward arrows indicate down‐regulated genes by the inoculation.
ACCd Bacteria Regulated Genes and Associated Cellular and Metabolic Pathways for Improving Drought Tolerance
4.1
Among all the up‐regulated genes, OVERLY TOLERANT TO SALT 1 (OTS1) demonstrated the highest fold‐change (by 1.6 FC) in plants inoculated with P. aspalathi “WSF23” exposed to drought stress. OTS1 codes for SUMO (Small Ubiquitin‐Like Modifier) protease, which controls filament elongation and cell expansion or elongation and plays key roles in root development and seed germination, as well as plant tolerance to abiotic stress, including drought and salt stress (Srivastava et al. 2016). Overexpression of OTS1 in sugarcane ( Saccharum spp. hybrids), which is another perennial grass species, similarly resulted in improved drought tolerance, with plants demonstrating higher stomatal conductance, chlorophyll content, and photosynthesis rate, as well as lower protease activity in OTS1 overexpression plants subjected to drought stress (Masoabi et al. 2023). Overexpression of OTS1 in winter wheat ( Triticum aestivum ) enhanced root growth, chlorophyll content, and antioxidant activity under drought stress conditions (Le Roux et al. 2019). Overexpressing OTS1 in rice increased root length and fresh weight in plants exposed to salt stress (Srivastava et al. 2016). The alcohol dehydrogenase gene, ADH2, was the second‐highest up‐regulated gene in inoculated roots during drought stress (by 1.2 FC). ADH2 codes for alcohol dehydrogenase, which oxidizes alcohols to generate energy for cellular activities and detoxifies cellular damage from the accumulation of alcohol under anaerobic conditions. ADH2 may be induced by soil dry down during drought stress and has conferred abiotic stress resistance in various plant species (Shi et al. 2017; Wang et al. 2024; Zhang et al. 2024). The increased transcript levels of OTS1 and ADH2 following ACCd bacteria inoculation suggest that stress defense systems are activated to protect against cellular damage from dehydration and alcohol to maintain root growth and whole‐plant tolerance to drought stress (Errickson et al. 2023).
P. aspalathi “WSF23” inoculation led to significant upregulation of DJ‐1C under drought stress. This gene belongs to the DJ‐1 superfamily of nuclear gene‐encoding proteins (Lin et al. 2011). DJ‐1C has diverse biological functions, including plastid development and protection against oxidative stress, as well as stress responses (Lin et al. 2011; Wang, Shao, et al. 2023). DJ‐1 proteins have also been shown to contribute to reducing oxidative stress in Arabidopsis by interacting with superoxide dismutase 1 (SOD1) and glutathione peroxidase 2 (GPX2) (Xu et al. 2010; Lin et al. 2011). The improvement of drought and salinity stress tolerance in rice associated with the positive expression of DJ‐1C has been attributed to enhancing root growth, photosynthesis, antioxidant activity, and redox homeostasis (Rathore et al. 2024). Up‐regulation of another gene, 1CYSPRXA, by P. aspalathi “WSF23” inoculation also likely contributed to a reduction in ROS. This gene codes for a peroxiredoxin, which is an antioxidant that reduces alkyl hydroperoxides to alcohols and hydrogen peroxide to water, protecting lipids, enzymes, and DNA from oxidative damage (Lee et al. 2000; Haslekas et al. 2003; Kim et al. 2011). Our results, combined with others, suggest that DJ‐1C and 1CYSPRXA could play important roles in ACCd bacteria‐regulated drought tolerance relating to oxidative stress defenses induced by drought stress, among other effects.
Desiccation‐related protein (DRP) gene, pcC‐13362, was also up‐regulated in inoculated roots subjected to drought stress. The LEA‐like DRP pcC‐13362 has been found to accumulate abundantly in tissues of highly desiccation‐tolerant species such as the resurrection plant raterostigma plantagineum and its close relative Lindera brevidens, while only accumulating to lower levels in the desiccation‐sensitive species Lindera subracemosa (Giarola et al. 2018). The DRP pcC‐13362 has also been identified as a key protein in drought‐tolerant cultivars of the bean ( Phaseolus vulgaris L.) (Pérez et al. 2024) and is up‐regulated in response to pathogen infection (Piatkowski et al. 1990). TBL27 was also up‐regulated (by 1.1 FC) by inoculation during drought stress. This gene is involved in the acetylation of xyloglucan in cell walls, which is important for the structure and elasticity of cell walls (Gille et al. 2011; Zhu et al. 2014; Daher et al. 2024), which contributes to improved desiccation tolerance (Moore et al. 2008). The up‐regulation of pcC‐13362 and TBL27 by P. aspalathi “WSF23” inoculation may lead to increased drought tolerance in creeping bentgrass, involving the maintenance of cell wall elasticity, which plays a critical role in protecting cells from dehydration during drought stress.
