Parvalbumin interneurons: the dark and bright sides of a key playmaker of neural circuits and behavior
Eesha Wirk, Charles Quairiaux, Thomas Marissal

TL;DR
This paper reviews parvalbumin interneurons, their role in brain circuits, and how their dysfunction can lead to neurological and psychiatric disorders.
Contribution
The paper provides an updated overview of parvalbumin interneurons and suggests new directions for future research.
Findings
Parvalbumin interneurons are crucial for hippocampal and cortical circuit dynamics and cognitive functions.
Dysfunction of these interneurons is linked to neurological and psychiatric disorders like epilepsy.
Some disruptions caused by parvalbumin interneuron dysfunction may be reversible.
Abstract
With their morphological and electrophysiological properties as well as exceptional connectivity, parvalbumin interneurons play a major role in the dynamics of the neural circuits of the hippocampus and cortex, along with associated cognitive functions. Their dysfunction, which is sometimes reversible, contributes to significant disruptions in network activity and behavioral deficits related to various diseases such as epilepsies or neuropsychiatric disorders. In this Mini Review, we present these parvalbumin interneurons, their characteristics, pathophysiological roles, and propose avenues for future investigations.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
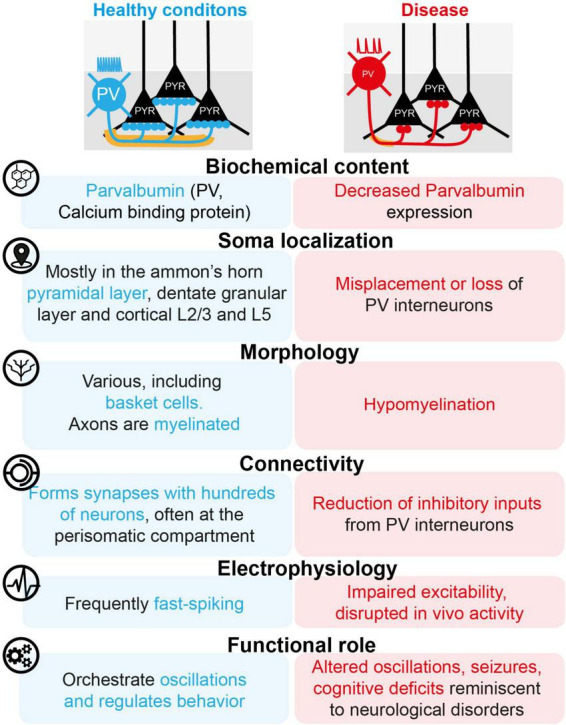 FIGURE 1
FIGURE 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroscience and Neuropharmacology Research · Neurogenesis and neuroplasticity mechanisms · Epilepsy research and treatment
Introduction
1
Parvalbumin interneurons (PVIs) constitute a small fraction of the total neurons of the hippocampus and cortex (2.5%–10%) (Houser, 2007; Bezaire and Soltesz, 2013; Druga et al., 2023). Even within the heterogeneous population of GABAergic inhibitory cells, PVIs are present in numbers comparable to or even lower than other subtypes, such as interneurons expressing VIP or NPY in the hippocampus (Rudy et al., 2011; Pelkey et al., 2017). However, PVIs receive disproportionate attention, which is explained by their many remarkable features in healthy conditions, that enable them to orchestrate network dynamics and control associated behaviors. Consequently, the artificial manipulation (using cell-specific genetic tools) or the alterations (in rodent models related to diseases) of PVI properties can lead to disruption of neuronal activity, the onset of epileptic seizures, and failure of cognitive functions. Here, we describe the exceptional characteristics of PVIs, their contribution to normal function and disease, as well as therapeutic approaches that target them.
Hippocampal and cortical PVIs: from the physiology to disease
2
PVIs under healthy condition
2.1
With their somata particularly concentrated in certain subregions, such as the stratum pyramidale of the Ammon horns, the edges of the granular layer in the dentate gyrus, and layers L2/3 and L5 of the cortex, PVIs display many distinctive properties, detailed below according to the traditional nomenclature used to classify interneurons (Ascoli et al., 2008; Figure 1).
The dark and bright sides of PVIs.
