C. elegans under starvation produce proteinaceous material that supports collective aggregation into web-like structures
May Li, Rex Kerr, Saul Kato

TL;DR
Starving C. elegans worms create a protein-rich material that helps them form web-like structures together.
Contribution
Discovery of a novel proteinaceous material produced by starved C. elegans that facilitates collective aggregation.
Findings
Starved C. elegans secrete a protein-rich, mucoid substance that supports aggregation into web-like structures.
The substance is likely composed of cuticles, dead worm remnants, and yolk.
The material is both secreted and consumed by C. elegans under starvation conditions.
Abstract
We observed the aggregation of starved C. elegans into web-like patterns scaffolded by an unknown chemoattractive extracellular substance that is protein-rich, mucoid, water-insoluble, elastic, and likely to be both secreted and consumed by C. elegans . Under time-lapse imaging, we observed the formation of both the aggregation structures and the proteinaceous substance after populating an NGM plate with 100-200 worms and letting them starve over seven days. We preliminarily characterized the substance using Coomassie, WGA, and DAPI staining. We surmise that the substance may be composed of cuticles, remnants of dead worms, and worm yolk.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
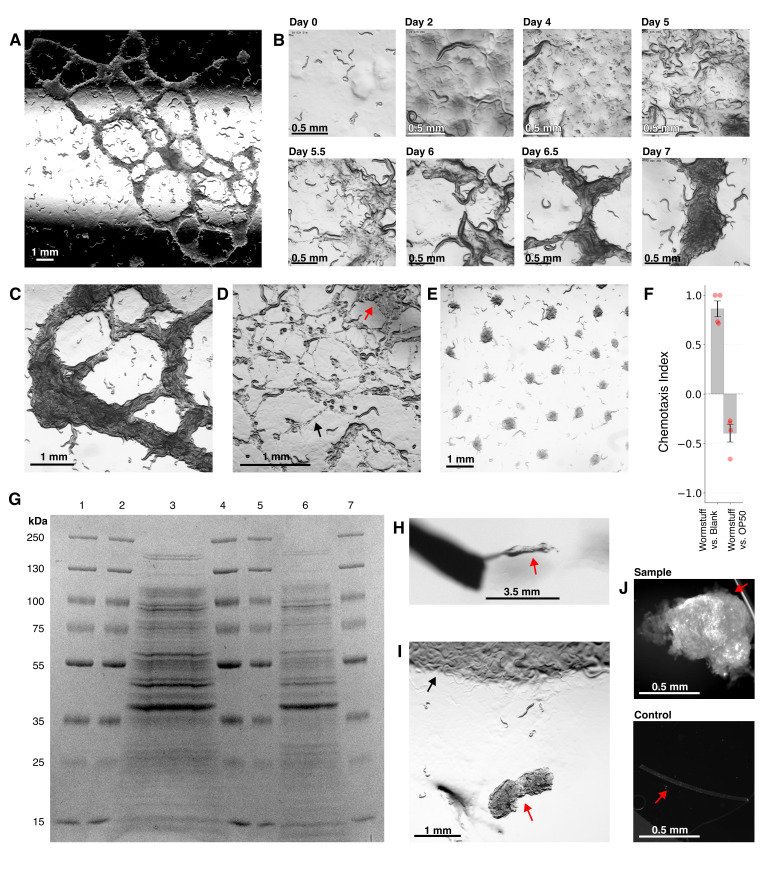 Figure 1
Figure 1|
Strain |
Genotype |
Corresponding Injection Plasmid |
Available from |
|
|
|
N/A |
CGC |
|
FOC126 |
|
pFC078 osm-10p::WOrMsChRmine::wrmScarlet @ 5ng/uL |
focolab.org |
|
FOC134 |
|
pFC082odr-10p::WOrMsChRmine::wrmScarlet@ 2.5ng/uL +
|
focolab.org |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Alzheimer's disease research and treatments · Cellular transport and secretion
Description
We observed the formation of interconnected web-like structures of aggregated mixed-age * C. elegans * approximately 7 days after seeding 100-200 starved worms on OP50 plates ( Fig. 1A ). The structures, which we term worm webs , bear some resemblance to previously reported transient spatial structures of worms. Transient structures have been induced with starved culture conditions combined with factors such as ethanol, humidity, population density, or condensation (Artyukhin et al., 2015, Sugi et al., 2019, Artyukhin et al., 2013, Chen et al., 2021), or result from collective feeding behavior (Demir et al., 2020, Ding et al., 2019). However, unlike other reported structures, worm webs maintained their geometry for days. Closer inspection of the worm webs revealed an extracellular proteinaceous biomaterial, which we term wormstuff , upon which the animals aggregated. We reliably reproduced worm webs using three different strains: N2 , FOC126, and FOC134, for a total of 60 replicates.