The KEGG Pathway Analysis indicated that pyruvate metabolism was the most positively enriched metabolic pathway related to up‐regulated DEGs in inoculated plants after 35 days of drought stress. Pyruvate's role in cellular energy metabolism (Fink 2007; McCommis and Finck 2015) has been well known, and recent studies have further demonstrated the role of pyruvate in promoting abiotic stress tolerance through acting as an antioxidant, encouraging expression of antioxidant enzymes, and up‐regulating stress‐responsive genes and transcription factors during abiotic stress (Alam et al. 2025). Furthermore, KEGG analysis also demonstrated that DEGs related to ribosome biosynthesis were highly up‐regulated in drought‐stressed plants due to ACCd bacteria inoculation. Ribosome biogenesis is responsible for the generation of ribosomes for protein synthesis in cells, which play positive roles in plant tolerance to drought and salt stress (Shiraku et al. 2021). These results suggest that P. aspalathi “WSF23” inoculation could enhance root growth and improve drought tolerance of creeping bentgrass (Errickson et al. 2023) through up‐regulating genes for sustained energy and protein metabolism, along with enhanced stress defense via the enrichment of the pyruvate metabolism pathway and ribosome biogenesis.
ACCd bacteria inoculation also resulted in the down‐regulation of some DEGs under drought stress, including some transcriptional factors or genes involved in stress responses (C3H15 by −1.5 FC, MYBAS1 by −1.3 FC, R2R3‐MYB by −1.2 FC, and GPCR by −1.2 FC) or stress‐induced programmed cell death (VDAC5 by −1.69 FC). C3H15 codes for a CCCH zinc‐finger protease. Previous RT‐qPCR studies have found that these CCCH zinc‐finger protease genes are induced by drought stress and ABA levels in plants under stress (Han et al. 2021). MYBAS1 and R2R3‐MYB are involved in ABA‐mediated leaf senescence (Guo et al. 2017) and have been associated with reduced biomass (Fávero Peixoto‐Junior et al. 2018). MYBs exhibited lower expression in a drought‐tolerant rice genotype relative to that in a drought‐susceptible genotype (Nawae et al. 2020). GPCR, encoding a G‐coupled protein receptor, was also down‐regulated in inoculated roots under drought stress conditions relative to non‐inoculated controls. GPCRS are membrane proteins that are important for signal transduction, with G proteins being induced in response to ABA and other abiotic stresses (Nitta et al. 2015). This corresponds to our previous work, in which ABA levels were lower in creeping bentgrass inoculated with P. aspalathi compared to non‐inoculated controls (Errickson et al. 2023), suggesting that stress‐responsive genes induced by ABA may be down‐regulated as a result of inoculation. VDAC5 was also down‐regulated and is a member of the VDAC family, which comprises voltage‐dependent anion channels located on the membrane of the mitochondria. VDACs have been implicated in programmed cell death induced by abiotic stress, including drought (Godbole et al. 2003, 2013; Desai et al. 2006; Homblé et al. 2012). These results suggest that plants colonized by P. aspalathi “WSF23” may experience a lesser extent of drought stress damage as demonstrated by the down‐regulation of stress‐responsive transcriptional factors or genes associated with stress‐induced cell death, while genes associated with stress protection were up‐regulated.
P. aspalathi “WSF23” inoculation also resulted in the down‐regulation of genes associated with amino acid catabolism, such as ASNASE1 (by −1.45 FC). ASNASE1, coding for asparaginase, was down‐regulated with P. aspalathi inoculation under drought stress. Reduced asparaginase activity also correlates with the increased asparagine levels observed in drought‐stressed creeping bentgrass inoculated with P. aspalathi “WSF23” (Errickson and Huang 2023). Our results indicated that the suppression of asparagine catabolism by P. aspalathi inoculation could contribute to the maintenance of this important amino acid to support plant growth under drought stress for creeping bentgrass.
