Non-coding RNAs in the viral host-pathogen interaction: molecular regulation and therapeutic potential
Maham Yamin, Nirmin Alsahafi, Rwaa Hussin Abdulal, Muhammad Asad, Mohammad Bosaeed, Ali Zohaib

TL;DR
Non-coding RNAs regulate viral infections by affecting host immunity and viral replication, offering potential for new treatments and diagnostics.
Contribution
This review provides an updated overview of the roles and therapeutic potential of non-coding RNAs in viral host-pathogen interactions.
Findings
Non-coding RNAs modulate host immune responses and viral replication in infections like HCV, DENV, and SARS-CoV.
ncRNAs such as Miravirsen and lncRNA NEAT1 show promise as biomarkers and therapeutic agents for viral diseases.
Challenges remain in translating ncRNA research into effective clinical treatments for viral infections.
Abstract
Non-coding RNAs (ncRNAs), including microRNA (miRNA), long non-coding RNA (lncRNA) and circular RNA (circRNA), serve as key regulatory molecules in the context of viral infection. They play dual roles by modulating host immune responses and influencing viral replication, persistence, and disease progression. Numerous ncRNAs have been implicated in infections caused by viruses such as HCV, DENV and SARS-CoV. This review highlights the biogenesis and multifaceted functions of both host-encoded and virus-encoded ncRNAs in shaping host-pathogen interactions. It also examines their potential as novel biomarkers and therapeutic agents for viral infections. We discuss translational applications such as Miravirsen, a miRNA inhibitor that reached clinical trials for Hepatitis C Virus (HCV) and diagnostic relevance of lncRNA NEAT1 in SARS-CoV-2 infection. In the end, we have also addressed the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
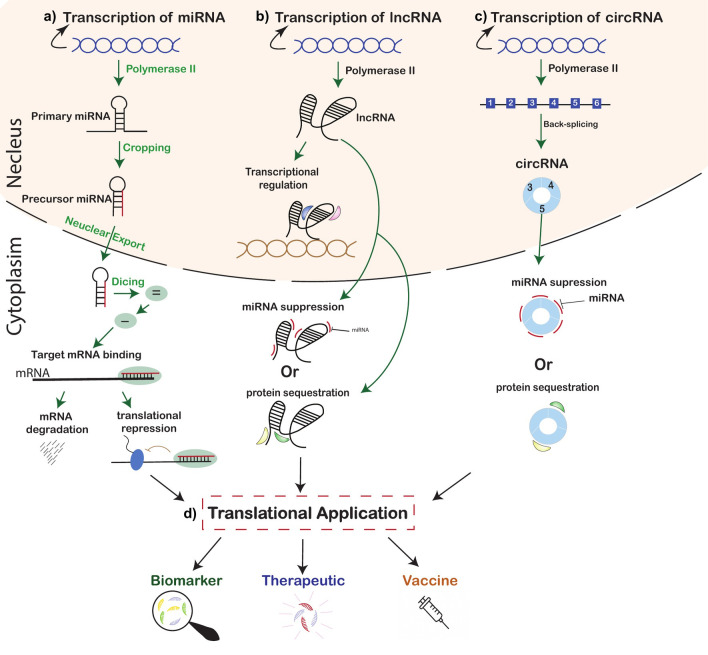 Figure 1
Figure 1| miRNA | Role in viral replication | Associated virus | Reference |
|---|---|---|---|
| miR-122 | Pro-viral effect for hepatitis C virus (HCV) replication, | HCV, HBV | ( |
| miR-1 | Promotes HBV replication | HBV | ( |
| miR-501-3p | Promotes HCV replication | HCV | ( |
| miR-27a | Inhibits HCV replication | HCV | ( |
| miR-28 | Contributes towards human immunodeficiency virus | HIV | ( |
| miR29a | Inhibit HIV-1 replication | HIV-1 | ( |
| miR-127-3p | Inhibit influenza A virus (IAV) replication | IAV | ( |
| miRNA-133a | Inhibits dengue virus (DENV) replication | DENV | ( |
| miR-36 | Inhibits Kaposi’s sarcoma-associated herpesvirus (KSHV) replication | KSHV | ( |
| miR-15a | Inhibits SARS-CoV-2 replication | SARS-CoV-2 | ( |
| lncRNA | Role in viral replication | Associated virus | Reference |
|---|---|---|---|
| lncRNA Dreh | Repress HBV‐related HCC growth | HBV | ( |
| LncRNA | Low expression is associated with poor prognosis in HBV‐related HCC. | HBV | ( |
| lncRNA NRON | Suppress viral transcription | HIV | ( |
| lncRNA#32 | Regulates encephalomyocarditis virus (EMCV), HBV and HCV replication | EMCV, HBV, HCV | ( |
| lncRNA-IFI6 | Regulates antiviral innate immunity | HCV | ( |
| lncRNA MALAT1 | A promoter of HIV-1 transcription | HIV | ( |
| lncRNA NRAV | Modulates antiviral immune responses | IAV | ( |
| lncRNA TSPOAP1 | Promotes IAV replication | IAV | ( |
| lncRNA NRAV | Promotes RSV replication | RSV | ( |
| lncRNA BC200 | Promotes Epstein-Barr virus (EBV) associated nasopharyngeal carcinoma | EBV | ( |
| lncRNA FANCI-2 | Play role in modulating oncogenic pathways in human papillomavirus (HPV) related cancers | HPV | ( |
| lncRNA | Promotes herpes simplex virus (HSV) proliferation | HSV | ( |
| lncRNA PIRAT | Host immune regulation during SARS-CoV-2 infection | SARS-CoV-2 | ( |
| lncRNA VILMIR | Host immune regulation during SARS-CoV-2 and RSV infections | SARS-CoV-2, RSV | ( |
| lncRNA CHROMR | Regulates host antiviral immune responses | SARS-CoV-2 | ( |
| lncRNA ANRIL | Potential circulatory biomarkers | SARS-CoV-2 | ( |
| circRNA | Viral infection | Mode of action | Reference |
|---|---|---|---|
| circ_0000479 | Hantaan Virus | Indirectly regulated RIG-I expression by sponging miR-149-5p | ( |
| circ_MerTK | IAV | Negatively regulates innate immune response | ( |
| circ_VAMP3 | IAV | Restricts IAV replication | ( |
| circ_Slco3a1 | IAV | May be involved in IAV induced lung injury | ( |
| circ_0000479 | SARS-CoV-2 | Regulates the immune response against SARS-CoV-2 | ( |
| circRNA_10156 | HBV | Pro-tumorigenic | ( |
| cFAM210A | HBV | Inhibits HCC tumorigenesis | ( |
| circ_0000976, circ_0007750 and circ_013989 | HBV-related HCC | Diagnostic panel for HCC | ( |
