Loss of the distal-proximal GLP-1/Notch activation gradient in the aging C. elegans germline
Rustelle Janse van Vuuren, Pier-Olivier Martel, Patrick Narbonne

TL;DR
This study shows that a key signaling gradient in the germline of C. elegans declines with age, which may affect stem cell maintenance.
Contribution
The study introduces the use of a genetically encoded sensor to visualize Notch activity in the aging germline in vivo.
Findings
A strong distal-proximal Notch activation gradient is present in young C. elegans adults.
This gradient is largely lost during aging, indicating reduced niche signaling.
The loss correlates with earlier observations of decreased Notch target gene transcription.
Abstract
In C. elegans hermaphrodites, the distal tip cells (DTCs) capping the gonad arms provide a Notch ligand, the niche signal that maintains germline stem cell pools. Using fixed germlines, it was recently shown that the transcription of a Notch target gene decreased relatively early-on during adulthood. Here, we used the genetically encoded Notch Sensor Able to detect Lateral Signaling Activity (SALSA) to examine the pattern of GLP-1/Notch activity across the aging distal gonad in vivo . Interestingly, we find that the robust and progressively decreasing distal-proximal Notch activation gradient that is observed in young adults gets largely lost during aging.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
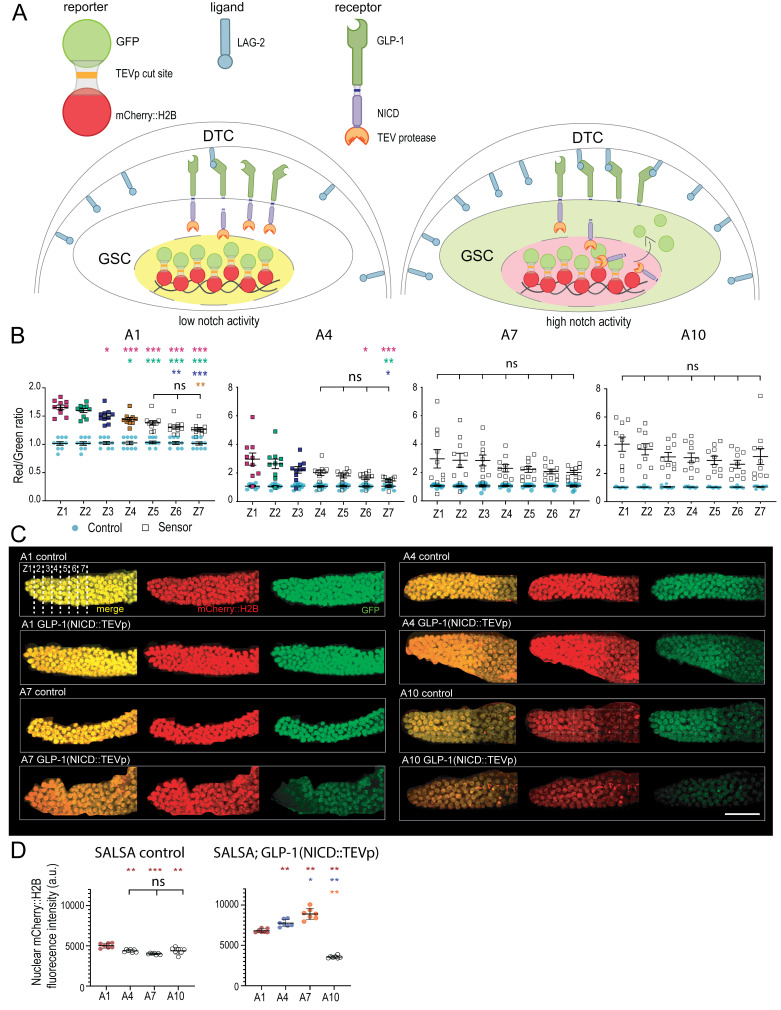 Figure 1
Figure 1|
|
|
|
|
GS9317 |
|
(Shaffer and Greenwald, 2022) |
|
GS9447 |
|
(Shaffer and Greenwald, 2022) |
|
|
Standard
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Developmental Biology and Gene Regulation · Planarian Biology and Electrostimulation
Description
The stem cells' local microenvironment is essential to maintain their stemness. This microenvironment usually consists of other cells or tissues that immediately surround stem cells and provide them with differentiation-preventing and/or self-renewal-promoting “niche” signals. An example is the activation of a Wnt signal in intestinal stem cells by neighbouring enteroendocrine niche cells, something needed for maintaining their proliferation (Fevr et al., 2007). Although the stem cell microenvironment is known to deteriorate with age, the repercussions within stem cells remain largely unascertained.