Time-lapse video and visual assessment of multiple plates revealed that the formation of worm webs occurs in a stereotypical sequence of visually distinctive phases ( Fig. 1B ; Extended Data). Initially, worms disperse throughout the plate in typical foraging behavior ( Fig. 1B, Day 0). New colonies of bacteria then grow, presumably as a result of residual bacteria deposited from the gut or cuticle from prior exposure to OP50 , providing sufficient food for animals to reach adulthood and lay eggs ( Fig. 1B, Day 2 and Day 4). As progeny begin to exhaust the food, thick mucoid deposits of material that differ in shape and color from existing bacteria begin to appear ( Fig. 1B, Day 5; Fig. 1D, red arrow). Worms aggregate upon these patches of material, which narrow into thin threads or coalesce into broader ropes, resulting in a web-like structure ( Fig. 1B, Day 5.5 and 6; Fig. 1D ). When bacteria are exhausted, the majority of the worm population aggregates upon this collective structure while displaying active pharyngeal pumping and webs often thicken and consolidate ( Fig. 1B, Day 6.5 and 7; Fig. 1C ). The structure then gradually disappears as the population continues to starve, suggesting that the worms can consume the material, albeit slowly relative to typical bacterial consumption rate.
This web formation is markedly different from starvation in typical culture conditions where worms are introduced to a seeded plate, starve, and aggregate into disk-shaped clusters that do not contain external material ( Fig. 1E ; Artyukhin et al, 2015). Worm webs differ from previously reported collective structures because they are stable for multiple days and appear to rely on an external substance (Chen et al., 2021, Sugi et al., 2019, Artyukhin et al, 2015). To assess whether wormstuff was bacterial or fungal in nature, we isolated the material on separate plates where it maintained its shape, neither growing nor shrinking. When not sterilized, bacterial colonies grew around it; heat sterilization (5 minutes in PBS buffer at 98°C) prevented bacterial growth. We then performed spinning disk confocal imaging of the sample with DAPI staining and observed no obvious cellular structures or nuclei. Thus, we tentatively conclude that wormstuff is not predominantly bacteria or fungi, and therefore is either excreted by worms or composed of worm remnants.
We conducted a preliminary investigation of the structural and biochemical properties of wormstuff. When it first appears in pools ( Fig. 1D, red arrow), the material is viscous, gel-like, and difficult to isolate with a pick. The initial thin thread-like strands ( Fig. 1D, black arrow) were also too sparse to harvest easily. However, on around Day 6, elastic masses of wormstuff visible to the naked eye could be lifted up using a worm pick while maintaining its integrity ( Fig. 1H ) and strands could be amassed. We found that wormstuff is not readily water-soluble and remains intact when submerged in distilled H 2 O, which allows removal of worms by washing. However, heat treatment altered its structure: after 5 minutes at 98°C in buffer and reducing agent, the material separated into a soluble supernatant and insoluble dense pellet.
To test wormstuff for protein components, we first confirmed protein existence with a BCA assay, then ran an SDS-PAGE Coomassie gel to assess the distribution of protein sizes present ( Fig. 1G ). We observed a complex banding pattern that was less complex than that of whole-animal gels, but too complex to suggest or confirm individual protein constituents. We also stained, washed, and processed Day 8 wormstuff samples with wheat germ agglutinin (WGA) stain and found that the sample fluoresced brightly, indicating high glycoprotein/glycolipid content ( Fig. 1J, Sample, red arrow) compared to a negative control of a human hair ( Fig. 1J Control, red arrow).
One candidate component of wormstuff is worm yolk. Worm yolk is produced by post-reproductive adults which convert their intestines to yolk, then continuously vent the yolk to provide a nutritional resource for progeny (Kern et al., 2021), peaking at days 4-6 of adulthood. Another candidate component of wormstuff is molted cuticles or cuticular proteins; cuticles consist of glycoproteins, glycolipids, cross-linked collagens, and insoluble proteins (Page et al., 2007) and are shed repeatedly during development. A third candidate is worm corpses. During acute starvation, many * C. elegans * strains are known to bag: larvae hatch inside the mother, consume its internal contents, and leave behind an empty cuticle (Chen et al., 2004). Although we observed bagged worms on worm webs from Day 7 onwards, we observed initial deposits of wormstuff at around Day 4 before the development of worm bagging; thus, wormstuff is not likely to be composed exclusively of worm corpses.