| circ_0001400 | KSHV | Aids in maintaining latent infection | ( |
| circ_ARFGEF | KSHV | Regulates KSHV mediated oncogenesis | ( |
| circ_0007321 | ZIKV | Regulates Zika virus replication | ( |
| circ_FNDC3B | MERS-COV | Regulates virus replication | ( |
| circRNA_101764 | HBV-related HCC | Regulation of HCC associated tumorigenesis | ( |
| hsa_circ_0000220 | Japanese Encephalitis Virus (JEV) | Regulates immune responses and inflammation | ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer-related molecular mechanisms research · Circular RNAs in diseases · MicroRNA in disease regulation
Introduction
1
Emergence and reemergence of viral infections, such as SARS-CoV, MERS-CoV, Zika virus (ZIKV), Ebola virus, SARS-CoV-2, Crimean-Congo hemorrhagic fever virus (CCHFV) and influenza A virus (IAV), have persistently threatened global health (Schwartz, 2021). Moreover, factors such as climate change, global trade and an increase in international travel can contribute to the rapid spread of infectious diseases (Hoffman and Maldonado, 2024; Almulhim et al., 2025). In today’s globalized world, a small outbreak in one region can result in a global pandemic. In addition to these factors, these viral agents can rapidly evolve and evade the host’s immune surveillance system. The emergence of viral mutations at the genomic level is a persistent challenge and it continuously threatens public health. These mutations contribute towards virus escape, while on the other hand, they compromise vaccine effectiveness as well. In addition to this, these mutations can also result in false negative results, hindering effective surveillance efforts. This necessitates the development of novel diagnostic and therapeutic strategies (Han et al., 2023; Yu et al., 2024).
During viral infection, host immune system is activated and results in complex interactions. Although protein-based host-virus interactions have been extensively studied, little is known about the role of non-coding RNAs (ncRNAs) during viral infection. Recent studies have reported the integral role of ncRNAs, such as microRNA(miRNA), long non-coding RNAs (lncRNAs) and circular RNAs (circRNAs) during viral infection (Wang et al., 2017a; Liu et al., 2019a; Withers et al., 2019; Henzinger et al., 2020; Maarouf et al., 2023). One of the extensively studied ncRNAs in host-virus interactions is miRNA. miRNAs, either host-derived or virus-derived, have been reported to play a key role in virus infection and host immune responses. These small non-coding (sncRNA) RNAs target specific mRNAs involved in the host immune response mechanism and viral replication (Bannazadeh Baghi et al., 2024; Bahojb Mahdavi et al., 2025). Similarly, another class of ncRNAs, lncRNAs, has been extensively investigated during viral infections. Increasing evidence suggests that they regulate transcription, translation and protein interactions during viral infection (Liu et al., 2019a; Statello et al., 2021; Mattick et al., 2023). Recently, circRNAs have also been emerging as key regulators of immune responses during viral infection. circRNAs interact with miRNAs and modulate their interactions with target mRNAs, resulting in altered gene expression (Wang et al., 2017a; Belousova et al., 2018; Maarouf et al., 2023).
Given their essential roles in viral pathogenesis, ncRNAs are now increasingly explored for their potential as viral therapeutic or diagnostic tools. Several studies have reported that ncRNAs can inhibit viral infection. For example, miR-let-7c is a host-derived miRNA and has been reported to suppress the hepatitis C virus (HCV) replication (Chen et al., 2019). Similarly, lncRNA lncITPRIP-1 results in increased production of interferon, resulting in inhibition of HCV replication (Xie et al., 2018a). Like miRNAs and lncRNAs, circRNAs have been reported to inhibit viral replication via both direct and/or indirect mechanisms. For example, circRNA circVAMP3 not only directly inhibits IAV replication, but this specific circRNA also restores host immune responses against IAV (Min et al., 2023). Another miRNA, miR-122, is being studied as a promising therapeutic agent against HCV infection. Two antisense locked nucleic acid (LNA) inhibitors (i.e., Miravirsen and RG-101) have been developed. These LNA inhibitors directly target miR-122, which results in the effective suppression of HCV infection in chronic patients. However, following cessation of LNA inhibitor therapy, HCV rebound has been observed (Panigrahi et al., 2022).