PVIs morphologies
2.1.1
Parvalbumin interneurons in the hippocampus and cortex are divided into several subcategories, based on their axonal arborization. The two best-documented categories are basket cells (or BCs) and chandelier cells (also called “axo-axonic” cells, or AACs).
In Ammon’s Horn of the hippocampus, parvalbumin-expressing BCs have a large pyramid or spindle-shaped soma. Generally lacking spines, their dendrites extend from the alveus to the stratum lacunosum-moleculare. Emerging from the soma or a primary dendrite, their axons form numerous collaterals in the pyramidal layer, giving it a basket-like appearance, with some incursions into the strata oriens and radiatum (Pelkey et al., 2017).
In the dentate gyrus of the hippocampus, BCs have a similar morphology, extending their axons into the granular layer and their dendrites from the outer molecular layer to the hilus (Koh et al., 1995). Unlike Ammon’s Horn, dentate BCs are covered with dendritic spines, the density of which is regulated by experience (Kaufhold et al., 2024).
In the cortex, parvalbumin-positive BCs have a generally multipolar dendrite and an axon that forms a plexus, which can be large and cover several cortical layers and columns or be smaller and restricted to a single layer (Kubota, 2014; Tremblay et al., 2016).
In CA1, AACs have axons that branch out into the pyramidal layer and the superficial part of the stratum oriens (Li et al., 1992). The main branches often extend horizontally, with vertically arranged terminals and rows of synaptic boutons, giving them a distinctive candelabrum appearance (Pelkey et al., 2017). Their dendrites extend either radially from the alveus to the stratum lacunosum-moleculare (Li et al., 1992) or horizontally, spreading exclusively in the stratum oriens, parallel to the pyramidal layer, over several hundred micrometers (Ganter et al., 2004). AACs with similar morphology have been identified in CA3, the dentate gyrus, and the cortex (Somogyi, 1977; Buhl et al., 1994; Viney et al., 2013).
Parvalbumin can be expressed by other morphological subtypes, such as the bistratified cells (BiS) of Ammon’s horns (Halasy et al., 1996; Pelkey et al., 2017), the O-LM cells (Oriens lacunosum-moleculare) of Ammon’s horns and their equivalent in the dentate gyrus, called “HIPP” for “hilar perforant path associated cells” (McBain et al., 1994; Freund and Buzsáki, 1996; Pelkey et al., 2017), the multipolar bursting cells of the cortex (Blatow et al., 2003), as well as long-distance projection neurons of the hippocampus (Jinno et al., 2007; Wick et al., 2017; Yen et al., 2022) and the cortex (Bertero et al., 2020). It should be noted that the axons of PVIs in the hippocampus are often surrounded by a myelin sheath (Stedehouder et al., 2017).
Connectivity
2.1.2
While some PVI subtypes preferentially form synapses on the dendrites of excitatory cells, such as hippocampal O-LM or BiSs (Pelkey et al., 2017), the majority of hippocampal and cortical PVIs target the perisomatic compartment of numerous pyramidal cells and efficiently control their output (Freund and Katona, 2007; Stokes and Isaacson, 2010). AACs exclusively contact the initial segment of the axon of hundreds of pyramids in the hippocampus (Li et al., 1992) and cortex (Wang Y. et al., 2016). Parvalbumin-expressing BCs form synapses with the soma and proximal dendrites of 1500–2500 pyramids in CA1 of the hippocampus, forming half a dozen synapses with each of them (Freund and Buzsáki, 1996; Pelkey et al., 2017). In the cortex, each BC innervates 200–1000 pyramids with 5–15 terminal boutons (Karube et al., 2004; Kubota, 2014). The extensive dendrites of CA1 BCs receive a large number of convergent excitatory inputs, both local (from pyramidal neurons) and distant (e.g., entorhinal cortex) (Gulyás et al., 1999; Tukker et al., 2013). Interestingly, parvalbumin-containing BCs inhibit the deep pyramids of CA1 more strongly but receive more excitation from pyramids located in the superficial part of the pyramidal stratum. Similarly, PVIs tend to innervate pyramidal neurons projecting to the amygdala but receive preferential excitation from pyramids projecting to the prefrontal cortex (Lee et al., 2014). In the cortex and other structures such as the presubiculum (Peng et al., 2021), although observations suggest that PVIs randomly innervate surrounding pyramidal cells and receive excitatory inputs from most nearby pyramids, it appears that the strongest reciprocal connections occur between PVIs and pyramids participating in the same functional process (Packer and Yuste, 2011; Znamenskiy et al., 2024).
