Identification of Circulating MiR-4651 as Novel Biomarker for Metabolic Dysfunction–Associated Steatotic Liver Disease
Mélanie Kirchmeyer, Anthoula Gaigneaux, Florence A. Servais, Anita Arslanow, Claudia Rubie, Markus Casper, Matthias Glanemann, María L. Martínez-Chantar, Marcin Krawczyk, Frank Lammert, Iris Behrmann

TL;DR
The study identifies miR-4651 as a potential blood-based biomarker for detecting liver disease related to metabolic dysfunction.
Contribution
MiR-4651 is newly identified as a reliable biomarker for early detection of metabolic dysfunction–associated steatotic liver disease.
Findings
MiR-4651 and miR-21-5p levels were significantly lower in MASLD patients compared to healthy controls.
MiR-4651 effectively distinguishes simple steatosis from more severe liver disease stages like MASH or cirrhosis.
MiR-335-5p showed strong potential for differentiating hepatocellular carcinoma from healthy individuals.
Abstract
Metabolic dysfunction–associated steatotic liver disease (MASLD) affects >30% of adults and is becoming one of the leading causes of hepatocellular carcinoma (HCC). Reliable biomarkers are needed for the early diagnosis of HCC and detection of chronic liver diseases, like MASLD. Here we assessed the biomarker potential of circulating microRNAs in a cohort of patients genotyped for the risk allele of patatin-like phospholipase domain-containing protein 3 (PNPLA3), associated with increased susceptibility to chronic liver diseases. The cohort comprised 70 MASLD patients (40 with simple steatosis, 9 with metabolic dysfunction–associated steatohepatitis (MASH), 21 with cirrhosis), 47 HCC patients (32 with cirrhosis), and 14 healthy controls. Serum levels of miR-122-5p, miR-146a-5p, miR-146b-5p, miR-21-5p, miR-335-5p, miR-433-3p, miR-4530, miR-4651, miR-526a2, and miR-873-5p were quantified…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1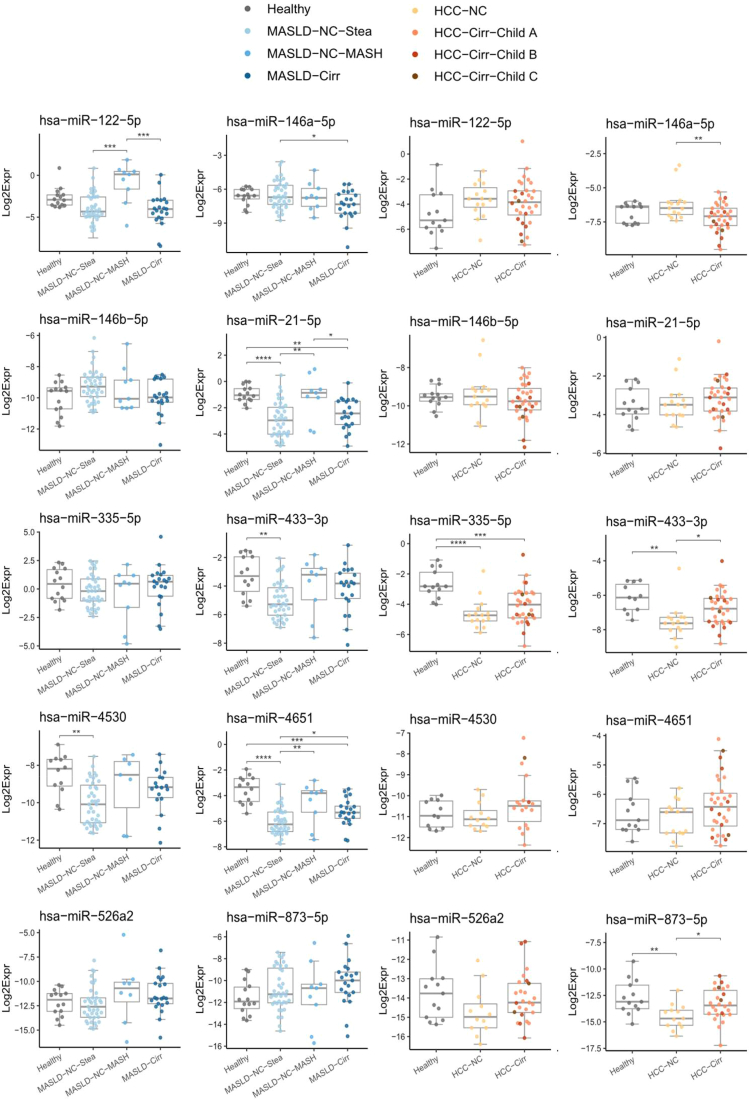 Figure 2
Figure 2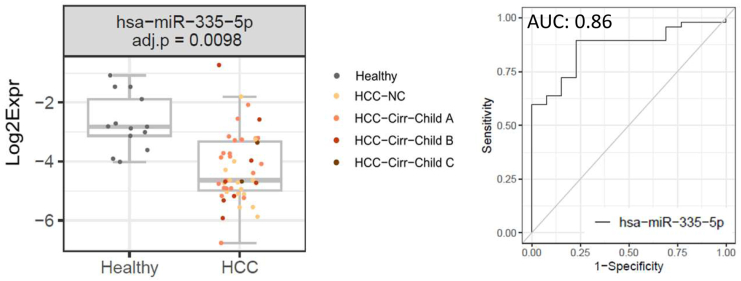 Figure 3
Figure 3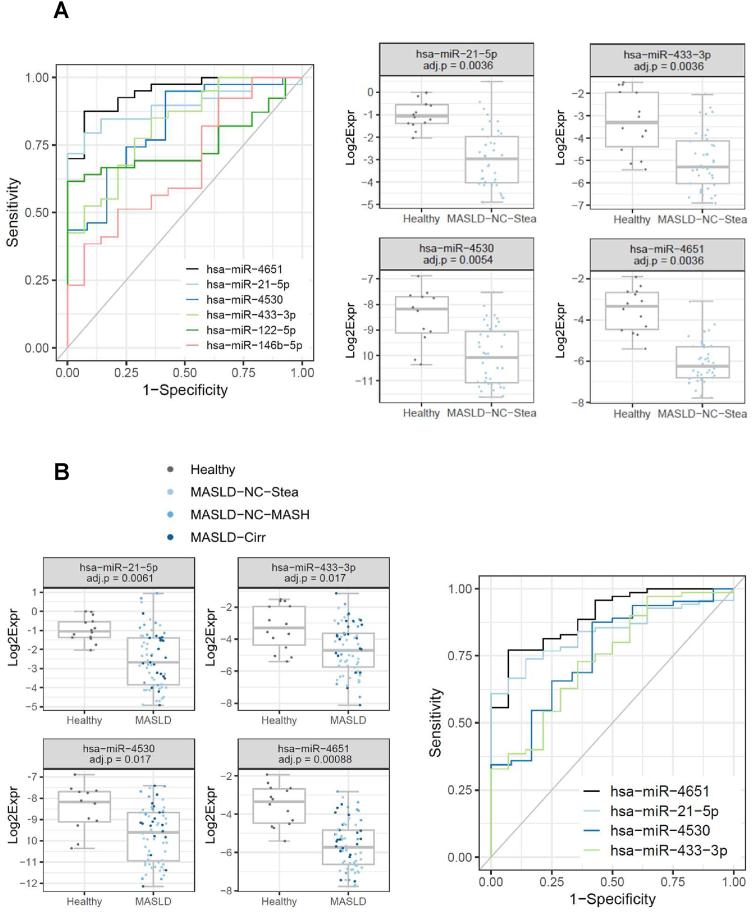 Figure 4
Figure 4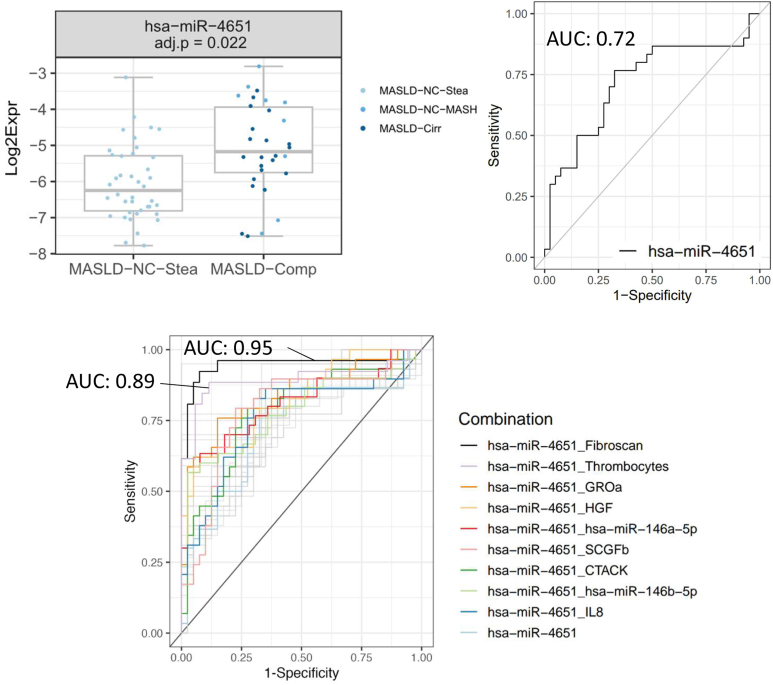 Figure 5
Figure 5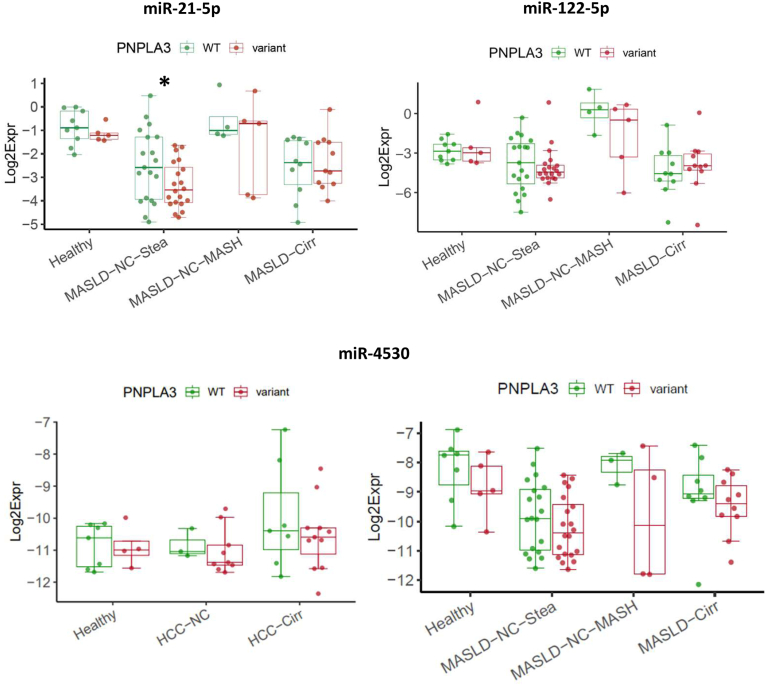 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLiver Disease Diagnosis and Treatment · MicroRNA in disease regulation · Liver Disease and Transplantation
Introduction
Metabolic dysfunction–associated steatotic liver disease (MASLD), the hepatic form of the metabolic syndrome, is a global challenge affecting approximately a third of the world population. While the earliest stage of MASLD, simple steatosis, has usually a benign course, in some patients it progresses to an inflammatory disease, namely metabolic dysfunction–associated steatohepatitis (MASH). MASH can lead to fibrosis and cirrhosis and increases the risk of developing hepatocellular carcinoma (HCC)1 which is the most common form of primary liver cancer. Recently MASLD has