Perhaps the most striking characteristic of the worm webs is that the worms aggregate on and within wormstuff ( Fig. 1A, Days 5+; Fig. 1C ; and Extended Data). When we transferred wormstuff and worms from a starved plate onto a seeded plate, many worms remained in the wormstuff rather than migrating to the OP50 lawn ( Fig. 1I ). In chemotaxis assays, worms strongly preferred wormstuff to an empty target area, but moderately preferred OP50 over wormstuff ( Fig. 1F ), indicating that attraction to wormstuff likely plays a role in the aggregation behavior. This attraction may not be to the proteinaceous material itself even though the animals appear to gradually consume the material; for instance, worm yolk is not chemoattractive even though it is consumed by larvae (Kern et al., 2021). It is also possible that the material becomes saturated with pheromones from worms, bacterial odors, or other small molecules. After treatment in PBS buffer for 5 minutes at 98°C, wormstuff no longer appeared to attract worms, suggesting either a change in composition or that the treatment removed chemoattractive cues.
We observed collective behavior of aggregation upon a chemoattractive proteinaceous substrate that likely originates from worms, creating three-dimensional spatial structures over the course of 7 days. Worm webs are a geometrical intermediate between dispersal and point-like aggregation. We hypothesize that the production of wormstuff-supported worm webs represents an adaptive strategy to extend the spatial coverage and prolong the lifetime of a pre-dauer (i.e. actively feeding) colony.
Methods
Worm Culture Conditions
C. elegans * strains were maintained at 20°C on nematode growth medium (NGM) plates seeded with * Escherichia coli * OP50 according to standard protocols (Stiernagle et al., 2006). To generate worm webs, 100-200 mixed-age unwashed worms were picked from either a prior starved or unstarved plate, introduced to unseeded NGM plates, and left for 7 days in 20°C. On the seventh day, wormstuff was gathered for assays. We found that while picking worms from plates that already contained worm webs would guarantee formation of a new worm web, doing so is not required to induce worm web formation. We presume that the emergence of bacterial colonies observed in the process of worm web formation, as evident in the timelapse video (Extended Data), was the result of residual bacteria carried over on or in the worms.
Chemotaxis Assays
Chemotactic attraction to wormstuff was quantified using a two-choice assay. Mixed-stage worms (100-200 worms per plate), either well-fed N2 or starved FOC126 from web-forming plates, were placed in a central loading zone on unseeded NGM plates. On opposite sides of the plate, two target areas were selected. In one, we placed isolated wormstuff (approximately 0.1 μL) 3 hours before the start of the assay. The other we either left blank or seeded with 1 μL of OP50 24 hours before the start of the assay. After 2 hours at room temperature, worms that migrated to each target area were counted. The chemotaxis index (CI) was calculated as:
CI = (# worms at wormstuff target − # worms at other target) / total # of worms
Four replicates were performed for each condition, two with well-fed N2 and two with starved FOC126; these did not differ dramatically from each other and thus were grouped. CI values range from -1 (complete aversion) to +1 (complete attraction), with 0 indicating no preference. Statistical significance was assessed using one-sample t-tests comparing CI to zero (no preference).
Imaging and Microscopy
Images of worm webs were captured on a Leica M165 FC stereo microscope equipped with a Leica K5 camera. Timelapse recordings were captured on a Leica M205 FCA stereo microscope with an Orca Flash 4.0 C13440 camera. High-resolution fluorescence imaging was performed using a Yokogawa W1-SoRa spinning disk confocal microscope. Contrast adjustments, when needed for clarity, were performed using ImageJ/FIJI version 2.16. Timelapse videos were extracted from Leica .lif files using a custom utility written in Scala 3.7 and utilizing BioFormats 8.3 image readers (available at https://github.com/focolab/lif-timelapse-to-mp4/releases/tag/v0.1.0 ). Movies were encoded using ImageJ/FIJI or the Bytedeco JavaCV FFmpeg API version 1.5.
Protein Extraction and Gel Electrophoresis
The wormstuff material was mechanically isolated from plates using platinum wire picks and washed three times with distilled H 2 O to remove adherent bacteria and worms. The isolated material was transferred to microcentrifuge tubes containing RIPA lysis buffer (50 mM Tris-HCl pH 8.0, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS) supplemented with protease inhibitor cocktail.
Samples were sonicated using a probe sonicator at amplitude 1 for three 5-second bursts with 30-second intervals. Following sonication, lysates were centrifuged at 17,000 × g for 10 minutes at 4°C to separate soluble and insoluble fractions. The supernatant (soluble fraction) was carefully removed and transferred to a new tube.
The remaining pellet (insoluble fraction) was subjected to additional extraction by resuspending in 2% SDS buffer and incubating at 55°C for 10 minutes with periodic vortexing. Following this treatment, samples were centrifuged again at 17,000 × g for 10 minutes at 4°C, and the supernatant representing the SDS-solubilized portion of the insoluble fraction was collected. Some material remained in the pellet. Both soluble and insoluble fractions were quantified using the Pierce BCA Protein Assay Kit.