In * C. elegans * hermaphrodites, a pool of germline stem cells (GSCs) is located distally within each of two gonad arms. Each GSC pool is maintained by a Notch signal expressed by a single niche cell, the distal tip cell (DTC), which caps the germline and extends processes enwrapping GSCs (Austin and Kimble, 1987; Crittenden et al. 2006). The Notch ligand, LAG-2 , present at the surface of the DTC and its processes, activates the GLP-1 /Notch receptor on the membranes of adjacent GSCs (Henderson et al., 1997). Upon receptor activation, the Notch intracellular domain (NICD) is cleaved and translocates to the nucleus where it brings a transcriptional activation complex together (Kopan and Ilagan, 2009). This complex activates two main target genes, * lst-1 * and * sygl-1 * , in a decreasing distal-proximal gradient, to prevent GSC differentiation (Kershner et al., 2014; Lee et al., 2016). As germ cells move proximally due to the ongoing proliferation, they progressively lose Notch stimulation, differentiate and eventually mature into gametes.
A functional decline in germline function occurs with age such that progeny production rapidly decreases in older hermaphrodites, even when sperm supplies are not limited (Hughes et al., 2007). Age-associated alterations in oocyte quality contribute to this reduction in progeny production (Luo et al., 2009). By the 8 ^th^ day of adulthood, germline organization is severely disrupted as the germline diameter drops and cellular debris and large vacuoles accumulate proximally (Hughes et al., 2011). Part of this functional decline may result from defects in GSC maintenance. Indeed, the progenitor zone (PZ) size progressively decreases with age, going down from 220 cells in young adults to 120 cells in day 5 adults (Kocsisova et al, 2019). A decline in stem cell cycling rates, as governed by reductions in insulin/IGF-1 and MPK-1 /ERK signalling levels, may contribute to the decreased PZ size and progeny production (Michaelson et al., 2010; Narbonne et al., 2015; Narbonne et al., 2017; Kocsisova et al, 2019; Robinson-Thiewes et al. 2021). However, the mechanisms underlying the PZ size decline are incompletely understood. Using single-molecule fluorescence in situ hybridization (smFISH) targeting a known transcriptional Notch target gene on fixed germlines isolated from aging hermaphrodites, it was established that distal Notch transcriptional activity significantly decreases relatively early-on during adulthood, but not dramatically (Urman et al., 2024). The initially sharp distal-proximal GLP-1 /Notch activation gradient nonetheless becomes flatter as the DTCs tend to drift proximally with age, resulting in a larger proximal pool of GSCs having equivalent Notch activity (Urman et al., 2024). The aging worm however undergoes global transcriptional changes, resulting in a decline in transcriptional fidelity (Debès et al., 2023), while mRNA turnover may also be impacted (Borbolis and Syntichaki, 2015). Whether the distal-proximal decreasing gradient of GLP-1 /Notch activity persists in older adults in vivo remained unclear.
To better characterize the age-related decline in niche signalling, we examined the pattern of GLP-1 /Notch activity across the aging distal germline in vivo , using the genetically encoded Notch sensor able to detect lateral signalling activity (SALSA) (Shaffer and Greenwald, 2022). The SALSA biosensor consists of a germline-expressed fusion protein made up of GFP, linked via a tobacco-Etch virus protease (TEVp)-cleavable site to a chromatin-tethered RFP (mCherry::H2B). A TEVp is also added to the endogenous Notch intracellular domain (NICD), such that upon receptor activation, NICD::TEVp cleavage and nuclear translocation, GFP is released from the nucleus, resulting in an increased nuclear RFP-to-GFP ratio ( Fig. 1A ). A strain containing only the fluorescent fusion protein is used as a control to determine the basal RFP/GFP ratio under each condition.