been recognized as the most rapidly growing cause of HCC.2 Next to lifestyle and environmental factors, several genes, such as patatin-like phospholipase domain-containing protein 3 (PNPLA3), affect the risk of chronic liver diseases and HCC. As the early stages of HCC are usually asymptomatic, it is often detected at an advanced stage.3 In this context, biomarkers from liquid biopsies hold promise for detecting HCC at earlier stages.
MicroRNAs (miRNAs), short noncoding RNA molecules (20–24 nucleotides), bind to specific mRNA targets, thereby influencing gene expression by inhibition of translation and/or degradation of mRNA. Apart from their intracellular roles, miRNAs can be found in all body fluids. Circulating miRNAs offer several advantages as biomarkers: they are stable, show considerable tissue-specificity, it is possible to quantitatively assess them, and their profiles change during diseases. Thus, there is considerable interest in miRNAs as possible diagnostic biomarkers for various diseases, including HCC and MASLD, either alone or in combination with other disease-associated parameters.4, 5, 6, 7 For example, levels of miR-34-5p together with YKL-40 (chitinase-3-like protein 1) are monitored in the NIS2+^TM^ panel to monitor MASLD progression.4
In this study, we analyzed the serum levels of a 10-miRNA panel in 70 MASLD patients, 47 HCC patients and 14 healthy controls. All individuals were genotyped for the PNPLA3 c.444 C>G rs738409 polymorphism. The risk allele encodes the I148M variant of the PNPLA3 lipase which facilitates hepatic steatosis and increases the risk for liver diseases, including MASLD and HCC. The variant allele has effects on the transcriptome of liver cells.8^,^9 Of note, few studies also reported about distinct miRNA profiles in individuals with chronic liver diseases carrying the PNPLA3 p.I148M risk allele.10, 11, 12 Here we demonstrate that miR-335-5p performs very well as a predictor of HCC in receiver operating characteristic (ROC) curve analyses. Moreover, we describe miR-4651 as a novel predictor for MASLD and for MASLD complication. We did not observe an impact of the PNPLA3 genotype on the serum levels of the miRNAs.
Patients and Methods
Patient Samples
Serum samples from healthy individuals and from patients with chronic liver disorders (MASLD or HCC) were collected at the Saarland University Hospital in Homburg (Germany) at the Institute for Occupational and Environmental Medicine and Public Health or the Department of Medicine II, respectively. Liver-specific diseases were excluded in the healthy controls, and they did not present with elevated liver function tests. The presence of chronic liver diseases other than MASLD—particularly alternative causes of hepatic steatosis such as alcohol-associated liver disease (threshold of alcohol consumption 20 g/day in women and 30 g/day in men) or drug-induced liver injury—was excluded in all patients; none of the patients showed signs of autoimmune, cholestatic, or viral liver diseases. HCC was diagnosed either histologically or noninvasively using contrast-enhanced imaging according to the Liver Imaging Reporting and Data System computer tomography/magnetic resonance imaging v2018 criteria.13