ACCd Bacteria Regulated Genes and Associated Cellular and Metabolic Pathways for Improving Post‐Drought Recovery Upon Re‐Watering
4.2
Our previous studies found that P. aspalathi “WSF23” inoculation resulted in more rapid growth recovery of creeping bentgrass from drought stress upon re‐watering (Errickson et al. 2023). In this study, several DEGs associated with plant growth were more highly expressed in plants inoculated with P. aspalathi “WSF23” upon re‐watering, following the drought stress period. Dhr6/RAD6, which encodes a ubiquitin‐conjugating enzyme associated with DNA repair and proliferation of new growth (Yamamoto et al. 2004), was up‐regulated by 2.4 FC, and D10, encoding a carotenoid cleavage dioxygenase and supporting strigolactone (SL) biosynthesis, was up‐regulated by 2.1 FC. D10 is involved in the regulation of lateral growth, including tillering and lateral root growth, with localization in the parenchyma of root cells in the stele and xylem parenchyma in the stem. D10 has been shown to stimulate SL biosynthesis, influencing root architecture (Kumar et al. 2015; Sun et al. 2015) and cytokinin biosynthesis that affects tiller production and leaf senescence (Zhang et al. 2010; Ding et al. 2024). These findings are consistent with previous work that has demonstrated increased cytokinin levels and enhanced tiller production in creeping bentgrass inoculated with P. aspalathi “WSF23,” which may ultimately contribute to a more rapid recovery upon re‐watering (Errickson et al. 2023). Carotenoid biosynthesis was also identified by KEGG analysis involving up‐regulated DEGs following 15 days of re‐watering. Among the many roles that carotenoids play in plant growth and development, carotenoids and their derivatives have been shown to affect root architecture and adaptation to the rhizosphere environment, including initiation of lateral roots (Ke et al. 2022), further supporting previous evidence of improved root growth with P. aspalathi inoculation (Errickson et al. 2023).
Several WRKY genes were also upregulated during re‐watering, including WRKY72 (by 1.55 FC) and WRKY74 (by 1.4 FC). WRKY72 can be induced by JA, ABA, and bacterial infection and plays a role in abiotic stress signaling (Ashwini et al. 2016). WRKY74 plays a positive role in plant tolerance to low inorganic phosphate (Pi) stress and root growth under low Pi conditions, and is inducible by bacterial inoculation (Dai et al. 2016). Improved post‐stress recovery in creeping bentgrass inoculated with P. aspalathi “WSF23” could also have resulted from up‐regulation of DUR3 (by 1.4 FC), which is a urea transporter that plays a role in root uptake of urea (Wang et al. 2012; Beier et al. 2019). ERF3 (+1.3 FC) was also up‐regulated in inoculated roots during re‐watering. ERF3 is essential for crown root development and functions in auxin and cytokinin‐responsive gene expression and signaling (Zhao et al. 2015). CIPK23 (+1.1 FC) encodes a CBL‐interacting protein kinase that activates K^+^ transporters to facilitate ion homeostasis and increased K^+^ uptake, which may have also contributed to increased root growth upon re‐watering and enhanced osmoregulation, with K^+^ serving as a compatible solute (Ragel et al. 2015). Accumulation of the compatible solute, proline, has also been associated with P. aspalathi inoculation (Errickson and Huang 2023).
In response to re‐watering, several stress‐responsive genes were down‐regulated by ACCd bacteria inoculation of roots. ERF99, an ethylene response factor that is induced by abiotic stress (Balfagón et al. 2020), was down‐regulated by 2.1 FC in inoculated plants during re‐watering. This may be a result of P. aspalathi “WSF23” limiting ethylene production to reduce the expression of stress‐responsive ERFs, as reductions in ethylene have been previously reported (Errickson et al. 2023). Another stress‐inducible DEG, CIPK6, was also down‐regulated (1.7 FC) in inoculated plants. CIPK6 is expressed during drought and salinity stress, and with increased ABA concentrations (Chen et al. 2013), which is consistent with our previous findings of reduced ABA in inoculated plants (Errickson et al. 2023). RPH1 is a transcriptional repressor that regulates stress‐responsive genes and was down‐regulated (1.3 FC) in inoculated plants. Oxidative stress and DNA damage induce phosphorylation of Rph1, which leads to activation of downstream stress‐activated genes (Liang et al. 2013). The downregulation of RPH1 is further evidence of reduced oxidative stress or DNA damage in plants inoculated with P. aspalathi “WSF23.” KEGG analysis identified linoleic acid metabolism as the pathway most affected by ACCd bacteria inoculation, involving down‐regulated DEGs in this pathway during re‐watering. Linoleic acid plays a diverse range of roles in regulating plant growth and development. It can be converted into signaling molecules like jasmonic acid, which is involved in multiple signaling pathways for plant defense responses against stress (Savchenko et al. 2014). The reduced expression of DEGs associated with linoleic acid metabolism may be indicative of inoculated plants rapidly changing from stress defense mode to regrowth upon re‐watering.