Biochemical markers
2.1.3
In PVIs, the expression levels of the calcium-binding parvalbumin protein itself differ depending on the morphological subtype. Thus, parvalbumin labeling is weaker in BiSs and O-LM cells in the hippocampus than in AACs or BCs (Ferraguti et al., 2004). Key to PVI function (Zhang et al., 2025), parvalbumin expression vary depending on experience (Donato et al., 2013) or neuronal activity (Patz et al., 2004; Rupert and Shea, 2022). This form of plasticity appears to be related to perineuronal nets (marked by aggrecan or vicia villosa agglutinin) that mainly envelop PVIs (Yamada et al., 2015).
In addition, PVIs, particularly BCs in the hippocampus and cortex, are characterized by the compartmentalized expression of a combination of proteins associated with rapid, strong, and efficient signaling (Hu et al., 2014). Thus, PVI possesses calcium-permeable AMPA-type glutamatergic receptors that lacks GluA2, but contains GluA1 and GluA4 subunits, as well as GABRA1-containing GABAergic receptors, which are associated with fast-acting excitatory and inhibitory postsynaptic currents (Geiger et al., 1995, 1997; Bartos et al., 2002; Hong et al., 2024). Similarly, the supercritical density of NaV1.1 and NaV1.6 sodium channels along axons, combined with myelination (Micheva et al., 2021), allows for rapid propagation of action potentials (Ogiwara et al., 2007; Lorincz and Nusser, 2008; Hu et al., 2014). At the end of the chain, presynaptic calcium channels of the Cav2.1 or P/Q types, closely coupled to the calcium sensor Synaptotagmin 2, enable rapid and precise secretion of the neurotransmitter GABA (Hefft and Jonas, 2005; Pang et al., 2006; Zaitsev et al., 2007; Bucurenciu et al., 2010; Eggermann et al., 2011; Sommeijer and Levelt, 2012; Rossignol et al., 2013; Lee et al., 2014).
Interestingly, certain protein markers can be used to distinguish between morphological subtypes of PVIs, particularly in the hippocampus. For example, somatostatin has been identified in BiSs and O-LMs (Klausberger et al., 2003, 2004), SATB1 in BCs and BiSs (Viney et al., 2013), NPY in BiSs (Klausberger et al., 2003), and mGluR1α in O-LMs (Ferraguti et al., 2004). AACs of the dentate gyrus are positive for PTHLH and Unc5b (Paul et al., 2017; Proddutur et al., 2023).
Electrophysiological properties and in vivo activity
2.1.4
Parvalbumin BCs of the cortex and hippocampus, as well as BiS and AACs in the hippocampus, are often correlated with a fast-spiking pattern (Hu et al., 2014). This mode is defined as the ability of PVIs recorded ex vivo to generate, following a membrane depolarization plateau, a continuous train of high-frequency action potentials without accommodation (Wang B. et al., 2016). A “stutter firing” pattern, characterized by bursts of action potentials separated by random periods of silence, has been observed in some BiSs (Pawelzik et al., 2002). With distinct electrophysiological characteristics (Tricoire et al., 2011), including strong adaptation of action potential discharge, hippocampal O-LMs rarely exceed an action potential frequency of 50 Hz (Pelkey et al., 2017). The preferential discharge of this type of interneuron occurs in the theta band (Gloveli et al., 2005), in a kainate receptor-dependent manner (Goldin et al., 2007). In the cortex, the multipolar bursting cells are distinguished by an initial burst of action potentials in response to a depolarizing step (Blatow et al., 2003).