After collection, the whole blood samples were kept at room temperature for 30 minutes for clotting. Then, the tubes were centrifuged (2000 g, 10 minutes, 4 °C). Serum samples were kept at −80 °C until miRNA extraction. Genomic DNA was isolated from ethylenediaminetetraacetic acid-anticoagulated blood samples using the membrane-based QIAamp DNA extraction protocol (Qiagen). The rs738409 polymorphism in PNPLA3 was assessed by polymerase chain reaction (PCR) (TaqMan, assay ID C_7241_10). For the results shown in this manuscript, we compared the homozygote wild-type (WT) genotype [CC] with the “variant” genotypes ([CG] + [GG]). For 101 of the 131 sera, we had previously quantified the levels of 22 cytokines, chemokines, growth factors, or soluble receptors (commonly referred to as “cytokines”).14 Liver stiffness measurements by transient elastography (Fibroscan) are available for most MASLD patients and the healthy individuals. Tables 1 and A1 provide further information on the cohort.Table 1. Characteristics of Patients Included in the Cohort (N = 131)ParametersHealthy (N = 14)MASLD-NC-Stea (simple steatosis) (N = 40)“MASLD complication” (N = 30)HCC-NC (no cirrhosis)(N = 15)HCC-Cirra (cirrhosis)(N = 32)Overall (N = 131)MASLD-NC-MASH (no cirrhosis, MASH)(N = 9)MASLD-Cirr (cirrhosis)(N = 21)PNPLA3 p.I148M genotype CC9 (64.3%)20 (50.0%)4 (44.4%)10 (47.6%)3 (20.0%)10 (31.3%)56 (42.7%) CG3 (21.4%)17 (42.5%)2 (22.2%)10 (47.6%)8 (53.3%)12 (37.5%)52 (39.7%) GG2 (14.3%)3 (7.5%)3 (33.3%)1 (4.8%)4 (26.7%)10 (31.3%)23 (17.6%)Gender F8 (57.1%)16 (40.0%)5 (55.6%)12 (57.1%)2 (13.3%)10 (31.3%)53 (40.5%) M6 (42.9%)24 (60.0%)4 (44.4%)9 (42.9%)13 (86.7%)22 (68.8%)78 (59.5%)Age Mean (SD)49.6 (9.52)55.9 (12.9)47.1 (10.5)59.7 (15.5)70.9 (6.57)66.6 (10.8)59.6 (13.7) Median [min, max]49.5 [25.0,67.0]55.5 [23.0, 80.0]48.0 [31.0, 59.0]62.0 [31.0, 84.0]72.0 [58.0, 80.0]64.0 [45.0, 92.0]61.0 [23.0, 92.0]BMI Mean (SD)30.2 (4.25)28.5 (4.81)29.1 (7.66)25.3 (6.11)29.0 (4.48)29.7 (6.50)28.8 (5.66) Median [min, max]29.5 [25.3, 38.3]28.6 [17.6, 41.1]26.0 [21.8, 41.0]24.8 [17.3, 37.3]29.3 [21.2, 38.1]28.7 [19.9, 46.2]28.3 [17.3, 46.2] Missing0 (0%)15 (37.5%)4 (44.4%)7 (33.3%)0 (0%)0 (0%)26 (19.8%)Fibroscan (kPa) Mean (SD)5.27 (1.53)5.34 (1.58)7.06 (2.00)37.3 (26.1)//12.3 (17.6) Median [min, max]4.55 [3.60, 8.70]5.00 [2.20, 9.10]6.30 [4.00, 10.2]26.3 [10.1, 75.0]//5.60 [2.20, 75.0] Missing0 (0%)0 (0%)0 (0%)4 (19.0%)4 (4.8%b)Demographics, PNPLA3 p.I148M genotype, BMI, and “Fibroscan” (liver stiffness assessed by transient elastography) are indicated for the 6 groups: Healthy, MASLD-NC-Stea, MASLD-NC-MASH, MASLD-Cirr, HCC-NC, HCC-Cirr. Age, BMI, and Fibroscan are presented as mean ± SD, and median. Moreover, the numbers of missing values are indicated.BMI, body mass index; SD, standard deviation.aOf the 32 patients, 22 were classified as Child A, 8 as Child B, and 2 as Child C.bFor Fibroscan, no data were collected for HCC patients. Thus, the percentage was calculated only for the healthy and MASLD samples (N = 84).