Conclusions
5
ACCd‐bacteria, P. aspalathi “WSF23,” improved drought tolerance and post‐stress recovery in creeping bentgrass could be associated with transcriptional regulation in the root system. The up‐regulation of genes affecting antioxidant metabolism (OTS1, DJ‐1C, 1CYSPRXA), energy generation (ADH2), desiccation tolerance (pcC‐13362), cell wall modification (TBL27), and root growth (OTS1, DJ‐1C), as well as pathways involved in pyruvate metabolism and ribosome biogenesis, could contribute to improved drought tolerance. ACCd‐bacteria promoted post‐stress recovery could be attributed to the up‐regulation of genes regulating DNA repair (RAD6), root development (D10, WRKY74, ERF3), nitrogen transport (DUR3), signal transduction (WRKY72), and osmoregulation (CIPK23), as well as pathways involved in carotenoid biosynthesis. These findings suggest novel mechanisms by which the ACCd bacteria P. aspalathi can promote drought stress tolerance and post‐drought recovery in cool‐season grass species, which may contribute to sustainable methods of reducing water use in turfgrass management. Future work may investigate gene–gene and gene‐protein interactions in cool‐season turfgrass inoculated with ACCd bacteria to further develop a mechanistic understanding of bacteria‐mediated improvement of drought tolerance.
Author Contributions
W.E. conducted the experiment, data analysis, and wrote the manuscript. B.H. developed the research ideas, experimental design, acquired funding, and revised the manuscript.
Funding
This work was supported by Center for Turfgrass Science New Jersey Agricultural Experiment Station Rutgers, The State University of New Jersey 59 Dudley Rd. New Brunswick, NJ 08901‐8520.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Table S1: Differentially expressed genes resultant from inoculation found in drought stressed plants and across other irrigation treatments relative to non‐inoculated roots (p ≤ 0.05). Table S2: Differentially expressed genes resultant from inoculation found after re‐watering plants following 35 days of drought stress that were also differentially expressed across other irrigation treatments relative to non‐inoculated roots (p ≤ 0.05).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alam, N. B. , S. Pelzang , A. Jain , and A. Mustafiz . 2025. “Cytoprotective Role of Pyruvate in Mitigating Abiotic Stress Response in Arabidopsis Thaliana.” Plant Science 352: 1–22. 10.1016/j.plantsci.2024.112325.39608574 · doi ↗ · pubmed ↗
- 2Ashwini, N. , R. S. Sajeevan , M. Udayakumar , and K. N. Nataraja . 2016. “Identification and Characterization of Os WRKY 72 Variant in Indica Genotypes.” Rice Science 23, no. 6: 297–305. 10.1016/j.rsci.2016.07.002. · doi ↗
- 3Balfagón, D. , S. I. Zandalinas , R. Mittler , and A. Gómez‐Cadenas . 2020. “High Temperatures Modify Plant Responses to Abiotic Stress Conditions.” Physiologia Plantarum 170, no. 3: 335–344. 10.1111/ppl.13151.32533896 · doi ↗ · pubmed ↗
- 4Beier, M. P. , T. Fujita , K. Sasaki , et al. 2019. “The Urea Transporter DUR 3 Contributes to Rice Production Under Nitrogen‐Deficient and Field Conditions.” Physiologia Plantarum 167, no. 1: 75–89. 10.1111/ppl.12872.30426495 · doi ↗ · pubmed ↗
- 5Brown, J. M. , C. E. Weldt , H. M. P. Holloway , et al. 2023. “Transcriptomic Analysis of Zoysiagrass ( Zoysia japonica ) Provides Novel Insights Into the Molecular Basis of Cold Acclimation.” Grass Research 3, no. 1: 9. 10.48130/GR-2023-0025. · doi ↗
- 6Chang, X. , B. Young , N. Vaccaro , et al. 2023. “Endophyte Symbiosis: Evolutionary Development, and Impacts of Plant Agriculture.” Grass Research 3, no. 1: 8. 10.48130/GR-2023-0018. · doi ↗
- 7Chen, H. , D. A. Bullock , J. M. Alonso , and A. N. Stepanova . 2022. “To Fight or to Grow: The Balancing Role of Ethylene in Plant Abiotic Stress Responses.” Plants (Basel) 11, no. 1: 33. 10.3390/plants 11010033.PMC 874712235009037 · doi ↗ · pubmed ↗
- 8Chen, L. , Q.‐Q. Wang , L. Zhou , F. Ren , D.‐D. Li , and X.‐B. Li . 2013. “Arabidopsis CBL‐Interacting Protein Kinase (CIPK 6) is Involved in Plant Response to Salt/Osmotic Stress and ABA.” Molecular Biology Reports 40: 4759–4767. 10.1007/s 11033-013-2572-9.23649767 · doi ↗ · pubmed ↗