Using this sensor, we measured GLP-1 /Notch activation across the PZ, up to 14 cell diameters from the distal tip, in up to 10-day-old adult (A10) hermaphrodites, which corresponds approximately to 2/3 of the typical adult lifespan (Herndon et al., 2002) and is well past the end of the reproductive period (Luo et al., 2009). To this end, germ cells were binned together in seven 2-cell diameter rows from the distal tip, termed zones Z1-Z7. As expected, a robust distal-proximal decreasing gradient of Notch activity was clearly observable in young day 1 adults ( Fig. 1B- C). This gradient was however progressively lost during aging as we failed to detect significant differences between zones from day 7 adults onwards ( Fig. 1B- C). Thus, GLP-1 /Notch receptor activation equalizes across the PZ as animals age.
Although this in vivo method allowed to compare Notch activity within living animals of the same age groups, a zone-by-zone quantitative comparison of Notch activity between timepoints was unfortunately not possible. First, nuclear RFP levels varied somewhat differentially with age in the sensor and control strains (Figs. 1C-D). This complication was heightened by a generalized increase in PZ nuclear RFP/GFP ratios over aging in the sensor strain ( Fig. 1B- C). For instance, the average nuclear RFP/GFP ratio for Z1 was 1.7 at A1, versus 4.1 at A10 ( Fig. 1B ). This would counterintuitively suggest that distal GLP-1 activity increases with age. However, this apparent increase in GLP-1 activity in aged GSCs may more likely be a result of their slower turnover, which may allow the NICD::TEVp to release nuclear GFP from distal germ nuclei over a prolonged duration. Moreover, since the ubiquitin–proteasome system becomes dysregulated with age (Koyuncu et al., 2021), the proteolytic NICD::TEVp degradation may become less efficient in aged GSCs. Finally, the inter-individual variability in RFP/GFP ratios tended to increase with age ( Fig. 1B ), likely reflecting the compounded effects of stochasticity (Meyer and Schumacher, 2024). We believe these results highlight important limitations of the SALSA assay towards aging experiments.
Overall, and combined with the literature, our results show that an age-dependent decline in the transcription of direct Notch target genes in GSCs, together with the proximal drift of the DTC and the reduced turnover rate of GSCs, collectively eliminate the detectable distal-proximal decreasing gradient of Notch signalling activity in vivo .
Methods
** * C. elegans * strains and maintenance **
C. elegans * were maintained at 20°C on standard nematode growth medium (NGM) seeded with E. coli bacteria of the strain OP50 (Brenner, 1974). Late L4 hermaphrodites were selected based on vulva development (Seydoux, 1993) and transferred to new plates in batches of 20 until they were collected for analysis, thus for up to 10 additional days. Worms were transferred to new plates on days 2, 5 and 7 to keep their progeny away.
Imaging
Animals at the indicated ages were paralyzed by soaking in an 8 µL drop of a 0.1% tetramisole (Sigma, L9756) M9 solution on a glass coverslip that was then flipped onto a 3% agarose/M9 pad placed on a microscope slide. Imaging was done within a 30 min window from mounting to minimize any stress signaling (Zellag et al., 2021). The intestine is intertwined with the gonad arms and upon mounting, half of it usually lies on top of one gonad arm. This reduces the detectable fluorescence in that “hidden” arm. Thus, imaging of both gonad arms per animal was rarely done; only when they both lay in the same focal plane and were not obstructed by the intestine. Covered pads were sealed with VALAP (1:1:1 Vaseline, lanolin, paraffin), and 0.35 μm z-step stacks were acquired with a Leica SP8 point scanning confocal microscope and a HC PL APO CS2 40x/1.30 numerical aperture oil objective. GFP and mCherry were excited/collected at 488/495-539nm and 552/590-632nm, respectively, each with 1% laser intensity.