Several studies have also reported the altered expression of ncRNAs during viral infection. Researchers are now focusing on the development of novel diagnostic and prognostic biomarkers using these altered expression profiles of ncRNAs during viral infections. Recently, Liu and colleagues have reported that COVID-19 patients display unique ncRNA expression patterns, correlating with disease severity (Liu et al., 2023). Another study on dengue virus (DENV) infection has reported that lncRNA NEAT1 displays altered expression patterns in dengue patients (Pandey et al., 2017). Taken together, these findings indicate that ncRNA expression patterns can be used as potential diagnostic and prognostic tools during viral infection.
Classification and biogenesis of ncRNAs
2
ncRNAs do not encode proteins, yet play a crucial role in gene expression. It is estimated that each cell has around 10 million RNA molecules, with ncRNAs constituting a significant portion of this total. Keeping in view their diverse functions, ncRNAs are divided into two groups: housekeeping ncRNAs and regulatory ncRNAs. Housekeeping RNAs include ribosomal RNA (rRNA), transcriptional RNA (tRNA) and small nuclear RNA (snRNA). These ncRNAs are generally involved in regulating cellular functions. Regulatory ncRNAs, on the other hand, are further subdivided based on their size. Those regulatory ncRNAs, which are longer than 200 nucleotides, are referred to as lncRNAs, whereas those shorter than 200 nucleotides are called smaller nuclear RNAs (snRNAs). The snRNAs further include miRNAs, small interfering RNA (siRNA) and piwi-interacting RNAs (piRNAs). circRNAs are another type of regulatory ncRNAs and are single stranded, covalently closed RNA molecules with sizes ranging from 100 nucleotides to 10,000 nucleotides (Belousova et al., 2018; Zhang et al., 2019; Caba et al., 2021; Cardon et al., 2021; Oo et al., 2022; Ghani et al., 2025).
Non-coding RNAs are produced through various biogenetic pathways involving various cellular machinery (Figure 1). miRNA biogenesis is a multi-step process that starts in the nucleus and concludes in cytoplasm. miRNAs are initially transcribed by RNA polymerase II and sometimes by polymerase III as long primary miRNAs (pri-miRNAs) transcripts. These pri-miRNAs possess characteristic stem-loop structures. Following transcription in the nucleus, pri-miRNAs are then processed by a microprocessor complex consisting of RNase III enzyme Drosha and its cofactor DGCR8. This processing results in precursor miRNAs (pre-miRNAs). Pre-miRNAs are typically around 70 nucleotides in length. These pre-miRNAs are then transported to the cytoplasm. Once in the cytoplasm, the pre-miRNAs are further processed by Dicer enzyme, which cleaves the pre-miRNAs to generate mature double-stranded miRNAs. One strand of this miRNA duplex is referred to as guide strand, is incorporated into the RNA-induced silencing complex (RISC). The second strand or passenger strand is usually broken down. The guide strand then directs RISC to target mRNAs to regulate gene expression (Balasubramaniam et al., 2018; O’Brien et al., 2018; Bannazadeh Baghi et al., 2024; Brillante et al., 2024; Hynes and Kakumani, 2024; Bahojb Mahdavi et al., 2025).
Biogenesis, mechanisms, and translational relevance of non-coding RNAs (ncRNAs). Schematic representation illustrating the major classes of non-coding RNAs and their biological and translational significance. (a) Biogenesis of microRNAs (miRNAs), transcribed by RNA polymerase II and sequentially processed through cropping and dicing to yield mature miRNAs that regulate gene expression via translational repression or mRNA degradation. (b) Biogenesis and functional roles of long non-coding RNAs (lncRNAs), transcribed by RNA polymerase II and acting at transcriptional and post-transcriptional levels as scaffolds, decoys, or modulators of miRNAs and proteins. (c) Formation and activity of circular RNAs (circRNAs), generated through back-splicing and functioning as molecular sponges or regulators of signaling pathways. (d) Translational applications: ncRNAs serve as promising diagnostic and prognostic biomarkers, therapeutic targets, and innovative vaccine platforms, bridging fundamental gene regulation with clinical translation.