Hippocampal PVI subpopulations can be differentiated in vivo according to the spatiotemporal dynamics of their activity in relation to oscillations, which play a role in the computation of behavior (Klausberger and Somogyi, 2008; Fernandez-Ruiz et al., 2023; Huang et al., 2024) and are found to be altered under pathological conditions (Uhlhaas and Singer, 2010). For instance, in awake animals, CA1 AACs fire preferentially during the middle of the descending phases of running-associated theta rhythms (5–10 Hz), while parvalbumin-expressing BCs and BiSs discharge later (Varga et al., 2014). During fast oscillations such as gamma related to running periods (25–90 Hz) and ripples recorded during rest (90–200 Hz), BCs preferentially discharge earliest during oscillatory cycles, followed by BiSs, AACs and O-LM cells (Royer et al., 2012; Varga et al., 2012, 2014; Viney et al., 2013). Double-projecting cells, a fraction of which express parvalbumin, discharge during the trough of theta cycles and just after pyramidal neurons during gamma waves recorded in anesthetized animals (Jinno et al., 2007). In the CA2 and CA3 regions of the hippocampus, PVIs also participate in ripple, theta, and gamma oscillations, but with a different discharge timing than those in CA1 (Tukker et al., 2013; Viney et al., 2013).
Functional properties
2.1.5
The development of genetic tools, based on Cre recombinase (Tsien et al., 1996), which allow selective targeting of biochemical subtypes of inhibitory neurons (Taniguchi et al., 2011) combined with strategies for manipulating neuronal activity using light with optogenetics (Boyden et al., 2005; Deisseroth, 2011) or under the effect of an inert ligand using chemogenetics (Armbruster et al., 2007; Roth, 2016), has enabled the investigation of the functional role of PVI (Raven and Aton, 2021; Tzilivaki et al., 2023).
In the hippocampus, the activity of PVIs controls the synchronization and timing of pyramidal cell firing, as well as the emergence of ripple, theta, or gamma oscillatory activity (Korotkova et al., 2010; Royer et al., 2012; Nguyen et al., 2014; Amilhon et al., 2015; Ognjanovski et al., 2017; Xia et al., 2017; Antonoudiou et al., 2020). Thus, PVIs in the hippocampus contribute substantially to spatial and working memory, memory consolidation (Korotkova et al., 2010; Donato et al., 2013; Xia et al., 2017), representation of novelty (Hainmueller et al., 2024), sensorimotor gating (Nguyen et al., 2014), and control of anxiety behavior (Tiwari et al., 2024; Volitaki et al., 2024). Interestingly, different facets of the same cognitive process are performed by distinct subpopulations of PVIs (Donato et al., 2015; Hainmueller et al., 2024).
Similarly, cortical PVIs promote the synchronization of excitatory cells (Jang et al., 2020), narrow the temporal windows of pyramidal neuron response to sensory afferent (Pedroncini et al., 2024), and orchestrate oscillations (Cardin et al., 2009; Sohal et al., 2009). Consequently, PVIs contributes to a wide variety of cortical functions, such as sensory processing (Yang et al., 2017), memory (Xia et al., 2017), social discrimination (Deng et al., 2019), emotion recognition (Fujima et al., 2025), avoidance behaviors (Ho et al., 2025), and attention (Kim et al., 2016).
In conclusion, PVIs display specific morphophysiological characteristics that enable them to act as essential components of the networks. However, these key interneurons are highly vulnerable to pathological factors (Ruden et al., 2021) and their dysfunction can have harmful effects on the functions of the hippocampus or cortex.
PVIs under pathological conditions
2.2
PVIs under artificial manipulation conditions
2.2.1
It is possible to use genetic tools to manipulate the molecular and electrophysiological properties or connectivity of PVIs in the hippocampus and cortex of mice in order to render them dysfunctional. These disruptions are sufficient to cause activity and network disorders like those observed in neurological disorders such as schizophrenia, autism, and epilepsy.
Thus, chemogenetic or optogenetic inhibition (Nguyen et al., 2014; Hu et al., 2025), depletion of parvalbumin expression (Wöhr et al., 2015), mitochondrial dysfunction (Inan et al., 2016), or deletions of Erbb4 (del Pino et al., 2013), D2-type dopamine receptors (Tomasella et al., 2018), type 5 metabotropic glutamate receptors (Barnes et al., 2015) or NMDA-type glutamate receptors (Korotkova et al., 2010) specifically in PVIs in the hippocampus or cortex lead to disturbances in oscillatory dynamics (e.g., increased or decreased theta and gamma activity) and to cognitive deficits (e.g., memory deficits, impaired locomotion, abnormal emotional and social behavior, impaired sensory-motor gating) that mimic symptoms identified in patients with schizophrenia or autism. Even more spectacularly, permanent silencing of PVIs in the subiculum, a region of the hippocampal formation, is sufficient to induce recurrent spontaneous limbic seizures in mice, a pathological feature reminiscent of temporal lobe epilepsy (Drexel et al., 2017).