MiRNA extraction from serum samples, quality control
We based the protocol largely on our previous study.15 For the HCC samples, we extracted miRNA as duplicates from 200 μL of serum each. We mixed the 2 duplicates, if both fulfilled the quality criteria (see below). For the MASLD samples, due to smaller volumes of serum available, we extracted miRNA as unicates from 50 μL. For the healthy samples, we extracted miRNA twice (in parallel to the patient samples, to ensure the same technical conditions), once as duplicates (200 μL) and once as unicates (50 μL); we analyzed the 2 batches separately in downstream analyses.
Before the addition of Qiazol, the serum was thawed at room temperature and centrifuged (30 minutes, 16,000 g, 4 °C) to remove cellular debris. The miRNA extraction was performed using the miRNeasy serum/plasma kit from Qiagen. We incorporated 3 exogenous C. elegans miRNAs as spike-in controls, cel-miR-39-3p, cel-miR-54-3p, and cel-miR-238-3p, at different concentrations to account for potential biases in the quantification of miRNAs with different levels of abundance. Furthermore, cel-miR-39-3p was used for standardization of data across all samples (see below).
Real-time quantitative polymerase chain reaction quality control was performed prior to array analysis using primers for cel-miR-39-3p, cel-miR-54-3p and cel-miR-238-3p, to control for variations in recovery and amplification efficiency between RNA preparations. Additionally, we assessed levels of miR-451a (highly expressed in erythrocytes) and of miR-23a-5p (high levels in serum and not in blood cells) as an indication for possible hemolysis and of SNORD95 to control for potential white blood cell contamination. 4 μL (for 200 μL extractions) or 6 μL (for 50 μL extractions) out of 14 μL eluted total RNA were reverse-transcribed in a 10 μL reaction volume with the miScript RT II kit (Qiagen) following the supplied protocol using Hispec buffer, which selectively amplifies only mature miRNAs. Real-time PCR detection of the above-mentioned mature miRNAs and of SNORD95 was carried out on a CFX96 Detection System (Bio-Rad) using 1 μL of 1:10 diluted cDNA, 2x iQ SYBR Green Supermix (BioRad), and 10× miRNA-specific primer assay (Qiagen). Specificity of the qPCR primers was assessed by a post-qPCR melting curve analysis. Samples not reaching sufficient quality metrics due to hemolysis, white blood cell contamination, or incomplete recovery of spiked-in controls were discarded.