The lncRNAs are a diverse group of ncRNAs with varying biogenesis pathways. lncRNAs often share similarities with mRNA biogenesis but with some notable distinctions. The majority of lncRNAs are generated by RNA polymerase II, whereas some can be transcribed by polymerase I too. Although lncRNAs can undergo 5’ capping, splicing and 3’ polyadenylation like mRNAs, however; lncRNAs are often less efficiently spliced or polyadenylated. lncRNAs can originate from different genomic locations, including introns and exons of protein-coding genes. Interestingly, some lncRNAs can serve as a precursor for miRNAs (Marchese et al., 2017; Suarez et al., 2020; Statello et al., 2021; Nojima and Proudfoot, 2022; Mattick et al., 2023).
circRNAs are endogenous ncRNAs that lack 5’ caps and 3’ poly(A) ends and have a covalently closed loop structure, which is formed through a unique splicing event. circRNAs are primarily produced by an alternative splicing mechanism called back-splicing or head-to-tail splicing (Belousova et al., 2018; Caba et al., 2021; Hwang and Kim, 2024). This occurs when a downstream 5’ splice donor site combines with an upstream 3’ splice acceptor site (Kristensen et al., 2019). This non-canonical splicing is enabled by complementary inverted repeat sequences, such as Alu elements or RNA-binding proteins (RBPs), which bring splice sites nearby. Depending on the sequences they comprise, there are various varieties of circRNAs (Ivanov et al., 2015). Exonic circRNAs are composed exclusively of exons. Circular intronic RNAs are produced from intronic lariats that do not undergo debranching. Intronic and exonic sequences are maintained by exon-intron circRNAs. Some factors that rigorously regulate the synthesis of circRNAs by either promoting or stopping circularization include MBL/MBNL1, QKI and FUS. circRNAs are generally more stable than linear RNAs due to their exonuclease resistance. Most circular circRNAs are in the cytoplasm; however, specific nuclear circRNAs have also been identified (Maass et al., 2017).
The role of virus-produced ncRNAs in viral infection and pathogenesis
3
Virus-produced ncRNAs, such as miRNAs, lncRNAs and circRNAs, play essential roles during viral infection. By modulating the host immune responses, these ncRNAs promote viral infection and pathogenesis (Tycowski et al., 2015). For example, Epstein-Barr virus (EBV) produces a variety of viral miRNAs, including miR-BARTs. miR-BART-7 and miR-BART-9 can modulate virus life cycle and cell proliferation (Caetano et al., 2024). Several studies also report that miR-BARTs can target host immune-related genes to prevent apoptosis and to avoid immune surveillance. These miRNAs can also alter viral gene expression and contribute towards virus latency (Iizasa et al., 2020; Ungerleider et al., 2021). Similarly, the Kaposi’s sarcoma-associated herpesvirus (KSHV) produces miRNAs that increase the virus’s ability to proliferate within the host by inhibiting the host’s antiviral responses (Zhao et al., 2023). In case of human immunodeficiency virus (HIV), viral miRNAs can modulate cell signaling pathways to promote immune evasion and chronic infection (Balasubramaniam et al., 2018). Similarly, lncRNA PAN, which is also produced by KSHV, regulates the viral gene expression (Rossetto and Pari, 2014). Several other viral lncRNAs have been reported to modify host cellular pathways to enhance viral persistence in the host (Liu et al., 2019a).
Recently, several studies have reported that circRNAs produced by certain viruses play essential roles in viral pathogenicity. Research indicates that specific viral circRNAs may function as sponges for host miRNAs, affecting the host’s immune response and cellular milieu (Panda, 2018). The circRNAs can sequester miRNAs that typically inhibit viral replication or stimulate antiviral pathways, augmenting the virus’s capacity to replicate within host cells (Wang et al., 2024).
miRNAs as regulatory molecules in viral infections
4
Introduction to miRNAs
4.1
miRNAs are a class of small, non-coding RNA molecules typically 18–25 nucleotides in length that play essential roles in the post-transcriptional regulation of gene expression. miRNAs function by binding to complementary sequences in the 3′ untranslated regions (3′ UTRs) of target messenger RNAs (mRNAs). This results in the degradation or translational repression of the target mRNAs (O’Brien et al., 2018; Bahojb Mahdavi et al., 2025).
miRNAs in viral replication and host signaling regulation
4.2
miRNAs not only regulate cellular signaling but, also play a critical role during viral replication (Bannazadeh Baghi et al., 2024; Bahojb Mahdavi et al., 2025). They affect viral replication by targeting either viral genome and/or by modulating the host immune responses (Table 1). Viral miRNAs aid in virus replication by targeting genes involved in host immune response mechanisms (Liao et al., 2021b; Bannazadeh Baghi et al., 2024; Bahojb Mahdavi et al., 2025). Conversely, the host-derived miRNAs can directly suppress viral RNAs or modulate host response mechanisms. For example, miR-22 has been reported to promote HCV replication by binding to 5UTR of HCV (Kunden et al., 2020). Similarly, miR-210 and miR-199a-3p have been reported to inhibit Hepatitis B Virus (HBV) (Zhang et al., 2010). During Japanese encephalitis virus (JEV) infection, expression of RNF11 is regulated by miR-19b-3p, which results in modulation of antiviral immune responses (Zhu et al., 2015). Recently, miR-98-5p has been identified as a potential suppressor of SARS-CoV-2 replication (Rizkita and Astuti, 2021). On the other hand, miR-146a modulates TRAF6 and promotes IAV replication (Yan et al., 2025). Similarly, miR-373 facilitates viral replication during HSV-1 infection (Dass et al., 2023). miRNA-mediated regulation of neuroinflammation has also been reported during HSV-1 infection (Xie et al., 2018b; Patrycy et al., 2022). During ZIKV infection, miRNAs also modulate host immune responses and fetal development (Carvalho-Silva et al., 2022; Lai et al., 2025). DENV is another arbovirus like ZIKV, which interacts with host miRNAs to regulate host-pathogen interactions. For instance, miR-4488 contributes to cellular immune responses during DENV infection and may serve as a potential biomarker (Filip et al., 2025).