Taken together, these data suggest that dysfunction of PVIs was sufficient to cause the development of symptoms associated with neurological diseases. This prompted the scientific community to take the following step: to investigate models that accurately reproduce the symptoms of neurological diseases to determine whether PVIs in the hippocampus and cortex were altered and whether they could represent a valid target for more specific therapeutic strategies.
Parvalbumin interneuron in disease models
2.2.2
Parvalbumin interneurons dysfunction has been identified in the cortex and hippocampus of many models that reliably reproduce the causes of neurological diseases (environmental, genetic, or a combination of both) as well as the symptoms identified in patients.
Thus, the pathological features of neurodevelopmental disorders such as autism and schizophrenia can be mimicked in rodents, for instance by perinatal immune activation (reproducing a microbial infection during development) or by the deletion of the DISC1 gene (linked to schizophrenia) or FMR1 gene (Fragile X syndrome) or the 22q11.2 locus (DiGeorge syndrome). In addition to frequent alterations in rhythmic activity and behavior, these models are often correlated with a disruption of the properties of PVIs (Figure 1), such as a loss of PVIs (Pignataro et al., 2023), a change in the expression and plasticity of parvalbumin itself (Sauer et al., 2015; Mukherjee et al., 2019), a reduction in the expression of ion channels (Qi et al., 2025), a reduction in the number of excitatory inputs received by PVIs (Sauer et al., 2015), a reduction in the number of inhibitory inputs received by pyramids from PVIs (Sauer et al., 2015), as well as misplacement (Meechan et al., 2012), hypomyelination (Maas et al., 2020; He et al., 2025), disruption of ex vivo excitability (Marissal et al., 2018; Hijazi et al., 2023), and reduction of sensory-evoked activity in vivo (Goel et al., 2018). Interestingly, specific chemoactivation of PVIs is sufficient to restore alterations in cortical and hippocampal network activity in vivo, as well as cognitive alterations in mouse models of environmentally or genetically induced neuropsychiatric disorders (Goel et al., 2018; Marissal et al., 2018; Mukherjee et al., 2019; Arime et al., 2023; Pignataro et al., 2023).
In the case of epilepsy, temporal lobe epilepsy (TLE) models are probably the most commonly used. These models are often based on an insult in the form of prolonged seizures (or Status Epilepticus) induced by the administration of kainate or pilocarpine. After a latency this leads to the emergence of epileptic seizures (primarily in the in the hippocampus, which is the main epileptic focus in TLE), and behavioral comorbidities. In these models, many alterations affect the PVIs of the hippocampus. Thus, some of the PVIs degenerate during the latent phases of the disease (Dinocourt et al., 2003), although other publications suggest that they may be relatively spared (Shuman et al., 2020; Matringhen et al., 2025). The survivors undergo changes in their morphological and electrophysiological properties. This is reflected in particular by the sprouting of the axons of commissurally-projecting PVIs (Wick et al., 2017) and by the decrease in their excitability in the dentate gyrus of TLE mouse models (Proddutur et al., 2023). Interestingly, a decrease in the excitability of PVIs in the hippocampus and cortex has also been found in mouse models of genetic forms of epilepsy (e.g., deletion of voltage-gated sodium channel NaV1.1 linked to Dravet syndrome) (Tai et al., 2014; Favero et al., 2018). On this basis, several therapeutic strategies have been tested to compensate for the loss of interneurons or restore the properties of PVIs (Marissal, 2021). For example, the transplantation of stem cells from the medial ganglionic eminence (Upadhya et al., 2019), a substantial proportion of which differentiate into PVIs, reduces seizures and improves the behavior of mice. Similarly, optogenetic stimulation of PVIs, sometimes coupled with a closed-loop system (Krook-Magnuson et al., 2013), can have a beneficial effect on seizures and behavioral deficits (Kim et al., 2020), although activation of PVIs can also have paradoxically pro-epileptic effects (Lévesque et al., 2019).