MiRNA Profiling by qPCR Arrays, Quality Control and Data Analysis
Ten miRNAs were selected for analysis with customized qPCR arrays (Qiagen): miR-122-5p, miR-146a-5p, miR-146b-5p, miR-21-5p, miR-335-5p, miR-433-3p, miR-4530, miR-4651, miR-526a2, and miR-873-5p. Before proceeding to the qPCR arrays, due to generally low amounts of miRNAs in serum samples, we implemented a preamplification step with 1:5 diluted cDNA using the miScript PreAMP PCR kit (Qiagen) along with the appropriate primer mixes (custom primer mix for custom qPCR arrays). Preamplification control experiments were performed by real-time quantitative polymerase chain reaction with primer assays targeting SNORD95, cel-miR-39-3p, and miRTC (internal miRNA reverse transcription control). miR-335-5p, miR-21-5p, and miR-122-5p were detected at highest levels (mean Cq values of 15.4, 16.5, and 17.5), miR-433-3p, miR-4651, and miR-146a-5p at intermediate levels (mean Cq values: 19.2, 19.6, and 20.6), while miR-146b-5p, miR-4530, miR-873-5p, and miR-526a were detected with the lowest levels (mean Cq values: 23.4, 23.8, 25.8, and 26.7). Of note, the relative values for miR-21-5p, miR-122-5p, and miR-146b-5p which we obtained upon preamplification match well to the ones obtained by López-Riera et al., without amplification (Cq values of 28, 29, and 37, respectively5). MiRNAs were profiled using custom qPCR arrays (Qiagen) and a CFX384 Detection System (Bio-Rad). Each qPCR array plate included default internal controls (miRTC, positive PCR control, and cel-miR-39-3p).
For qPCR array analysis, baselines and thresholds were adjusted according to the supplier's recommendations and Cq values were extracted for further analysis. Cq values > 40 or samples exhibiting bad melting curves were considered as “not detected”. To assess the quality of the melting curves, we used a Support Vector Machine–based method generated with the R ‘e1071’ package.15 Cq values obtained with the cel-miR-39-3p primers were used to calibrate the data sets: The mean Cq value of the two technical duplicates for cel-miR-39-3p was subtracted from each sample’s Cq value. From this calibrated data, we calculated the sample expression using the formula: “2^ˆ^-cq_calibrated”.
qPCR Analysis of MiRNAs Isolated from Primary Human Liver Cells upon Stimulation with Hyper-Interleukin-6
See supporting information, p. 2.
Statistical Analysis
Differences between groups in levels of each of the 10 miRNAs were tested, separately for the “50 μL” and “200 μL” batches, using pairwise t-tests from the rstatix package,16 followed by family-wise Holm adjustment. If only 2 groups were tested, a Benjamini-Hochberg (BH) correction17 was applied between variables. Associations between categorical factors were assessed using a chi-square test. Values for all miRNAs, clinical parameters, and cytokines were log-transformed (for exceptions see14). Correlations between miRNAs and cytokines or numeric clinical parameters (with at least 10 pairs of values, see14 for further information) were analyzed with Pearson correlation and P values were corrected using the BH algorithm. The basis of the predictions was a generalized linear model model using a binomial distribution. This model predicts the binary class of a dependent variable (y), given one or several independent variable(s). It was run for each of the 10 miRNAs; P values were corrected using the BH algorithm. When this generalized linear model showed an adjusted P value <.05, we selected it for further ROC curve analysis. The cut-off represents the threshold in y value (class prediction) and was chosen as giving the highest Youden's index. For models with BH-adjusted P values <0.05, we also built models with more variables (second miRNA, a cytokine, clinical parameter, or genotype), with the maximum of 1 predictor per 10 events (“rule-of-thumb limit”). When several variables were included in a model, we excluded correlated pairs of variables (ie, P < .05, R > 0.4 or R < −0.4) to avoid collinearity. All data analysis was conducted using R version 4.2.218 and rstatix package.16 Graphs were generated using ggplot219 and corrplot.20 ROC curves were obtained using ROCit package.21
Results
Study Cohort and PNPLA3 p.I148M Genotype Distribution
We quantified the levels of miRNAs in sera of patients with MASLD or HCC: 70 individuals with MASLD (21 with cirrhosis, “MASLD-Cirr”; 9 with MASH but without cirrhosis, “MASLD-NC-MASH”; 40 with simple steatosis, “MASLD-NC-Stea”), 47 individuals with HCC (32 with cirrhosis, “HCC-Cirr”; 15 without, “HCC-NC”). 14 healthy individuals without chronic liver diseases were in the control group. Demographics and some characteristics of the cases and controls are presented in Table 1; more information on clinical parameters is available in Table A1. All individuals were genotyped for their allele status of PNPLA3. In total, 42.7% of the participants were homozygous for the WT allele [CC], 39.7% were heterozygous [CG], and 17.6% were homozygous for the variant allele [GG].