miRNAs can modulate the host immune responses in diversified ways, from expression regulation of pro-inflammatory cytokines to influencing the activity of pattern recognition receptors (PPRs). For example, during viral infection, miR-155 is upregulated, resulting in enhanced interferon production and stronger TH1 responses (Arroyo et al., 2020). Interestingly, miR-146 plays a dual role during viral infection. It can behave as pro-viral or anti-viral, depending upon the context (Nahand et al., 2020). Recently, Chen and colleagues have reported that miR-155 disrupts the mitochondrial homeostasis and regulates interferon responses (Chen et al., 2023). Another miRNA, miR-29, regulates IFN-γ production and Th1 cell development by suppressing T-bet and Eomes (Steiner et al., 2011). Moreover, miR-150 and miR-223 have been reported to attenuate STAT1 signaling in T cells (Moles et al., 2015). Another study highlights that miR-126-5p negatively regulates host antiviral immune responses (Wang et al., 2022).
Translational applications of miRNAs
4.3
RNA-based therapeutics, for example, miRNA mimics and inhibitors, may offer unique opportunities to combat viral infections. They target either host immune responses or virus-specific molecular pathways. One such example is Miravirsen, which is the inaugural miRNA anti-HCV therapeutic. It functions by inhibiting miR-122, which is a proviral miRNA (Brillante et al., 2024). Research is being conducted to explore the potential of miRNA-based antiviral therapies. For instance, miR-122 mimics are being studied for their potential to suppress HBV replication (Mahmoudian-Sani et al., 2019). Additionally, circulating miR-122 has also emerged as a potential biomarker for liver infection, especially in case of HCV infection (Ullah et al., 2022). Similarly, miR-199A-3p is being investigated as a potential therapeutic for HBV. Additionally, anti-miR-210 is being researched to stop cancer-causing effects in liver diseases related to HBV (Zhang et al., 2010).
Studies have shown that viruses can alter host miRNA expression profiles and understanding these miRNA-virus interactions may offer a basis for miRNA-based diagnostic and therapeutic tools. For instance, altered miRNA expression profiles have been observed during SARS-CoV-2 infection. Some host-derived miRNAs were found to be specifically depleted during SARS-CoV-2 infection (Bartoszewski et al., 2020). On the contrary, certain miRNAs are upregulated and contribute towards inflammation and lung injury during viral infection. Targeting these host-virus-miRNAs interactions can result in novel therapeutics.
Circulating miRNAs are stable in serum and can also serve as diagnostic biomarkers. During DENV infection, miR-150 expression levels have been reported to correlate with disease severity (Chen et al., 2014). Wong and colleagues have recently reported that miR-21-5p, miR-146a-5p, miR-590-5p, miR-188-5p and miR-152-3p can be used to detect DENV infection (Wong et al., 2020). Furthermore, miRNA profile in correlation with COVID-19 disease severity has also been reported (Pius-Sadowska et al., 2025). Collectively, these findings indicate the promising potential of miRNA-based therapeutics and diagnostics as new approaches against viral infections.
lncRNAs as regulatory molecules in viral infections
5
Introduction to lncRNAs
5.1
lncRNAs are RNA molecules longer than two hundred nucleotides. They do not code for proteins; however, they play important roles in gene regulation at transcriptional and post-transcriptional levels. Increasing evidence suggests that lncRNAs play important roles during viral infection (Liu et al., 2019a; Statello et al., 2021). Certain lncRNAs enhance viral replication or alter the host cell environment to facilitate viral pathogenesis, whereas others may alter interferon signaling to block infection and support viral clearance. Their wide range of functions indicates that lncRNAs can serve as important candidates for diagnostic and therapeutic applications (Liu et al., 2019a; Aldweik et al., 2025).
lncRNAs in viral replication and host signaling regulation
5.2
lncRNAs have recently been identified as key regulators of the innate immune system and cellular defenses against viral infections. Rather than encoding proteins, these molecules function as modulators that influence key immune signaling pathways, including NF-κB, IRF3 and JAK/STAT. By interacting with transcription factors, chromatin modifiers and RNA-binding proteins, lncRNAs determine whether viral infections are effectively recognized and eliminated or allowed to persist in the host (Table 2). For example, interferon-inducible lncRNAs such as USP30-AS1, NRAV, TSPOAP1-AS1, lnc-MxA and lnc-Lsm3b play pivotal roles in fine-tuning interferon signaling and host antiviral immune responses during IAV infection (Wang and Cen, 2020; Wang et al., 2022; Cao et al., 2025). Similarly, IVRPIE enhances host defense against the influenza A virus by upregulating IFN-β1 and key ISGs through histone modification (Zhao et al., 2020). Another lncRNA#32 promotes type I IFN signaling and ISG expression, thereby restricting EMCV, HBV and HCV replication (Nishitsuji et al., 2016). lncRNA IFITM4P stabilizes IFITM1/2/3 expression by sponging miR-24-3p, which results in increased innate antiviral responses (Xiao et al., 2021).