Discussion
3
Parvalbumin interneurons possess exceptional morphophysiological properties that enable them to contribute significantly to the dynamics of cortical and hippocampal networks, as well as to behavior. Their importance in healthy conditions partly explains why their malfunction is frequently found to be associated with disease.
However, the respective pathophysiological roles of each heterogeneous subtypes of PVIs are poorly understood and should be explored in the future. Recently, tools have become available to selectively target certain subtypes, such as AACs using strategies based on the PTHLH or Unc5b markers (Raudales et al., 2024), and enabled the identification of the changes undergone by AACs after an epileptic insult (Proddutur et al., 2023).
Moreover, it remains to be determined how PVI dysfunction is affected by and affects other elements of the inhibitory microcircuits of the hippocampus and cortex in pathological conditions. Computational and experimental data suggest that PVIs dynamically cooperate under non-pathological conditions with other interneuron subtypes such as calretinin-, VIP-, somatostatin, or CCK-containing interneurons to modulate cortical and hippocampal plasticity, activity and behavior in a manner dependent on the context or behavioral state (Wang et al., 2004; Jang et al., 2020; Udakis et al., 2020; Dudok et al., 2021; Bos et al., 2025; Onorato et al., 2025; Parker et al., 2025). How this “division of labor” between interneurons is disrupted in conditions of disease is an important avenue for investigation with the aim of developing more specific and effective therapeutic strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amilhon B. Huh C. Y. L. Manseau F. Ducharme G. Nichol H. Adamantidis A. (2015). Parvalbumin interneurons of hippocampus tune population activity at theta frequency. Neuron 86 1277–1289. 10.1016/j.neuron.2015.05.027 26050044 · doi ↗ · pubmed ↗
- 2Antonoudiou P. Tan Y. L. Kontou G. Upton A. L. Mann E. O. (2020). Parvalbumin and somatostatin interneurons contribute to the generation of hippocampal gamma oscillations. J. Neurosci. 40 7668–7687. 10.1523/JNEUROSCI.0261-20.2020 32859716 PMC 7531548 · doi ↗ · pubmed ↗
- 3Arime Y. Saitoh Y. Ishikawa M. Kamiyoshihara C. Uchida Y. Fujii K. (2023). Activation of prefrontal parvalbumin interneurons ameliorates working memory deficit even under clinically comparable antipsychotic treatment in a mouse model of schizophrenia. Neuropsychopharmacology 49:720. 10.1038/s 41386-023-01769-z 38049583 PMC 10876596 · doi ↗ · pubmed ↗
- 4Armbruster B. N. Li X. Pausch M. H. Herlitze S. Roth B. L. (2007). Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. U. S. A. 104 5163–5168. 10.1073/pnas.0700293104 17360345 PMC 1829280 · doi ↗ · pubmed ↗
- 5Ascoli G. A. Alonso-Nanclares L. Anderson S. A. Barrionuevo G. Benavides-Piccione R. Burkhalter A. (2008). Petilla terminology: Nomenclature of features of GAB Aergic interneurons of the cerebral cortex. Nat. Rev. Neurosci. 9 557–568. 10.1038/nrn 2402 18568015 PMC 2868386 · doi ↗ · pubmed ↗
- 6Barnes S. Pinto-Duarte A. Kappe A. Zembrzycki A. Metzler A. Mukamel E. (2015). Disruption of m Glu R 5 in parvalbumin-positive interneurons induces core features of neurodevelopmental disorders. Mol. Psychiatry 20 1161–1172. 10.1038/mp.2015.113 26260494 PMC 4583365 · doi ↗ · pubmed ↗
- 7Bartos M. Vida I. Frotscher M. Meyer A. Monyer H. Geiger J. R. P. (2002). Fast synaptic inhibition promotes synchronized gamma oscillations in hippocampal interneuron networks. Proc. Natl. Acad. Sci. U. S. A. 99 13222–13227. 10.1073/pnas.192233099 12235359 PMC 130614 · doi ↗ · pubmed ↗
- 8Bertero A. Zurita H. Normandin M. Apicella A. J. (2020). Auditory long-range parvalbumin cortico-striatal neurons. Front. Neural Circuits 14:45. 10.3389/fncir.2020.00045 32792912 PMC 7390902 · doi ↗ · pubmed ↗