Focus on 10 Circulating MiRNAs with Possible Relevance to Inflammatory Liver Diseases
For this study, we selected a panel of 10 miRNAs: MiR-146b-5p, miR-4530, miR-4651 were chosen as we identified them to be upregulated by the inflammatory cytokine hyper-interleukin-6 (IL-6) in primary hepatocytes22 and (Figure A1); miR-146a-5p was included for comparison with miR-146b-5p. MiR-526a2 (equivalent to miR-518d-5p and miR-520c-5p) and miR-873-5p were included based on previous evidence suggesting their relevance in liver diseases.23^,^24 Based on preliminary unpublished data, we added miR-335-5p and miR-433-3p, which were well detectable in the supernatant of hepatocytes (Cq values < 30) and at increased concentration upon hyper-IL-6-stimulation. Our miRNA panel also included miR-122-5p and miR-21-5p, both well-studied miRNAs in the context of inflammation and liver diseases, (eg5^,^25, 26, 27).
Altered MiRNA Profiles in Sera from Patients with Chronic Liver Diseases
The serum levels of miR-335-5p, miR-433-3p, and miR-873-5p were decreased in HCC patients compared to healthy controls (Figure A2). When analyzing HCC subgroups, categorized according to the presence or absence of cirrhosis, circulating levels of miR-433-3p and miR-873-5p were higher in HCC patients with cirrhosis (HCC-Cirr) compared to those without (HCC-NC). In contrast, serum concentrations of miR-146a-5p were lower in HCC patients with cirrhosis (Figure 1).Figure 1. Profiles of 10 miRNAs in patients with chronic liver diseases. The horizontal line within the box plots represents the median, vertical lines from the boxes (whiskers) indicate the variability outside the upper and lower quartiles. Pairwise t-tests were performed, and P values were adjusted for each miRNA using the Holm method. ∗∗∗∗: P < .0001, ∗∗∗: P < .001, ∗∗: P < .01, ∗: P < .05.
In the MASLD patients, serum levels of miR-21-5p, miR-433-3p, miR-4530, and miR-4651 were decreased compared to healthy controls (Figure A2). The decrease of miR-4530 and miR-433-3p levels in the sera from MASLD patients (compared to the healthy controls) was significant only for patients with simple steatosis (MASLD-NC-Stea; Figure 1). Sera of patients with MASH (MASLD-NC-MASH) showed higher levels of miR-122-5p and of miR-21-5p as compared to patients with simple steatosis (MASLD-NC-Stea) and to patients with MASLD and cirrhosis (MASLD-Cirr). For miR-4651, both groups with complicated MASLD (MASH, cirrhosis) displayed higher serum levels than the simple steatosis group (Figure A3A). Similar findings (albeit with higher P values) were observed for miR-21-5p, miR-433-3p, miR-4530, and miR-526a2, while miR146a-5p and miR-146b-5p were lower in patients with complicated MASLD (Figure A3A). Notably, miR-146a-5p levels were significantly lower in patients with MASLD-cirrhosis compared to those with simple steatosis, similar to the observation for HCC patients, with or without cirrhosis (see above, Figure 1).
MiRNAs as Predictive Factors for Liver Diseases
We evaluated the suitability of the miRNAs in our panel to serve as discriminative markers in ROC curve analysis-based prediction models.
MiR-335-5p Discriminates Best HCC from Healthy Samples
MiR-335-5p, found at lower levels in HCC as compared to healthy samples (Figures 1, A2), was the only miRNA showing statistical significance to discriminate HCC cases from healthy individuals, with an area under the curve (AUC) value of 0.86 (Figure 2) and high scores for sensitivity and specificity (Table 2). However, it is to note that miR-335-5p does not outperform routine clinical parameters for liver damage, ie, gamma-glutamyl transferase (GGT) and aspartate amino transferase (AST), which were both detected at higher levels in sera from HCC patients in comparison to healthy controls (AUC values of 0.96; Table 2).Figure 2. MiR-335-5p as predictor of HCC vs healthy controls. The ROC curve for miR-335-5p is shown as well as the concentration plots for the healthy group (14 samples) and the HCC group (47 samples), indicating also the BH-adjusted P value of the GLM. GLM, generalized linear model.Table 2ROC Curve Analysesx (/x2)AUCCut-offPPVNPVSENSSPECEquationPrediction: HCC vs healthy (Figure 2) AST0.960.650.940.850.940.85y ∼ −26.29 + 19.01 ∗x GGT0.960.850.970.650.840.93y ∼ −9.65 + 6.40 ∗x PNPLA3 p.I148M “variant” ([CG] + [GG])0.680.870.870.410.720.64y ∼ 0.37 + 1.55 ∗x miR-335-5p0.860.710.930.670.890.77y ∼ −2.73 + −1.15 ∗xPrediction: MASLD-NC stea vs healthy (Figure 3A) miR-46510.950.730.970.720.880.93y ∼ −8.49 + −1.98 ∗x miR-21-5p0.890.740.970.620.790.93y ∼ −1.74 + −1.53 ∗x miR-45300.820.560.880.780.950.58y ∼ −9.78 + −1.19 ∗x miR-433-3p0.820.670.870.600.850.64y ∼ −2.71 + −0.91 ∗x miR-122-5p0.750.791.000.480.621.00y ∼ −0.58 + −0.48 ∗x miR-146b-5p0.680.830.940.350.380.93y ∼ 8.65 + 0.79 ∗xPrediction: MASLD vs healthy (Figure 3B) GGT0.940.790.950.760.840.93y ∼ −9.52 + 6.18 ∗x AST0.930.790.950.750.840.92y ∼ −21.31 + 15.49 ∗x miR-46510.900.880.980.450.770.93y ∼ −4.92 + −1.44 ∗x miR-21-5p0.840.891.000.340.611.00y ∼ −0.06 + −0.97 ∗xPrediction: MASLD-comp vs MASLD-NC-stea (Figures 4, A3B) miR-46510.720.390.640.790.770.68y ∼ 3.75 + 0.72 ∗x miR-4651/Fibroscan0.950.330.890.950.920.92y ∼ 13.49 + 1.57 ∗x + −0.01 ∗x2 miR-4651/Thrombocytes0.890.430.850.910.880.89y ∼ −5.96 + 0.85 ∗x + 11.72 ∗x2 miR-4651/GROα0.840.530.790.830.760.85y ∼ 28.08 + 0.67 ∗x + −10.62 ∗x2 miR-4651/HGF0.820.530.860.770.620.92y ∼ 6.37 + 0.78 ∗x + 2.27 ∗x2 miR-4651/miR-146a-5p0.810.570.860.770.630.92y ∼ 1.31 + 0.80 ∗x + 2.71 ∗x2 miR-4651/SCGFβ0.790.460.720.840.790.78y ∼ −5.34 + 0.71 ∗x + 4.80 ∗x2 miR-4651/CTACK0.780.370.680.830.790.72y ∼ −12.42 + 0.66 ∗x + 6.00 ∗x2 miR-4651/miR-146b-5p0.780.630.940.750.570.97y ∼ 8.95 + 0.93 ∗x + −1.10 ∗x2 miR-4651/IL-80.760.370.670.850.830.70y ∼ −0.06 + 1.20 ∗x + −0.96 ∗x2 Fibroscan0.920.400.880.880.810.92y ∼ −9.53 + 10.44 ∗x Thrombocytes0.870.420.880.890.850.91y ∼ 25.6 + −11.18 ∗x GROα0.810.540.90.790.660.95y ∼ −9.66 + 5.00 ∗x HGF0.770.600.940.750.550.98y ∼ −16.37 + 6.08 ∗x miR-146a-5p0.640.360.520.760.830.41y ∼ −3.3 + −0.45 ∗x SCGFβ0.710.350.60.850.860.58y ∼ −13.86 + 3.14 ∗x CTACK0.720.370.580.770.760.60y ∼ −6.91 + 2.74 ∗x miR-146b-5p0.640.460.620.730.670.69y ∼ −4.78 + −0.47 ∗x IL-80.660.410.640.70.550.78y ∼ −5.79 + 3.90 ∗xFor the ROC curve analyses shown in the indicated figures, the AUC values, positive predictive values, negative predictive values, and the values for sensitivity (SENS) and specificity (SPEC) are indicated. The cut-offs refer to the outcome of the regression (y). x, x2: independent parameters; y: binary outcome (0/1).SENS, sensitivity; SPEC, specificity; NPV, negative predictive value; PPV, positive predictive value; GROα, growth-regulated oncogene-alpha; HGF, hepatocyte growth factor; SCGFβ, stem cell growth factor-beta.
ROC Curve Analyses Identify MiR-4651 as a Potent Novel Predictor of MASLD
For six miRNAs, the prediction analyses to discriminate healthy from MASLD-NC-Stea (ie, simple steatosis) samples yielded significant results. MiR-4651 performed best, with an AUC value of 0.95 and high values for the positive and negative predictive values, sensitivity, and specificity (Figure 3A, Table 2). MiR-21-5p, miR-4530, and miR-433-3p follow, with AUC values of 0.89, 0.82, and 0.82, respectively. Notably, thrombocytes, as the clinical parameter performing best in this analysis, only yielded an AUC of 0.71 (data not shown).Figure 3ROC curve analyses for prediction of MASLD vs healthy controls. (A) Discrimination of simple steatosis from healthy samples. Left: ROC curves are shown for 6 miRNAs: miR-4651, miR-21-5p, miR-4530, miR-433-3p, miR-122-5p, and miR-146-5p. Right: Concentration plots for the 4 best performing miRNAs, for the healthy group (14 samples) and the MASLD-NC-Stea group (40 samples). The BH-adjusted P values of the GLMs are indicated. (B) miR-4651 and miR-21-5p are best predictors to differentiate (all) MASLD from healthy samples. Left: Plots of the relative levels of the 4 miRNAs with BH-adjusted P values of the GLMs <0.05: miR-4651, miR-21-5p, miR-4530, and miR-433-3p; the healthy group comprises 14 samples, the MASLD group 70 samples. Right: ROC curves for these 4 miRNAs. GLM, generalized linear model.
MiR-4651 also performed best in the ROC curve analyses for discrimination of (all) MASLD from healthy samples (AUC: 0.90), followed by miR-21-5p, miR-4530, and miR-433-3p, with AUC values of 0.84, 0.76, and 0.74, respectively (Figure 3B, Table 2). For this comparison, the clinical parameters GGT and AST yielded better prediction results (AUCs: 0.94 and 0.93, respectively; Table 2).