Numerous lncRNAs have been documented to enhance viral propagation by inhibiting host immune responses. For instance, lncRNA-BTX enhances viral replication by modulating the localization of RNA-binding proteins DHX9 and ILF3 (Cao et al., 2023). Recently, Yan and colleagues have reported that lncRNA-DRNR disrupts the STAT1 activation. This disruption results in the reduced IFN-β mediated antiviral responses (Yan et al., 2024). Another lncRNA, THRIL, has been reported to inhibit IRF3 activation. This inhibition results in a reduction in type I and III interferons and enhanced IAV replication (Chen et al., 2025).
Moreover, several studies have also reported the integral roles of lncRNAs in various host signaling pathways. For example, lncRNA-ACOD1 interacts with GOT2 enzyme, which facilitates viral replication (Wang et al., 2017b). Another lncRNA, HEAL, interacts with FUS protein during HIV infection and facilitates HIV transcription (Chao et al., 2019). A recent study suggests that EBV-BART lncRNAs regulate the host gene expression (Verhoeven et al., 2019).
Translational applications of lncRNAs
5.3
Many lncRNAs have been reported to display expression patterns that correlate with the stage and often severity of viral infection. In recent studies, specific lncRNA expression patterns were observed during influenza and SARS-CoV-2 infections, which can aid in early diagnosis of these viral infections (Van Solingen et al., 2022; Aldweik et al., 2025). High-throughput sequencing has also revealed several differentially expressed lncRNAs during viral infections. In a study, a group of seven lncRNAs has been developed to effectively differentiate between severe and non-severe cases of COVID-19 (Cheng et al., 2021). Another study has also reported that lncRNAs such as HOTAIRM1, PVT1 and AL392172.1 are important regulators of SARS-CoV-2 infection (Moazzam-Jazi et al., 2021). Furthermore, studies conducted with EV71 and DENV have also reported that lncRNAs are associated with immune responses (Wang et al., 2017c; Li et al., 2018). Similarly, during ZIKV infection, altered expression of lncRNAs in neuroprogenitor cells has also been observed (Venkatesan et al., 2022). Recently, Arslan and colleagues have reported that lncRNAs such as ERCP FER1L4 and LOC1100133669 can be used as potential biomarkers to determine the disease severity during CCHFV-mediated infection (Arslan et al., 2022). Taken together, these findings indicate that lncRNAs can be used as novel tools not only to identify viral infection, but also to assess the severity of the disease. However, developing a standardized method to detect and quantify lncRNAs is an open venue, requiring further research.
Similar to diagnostic, therapeutic potential of lncRNAs is also being studied. Several studies have reported both pro- and anti-viral lncRNAs. For example, lncRNA-ACOD1 has been reported to be upregulated during several viral infections, indicating its potential pro-viral activity (Wang et al., 2017b). Moreover, research is being carried out to target virus encoded lncRNAs to determine their use in antiviral therapies. For instance, EBV sisRNAs or KSHV PAN RNA have been targeted to assess the impacts on viral latency and immune clearance (Min et al., 2022; Media et al., 2025).
Moreover, altering certain host-derived lncRNAs expression profiles can aid in the reversal of virus latency and possible approach for eradicating chronic infections (Kulkarni et al., 2023). Current research is focused on optimizing delivery mechanisms, such as lipid nanoparticles and plant-derived exosomes, for the administration of synthetic lncRNAs or CRISPR modulators (Kazemian et al., 2022; Hillman, 2023).
circRNAs as regulatory molecules in viral infections
6
Introduction to circRNAs
6.1
circRNAs are noncoding RNA without a 5′-cap structure or 3′ poly(A) tail. Their 3′ and 5′ ends are covalently bonded, forming a closed circular shape. circRNAs are generated from viral genome transcription/splicing or from host cell gene splicing. These ncRNAs play an important role in regulating host immune responses and viral replication (Belousova et al., 2018; Panda, 2018; Maarouf et al., 2023; Wang et al., 2024).
circRNAs in viral replication and host signaling regulation
6.2
Recent evidence shows that circRNAs can regulate viral replication by acting as competitive endogenous RNAs (ceRNAs) (Table 3). For example, in Hantaan virus infection, circ_0000479 was shown to sponge miR-149-5p, thereby lifting miR-149-5p suppression of RIG-I, which likely enhances antiviral defense (Lu et al., 2020; Yin et al., 2024). In another study with MERS-CoV, knockdown of host circRNAs (e.g., circFNDC3B, circCNOT1) led to significantly reduced viral load, consistent with a ceRNA-based regulatory mechanism (Zhang et al., 2020). Host circRNAs can act as miRNA sponges during viral infection, binding intracellular miRNAs to block their activity and modulate host gene expression, ultimately creating conditions favorable for viral replication. circRNAs–miRNAs interactions are vital in HBV-related hepatocellular carcinoma (HCC). Rui Liao and colleagues studied circRNA_101764, circRNA_100338, circ-ARL3 and circ-ATP5H in HBV-HCC, showing their roles in tumor development and metastasis (Liao et al., 2021a). circRNAs appear to play important roles in virus-induced immune responses. Li and colleagues have recently performed genome-wide sequencing of brain tissues from Japanese encephalitis virus (JEV)-infected mice. This study has revealed the altered expression of circRNA hsa_circ_0000220 (Li et al., 2020b). Which acts like a sponge for miR-326-3p, leading to an increase in cytokine production. Recent studies also indicate that disruptions in ceRNA networks, which involve lncRNAs, circRNAs and ncRNAs, play significant roles in the pathogenesis of SARS-CoV-2 (Aghajani Mir, 2024).