Samples were prepared for electrophoresis by adding NuPAGE LDS Sample Buffer (4×) and NuPAGE Sample Reducing Agent (10×) to achieve final concentrations of 1× each. Samples were denatured by heating at 98°C for 5 minutes, which both unfolds proteins and reduces disulfide bonds to linearize the polypeptide chains.
Proteins were separated by SDS-PAGE using precast NuPAGE 4-20% Bis-Tris protein gels with 10 wells. Electrophoresis was performed in NuPAGE MES SDS Running Buffer at 200 V for approximately 45 minutes until the dye front reached the bottom of the gel. Protein molecular weight markers (PageRuler Plus, Thermo Fisher Scientific) were loaded in lanes 1, 2, 4, 5, and 7 to enable molecular weight determination.
Following electrophoresis, gels were stained with Coomassie Brilliant Blue R-250 using a standard staining protocol. Briefly, gels were fixed in 40% methanol and 10% acetic acid for 30 minutes, stained in Coomassie Brilliant Blue R-250 solution for 1 hour, and destained in 40% methanol and 10% acetic acid until protein bands were clearly visible against a clear background.
Wheat Germ Agglutinin (WGA) Fluorescence Staining To detect glycoproteins and glycolipids in the wormstuff, samples were stained with fluorescently conjugated wheat germ agglutinin (WGA), which binds to N-acetylglucosamine and sialic acid residues. Isolated wormstuff was fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) for 30 minutes at room temperature.
Fixed samples were permeabilized with 0.2% Triton X-100 in PBS for 20 minutes to allow antibody penetration, followed by one PBS wash. Samples were then incubated with WGA conjugated to PhenoVue Fluor 488 fluorophore (20 μg/mL in PBS) for 30 minutes at room temperature in the dark. Following two final PBS washes to remove unbound WGA, samples were mounted on glass slides using DAPI Fluoromount-G mounting medium with spacers to prevent compression of three-dimensional structures. As a negative control, human hair samples were processed identically to test for non-specific binding of WGA.
Images of stained material were captured on the spinning disk confocal and assessed visually. Staining was bright but amorphous so the images were not analyzed further.
DAPI Staining
To assess whether the proteinaceous material contained living cells or contaminating organisms, samples were stained with 4',6-diamidino-2-phenylindole (DAPI). Isolated wormstuff was fixed with 4% paraformaldehyde in PBS for 30 minutes at room temperature.
Samples were permeabilized with 0.2% Triton X-100 in PBS for 20 minutes to allow DAPI penetration, followed by two PBS washes.
Samples were mounted directly using DAPI Fluoromount-G mounting medium without spacers and imaged on a spinning disk confocal. Staining was dim, even at maximum laser power, and amorphous, so the images were not analyzed further.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Artyukhin Alexander B. Schroeder Frank C. Avery Leon 2013927 Density dependence in Caenorhabditis larval starvation Scientific Reports 312045-232210.1038/srep 02777 PMC 378496024071624 · doi ↗ · pubmed ↗
- 2Artyukhin Alexander B. Yim Joshua J. Cheong Cheong Mi Avery Leon 2015527 Starvation-induced collective behavior in C. elegans Scientific Reports 512045-232210.1038/srep 10647 PMC 444503826013573 · doi ↗ · pubmed ↗
- 3Chen J Caswell-Chen EP 200461 Facultative Vivipary is a Life-History Trait in Caenorhabditis elegans.J Nematol 3620022-300X 10711319262794 PMC 2620758 · pubmed ↗
- 4Chen Yuping Ferrell James E. 2021816 C. elegans colony formation as a condensation phenomenon Nature Communications 1212041-172310.1038/s 41467-021-25244-9PMC 836817834400648 · doi ↗ · pubmed ↗
- 5Demir Esin Yaman Y Ilker Basaran Mustafa Kocabas Askin 202046 Dynamics of pattern formation and emergence of swarming in Caenorhabditis eleganse Life 92050-084X 10.7554/elife.52781 PMC 720289532250243 · doi ↗ · pubmed ↗
- 6Ding Siyu Serena Schumacher Linus J Javer Avelino E Endres Robert G Brown André EX 2019425 Shared behavioral mechanisms underlie C. elegans aggregation and swarminge Life 82050-084X 10.7554/elife.43318 PMC 652222031021320 · doi ↗ · pubmed ↗
- 7Kern Carina C. Townsend St John Salzmann Antoine Rendell Nigel B. Taylor Graham W. Comisel Ruxandra M. Foukas Lazaros C. Bähler Jürg Gems David 2021105 C. elegans feed yolk to their young in a form of primitive lactation Nature Communications 1212041-172310.1038/s 41467-021-25821-y PMC 849270734611154 · doi ↗ · pubmed ↗
- 8Page Anthony 2007 The cuticle Worm Book 1551-850710.1895/wormbook.1.138.1PMC 478159318050497 · doi ↗ · pubmed ↗