Quantification and modelling
Notch signalling quantification was performed using Imaris 9.2.1 using the spots tool, by modelling 3.6 µm diameter spheres over individual GSC nuclei based on the red ( mCherry::H2B ) channel. The intensity sums for the red and green channels from within the spheres were measured to generate individual RFP/GFP ratios for each GSC nucleus (Martel et al., 2025 ). We considered the cells contained within the first 14 cell diameters from the distal tip, binned into 7 two-cell diameters zones (Z1-Z7). The switchless control RFP/GFP ratios were normalized to 1, to correspond to the expected equimolar nuclear quantities of GFP and RFP; sensor values were transformed according to their respective switchless control average values per zone (Shaffer and Greenwald, 2022).
Statistical Analysis
Statistical analyses were performed using GraphPad Prism 10.2.0. For multi-group comparisons we used the one-way ANOVA with Tukey’s multiple comparisons when the data set passed the Shapiro-Wilk normality test (Figs. 1B: A1, A7; D), and the Kruskal-Wallis with Dunn’s multiple comparisons for sets with non-Gaussian distributions ( Fig. 1B : A4, A10). For Fig. 1D, as A1 data failed the Shapiro-Wilk normality test we used again the Kruskall-Wallis test, this time with Mann-Whitney U tests followed by a Bonferroni correction of the resulting P-values.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Austin Judithe Kimble Judith 1987111 glp-1 Is required in the germ line for regulation of the decision between mitosis and meiosis in C. elegans Cell 5140092-867458959910.1016/0092-8674(87)90128-03677168 · doi ↗ · pubmed ↗
- 2Borbolis Fivos Syntichaki Popi 2015121 Cytoplasmic m RNA turnover and ageing Mechanisms of Ageing and Development 1520047-6374324210.1016/j.mad.2015.09.00626432921 PMC 4710634 · doi ↗ · pubmed ↗
- 3Brenner S 197451 THE GENETICS OF CAENORHABDITIS ELEGANS Genetics 7711943-2631719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 4Crittenden Sarah L. Leonhard Kimberly A. Byrd Dana T. Kimble Judith 200671 Cellular Analyses of the Mitotic Region in the Caenorhabditis elegans Adult Germ Line Molecular Biology of the Cell 1771059-15243051306110.1091/mbc.e 06-03-017016672375 PMC 1552046 · doi ↗ · pubmed ↗
- 5Debès Cédric Papadakis Antonios Grönke Sebastian KaralayÖzlem Tain Luke S. Mizi Athanasia Nakamura Shuhei Hahn Oliver Weigelt Carina Josipovic Natasa Zirkel Anne Brusius Isabell Sofiadis Konstantinos Lamprousi Mantha Lu Yu-Xuan Huang Wenming Esmaillie Reza Kubacki Torsten Späth Martin R. Schermer Bernhard Benzing Thomas Müller Roman-Ulrich Antebi Adam Partridge Linda Papantonis Argyris Beyer Andreas 2023412 Ageing-associated changes in transcriptional elongation influence longevity Nature 61679580028-083681482110.1038/s 415 · doi ↗ · pubmed ↗
- 6Fevr T, Robine S, Louvard D, Huelsken J. 2007. Wnt/β-catenin is essential for intestinal homeostasis and maintenance of intestinal stem cells. Molecular and cellular biology. 27: 7551–7559. 6.10.1128/MCB.01034-07PMC 216907017785439 · doi ↗ · pubmed ↗
- 7Henderson S T Gao D Christensen S Kimble J 199791 Functional domains of LAG-2, a putative signaling ligand for LIN-12 and GLP-1 receptors in Caenorhabditis elegans.Molecular Biology of the Cell 891059-15241751176210.1091/mbc.8.9.17519307971 PMC 305734 · doi ↗ · pubmed ↗
- 8Herndon Laura A. Schmeissner Peter J. Dudaronek Justyna M. Brown Paula A. Listner Kristin M. Sakano Yuko Paupard Marie C. Hall David H. Driscoll Monica 2002101 Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans Nature 41969090028-083680881410.1038/nature 0113512397350 · doi ↗ · pubmed ↗