circRNAs can also alter essential cellular pathways by modulating protein stability, phosphorylation and ubiquitination. For instance, host-derived circ_0001400 binds to splicing factor PNISR and downregulates the KSHV gene expression. This results in aversion of apoptosis and development of latency (Tagawa et al., 2023). During IAV infection, the host-derived circVAMP3 disrupts the NP-polymerase interactions as circVAMP3 acts as a decoy for viral nucleoprotein (NP) and non-structural protein 1 (NS1). This interference restores interferon-β production and suppresses viral replication (Min et al., 2023). Together, these findings indicate that circRNA–protein interactions influence both viral pathogenesis and host immune responses. During EV-A71 infection, hsa_circ_0045431 binds hsa_miR-584, forming the circRNAs/NLRP3 axis that activates pyroptosis (Hu et al., 2023). Similarly, hsa_circ_0007321, derived from the DIS3L2 gene, has been reported to play an important role during ZIKV infection.
This circRNA, which is derived from the host, was shown to regulate the viral replication pathway by sponging miR-492. This, in turn, affects the protein NFKBID, which is a negative regulator of the NF-κB pathway (Kang et al., 2023).
In another interesting study, purified circRNAs were introduced into HeLa cells to observe their effect. They seemed to trigger a strong immune response, which was also demonstrated by increased expression of antiviral genes such as RIG-I, PKR, MDA5 and OAS1. As expected, this made the cells more resistant to viral infection and lower infection rates were observed in subsequent assays (Chen et al., 2017; Liu and Chen, 2022).
Translational applications of circRNAs
6.3
Due to their covalently closed structure, circRNAs are highly stable in serum samples, rendering them ideal candidates for the novel diagnostic tools. Moreover, as they are key regulators of immune responses against viral infections, they are being investigated for potential targets for therapeutic interventions. It has recently been reported that CRISPR techniques can be utilized to knock out circRNAs and their role inside different cellular pathways can be further studied (Santer et al., 2019; Yin et al., 2024). A recent study has also reported that circRNAs can be used to diagnose HBV infection (Jiang et al., 2020). Similarly, another circRNA, circ_3205, can aid in the COVID-19 diagnosis. Moreover, circ_3205 is a virus-encoded ncRNA, which also acts as a has-miR-298 sponge and positively regulates SARS-CoV-2 infection (Barbagallo et al., 2022).
Not only are circRNAs highly stable, but they also trigger strong immune responses, making them ideal candidates for RNA-based vaccine platforms. Recently, a study has investigated a potential circRNA vaccine encoding receptor binding domain (RBD) of SARS-CoV-2. This vaccine elicited robust T-cell responses, which may provide broad protection against different variants (e.g., Delta and Omicron) of SARS-CoV-2 (Qu et al., 2022). Amaya and colleagues have further explored the potential of circRNA vaccine in murine tumor models and their study revealed effective CD8+ T cell responses by dendritic cells (Amaya et al., 2023). Initially, scientists were struggling with the circularization techniques; however, with emerging research in synthetic biology, novel circularization techniques are being developed to synthesize circular RNA (Abe et al., 2018). Progress in molecular virology and host immunology, along with synthetic biology, has greatly improved our understanding of ncRNAs-mediated host-pathogen interactions, paving the way for improved diagnostics and therapeutics.
Challenges in non-coding RNA translational research for viral diagnosis, biomarkers and therapeutics
7
Despite their promising potential, the translational aspect of ncRNAs faces several challenges, which are also active areas of research. Due to significant variability among sample collection, type of samples, processing strategies and storage conditions, the ncRNA profiles obtained are highly variable. Low abundance of ncRNAs and sequence similarity are also major challenges, in addition to quantification techniques, which have all been originally developed for longer RNAs. All these factors make detecting small fold changes in ncRNAs during infections a major challenge.
Additionally, ncRNAs exhibit diverse forms and functions and only a small number have been identified and functionally characterized (Kulkarni et al., 2023). Some ncRNAs are present in normal pathophysiological states as well, such as inflammation and tissue injury and are not specific to a particular infection, which makes it difficult to rely on them as specific biomarkers of infection. For example, some lncRNAs (such as LINCO2574, GAPLINC) are involved in regulating innate antiviral immunity but, also show overlapping activation in unrelated infections (Rai et al., 2022; Chen et al., 2024).
One of the major challenges in using ncRNAs as therapeutic agents is their efficient and tissue specific delivery. Even though significant progress has been made with delivery systems as lipid nanoparticles and exosomes, poor penetration and limited organ specificity are still considered major hurdles in ncRNAs application as therapeutic agents. The ncRNAs-based therapeutics also risk off target gene modulation due to not only sequence complementarity, but also competition with endogenous RNAs. Due to all these limitations, very few ncRNAs have reached advanced clinical stages as therapeutic agents, despite promising laboratory results.
Future directions
8
ncRNAs have emerged as central regulators of viral pathogenesis and have also been shown to play an important role in shaping the immune response to viral infections. As fine-tuned regulators, they modulate pattern recognition receptors (PRRs) as well as downstream cytokine and chemokine networks to either activate or suppress antiviral pathways (Tycowski et al., 2015; Zhang et al., 2019; Suarez et al., 2020). These mechanisms regulate the balance between pro- and anti-inflammatory responses, which determine the severity of viral infections. Several ncRNAs directly target viral RNA and interfere with its translation or stability, while others influence the differentiation and activation of macrophages and dendritic cells. The noncoding RNAs can also modulate host epigenetic landscapes, resulting in altered gene expression profiles that may favor either viral persistence or clearance (Tycowski et al., 2015; Wang, 2018; Zhang et al., 2019; Suarez et al., 2020).
Early clinical experience with using ncRNAs as therapeutic and diagnostic agents has been encouraging to an extent. For example, in the case of Miravirsen, which is an LNA based anti-miR against liver enriched miR-122, a dose dependent reduction was observed in the HCV RNA in patients, which showed its clinical effectiveness (Janssen et al., 2013). Additionally, lncRNA NEAT1 has been consistently reported as being upregulated in SARS-CoV-2 positive samples, which could serve as a potential biomarker (Rodrigues et al., 2021). However, a study using miR-34 mimic, a potential cancer therapeutic, had to be halted in phase-1 clinical trial due to immune related issues (Hong et al., 2020).
Understanding specific functions of ncRNAs requires advanced tools that can accurately analyze gene expression, protein expression and cell specific behaviors. For this, new technologies such as single cell sequencing and transcriptomic techniques are being increasingly used to understand how ncRNAs work in different cell types and during different infection stages.
To report the potential of ncRNAs, we must balance enthusiasm with realism. Many ncRNA types remain largely exploratory, while a few, such as miRNAs, have shown promising results. Our review has largely focused on ncRNAs that have substantial experimental validation. The overall success of using ncRNAs as diagnostic and therapeutic agents requires advances in delivery chemistry, omics tools and careful clinical validation. With careful and coordinated efforts, perhaps the next decade will see the ncRNA based tools move from research laboratories to real-world clinical applications.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abe N. Kodama A. Abe H. (2018). “ Preparation of Circular RNA In Vitro,” in Circular RN As: Methods and Protocols.. Eds. Dieterich C. Papantonis A. ( Springer, New York, NY), 181–192. doi: 10.1007/978-1-4939-7562-4_15, PMID: · doi ↗
- 2Aghajani Mir M. (2024). Illuminating the pathogenic role of SARS-Co V-2: Insights into competing endogenous RN As (ce RN As) regulatory networks. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 122, 105613. doi: 10.1016/j.meegid.2024.105613, PMID: 38844190 · doi ↗ · pubmed ↗
- 3Ahluwalia J. K. Khan S. Z. Soni K. Rawat P. Gupta A. Hariharan M. . (2008). Human cellular micro RNA hsa-mi R-29a interferes with viral nef protein expression and HIV-1 replication. Retrovirology. 5, 117. doi: 10.1186/1742-4690-5-117, PMID: 19102781 PMC 2635386 · doi ↗ · pubmed ↗
- 4Ahmed N. Francis M. E. Ahmed N. Kelvin A. A. Pezacki J. P. (2023). micro RNA-185 inhibits SARS-co V-2 infection through the modulation of the host’s lipid microenvironment. Viruses. 15, 1921. doi: 10.3390/v 15091921, PMID: 37766327 PMC 10536008 · doi ↗ · pubmed ↗
- 5Aldweik M. H. Hisham Y. Aldweik M. H. Hisham Y. (2025). Long non-coding RN As in viral immunity: from regulatory mechanisms to therapeutic potential. J. Inflamm. Infect. Med. 1, 3. doi: 10.53941/jiim.2025.100015 · doi ↗
- 6Almulhim M. Ghasemian A. Memariani M. Karami F. Yassen A. S. A. Alexiou A. . (2025). Drug repositioning as a promising approach for the eradication of emerging and re-emerging viral agents. Mol. Divers. 29, 5465–5485. doi: 10.1007/s 11030-025-11131-8, PMID: 40100484 PMC 12638364 · doi ↗ · pubmed ↗
- 7Amaya L. Grigoryan L. Li Z. Lee A. Wender P. A. Pulendran B. . (2023). Circular RNA vaccine induces potent T cell responses. Proc. Natl. Acad. Sci. 120, e 2302191120. doi: 10.1073/pnas.2302191120, PMID: 37155869 PMC 10193964 · doi ↗ · pubmed ↗
- 8Arroyo M. Salka K. Chorvinsky E. Xuchen X. Abutaleb K. Perez G. F. . (2020). Airway mir-155 responses are associated with TH 1 cytokine polarization in young children with viral respiratory infections. Plo S One. 15, e 0233352. doi: 10.1371/journal.pone.0233352, PMID: 32442188 PMC 7244143 · doi ↗ · pubmed ↗