Interestingly, miR-4651 was the only miRNA predicting, within the MASLD group, “complicated” MASLD cases (MASLD-Comp) vs uncomplicated ones (MASLD-NC-Stea, ie, simple steatosis); Figure 4). The moderate AUC value of 0.72 could be improved by adding a second parameter into the model: including miR-146a-5p or miR-146b-5p into the ROC curve analyses led to increased AUC values of 0.81 and 0.78, respectively. Adding serum levels of inflammatory cytokines such as growth-regulated oncogene-alpha, hepatocyte growth factor, stem cell growth factor-beta, or CTACK increased the AUCs to reach values of 0.84, 0.82, 0.79, and 0.78, respectively. (As shown in our previous manuscript14 and in Table 2, growth-regulated oncogene-alpha was most effective among 22 cytokines in a single-parameter model to predict complicated MASLD (AUC: 0.78), followed by hepatocyte growth factor, stem cell growth factor-beta, and CTACK (Table 2). As expected, the combination of miR-4651 with thrombocytes or liver stiffness (quantified by transient elastography) led to very high AUC values of 0.95 and 0.89, respectively (Figures 4; A3B and Table 2 also present their excellent values in single-parameter models).Figure 4. Two-parameter models including miR-4651 for an improved discrimination of “MASLD complication” from simple steatosis. Top left: Plots of the relative levels of miR-4651 in the simple steatosis group (MASLD-NC-Stea, 40 samples) and in the “MASLD complication” group (MASLD-Comp: 30 samples: 9 for MASLD-NC-MASH, 21 for MASLD-Cirr); the BH-adjusted P value of the GLM is indicated. Top right: ROC curve for miR-4651. Lower panel: The addition of second parameters to the model leads to increased AUC values. Among the significant combinations, the 10 best AUC combinations are colored and labeled, the others are shown in gray. GLM, generalized linear model.
Effect of the PNPLA3 p.I148M Risk Allele on Levels of Circulating MiRNAs
We analyzed whether the PNPLA3 p.I148M genotype was associated with the levels of the serum miRNAs. We noted lower levels of miR-21-5p in carriers of the risk allele in the steatosis group, whereas for the other comparisons no differences were detected (Figure 5 and data not shown), although miR-122-5p has been previously reported to be present at higher concentrations in carriers of the risk allele.10 Of note, in all groups miR-4530 levels tended to be decreased in carriers of the risk allele (Figure 5).Figure 5. The PNPLA3 p.I148M risk allele does not substantially affect levels of circulating miRNAs. Upper panel: Lower levels of miR-21-5p in carriers of the variant allele ([CG] + [GG]) compared to the homozygous carriers of the WT allele [CC] in the steatosis group, ∗: P = .046. Lower panel: Tendency (but no statistical significance) of lower levels of miR-4530 in carriers of the variant allele in all groups.
Discussion
In this study we show that 1) miR-4651 is the best predictor for MASLD. In particular, this miRNA can differentiate individuals with simple steatosis from healthy ones, it also emerged as the best predictor for “MASLD complication” vs simple steatosis; 2) miR-335-5p is present at lower levels in patients with HCC compared to healthy controls; and 3) the PNPLA3 p.I148M risk genotype does not seem to strongly affect serum levels of the 10 miRNAs.
MiR-4651 as a Novel MiRNA with Biomarker Potential for MASLD
MiR-4651 is downregulated in the sera of MASLD patients compared to healthy controls whereas no significant difference was observed between the HCC and healthy samples (Figures 1, A2). Of note, levels are particularly low for the MASLD-NC-Stea (ie, simple steatosis) group, with higher levels for the MASLD-NC-MASH and MASLD-Cirr groups. In ROC curve analyses, miR-4651 performed best in discriminating 1) healthy samples from MASLD-NC-Stea samples (Figure 3A), 2) healthy samples from (all) MASLD samples (Figure 3B), and 3) samples from MASLD patients with simple steatosis from those with complication (i.e., MASH/cirrhosis) (Figure 4). Of note, miR-4651 performed better than the “best” clinical parameter (thrombocytes) to predict simple steatosis vs healthy controls. In contrasts, for other prediction models (healthy vs MASLD, simple steatosis vs MASLD complication) the clinical parameters GGT and AST or “Fibroscan” and “thrombocytes” were more powerful than any of the miRNAs (Table A1, Figure A3B). In fact, combinations of miR-4651 with the parameters “Fibroscan” or “thrombocytes” led to the highest AUC levels in the model differentiating MASLD without complication (ie, simple steatosis) from MASLD with complication.
Only a few studies have reported on this miRNA so far. MiR-4651 was one of well-detectable serum miRNAs in a 8-miRNA panel able to predict HCC in patients with various liver diseases.6 In agreement with our data, Yamamoto et al. found that miR-4651 levels were reduced in a large cohort of patients with chronic hepatitis/liver cirrhosis compared to healthy controls and to HCC patients. Moreover, miR-4651 was reported to be a biomarker in HCC cases related to aflatoxin; serum levels of miR-4651 were positively related to the concentration of aflatoxin B1-albumin adduct levels.7 While Sun et al. did not detect differences in the sera of patients with cirrhosis compared to healthy controls,28 miR-4651 was previously found to be upregulated in the plasma of chronically hepatitis B virus-infected patients with fibrosis stages S1 to S4 compared to those with S0.29 Moreover, miR-4651 was part of a panel of 9 serum miRNAs that could serve as early biomarkers for new-onset type-2 diabetes, with higher levels in patients than in healthy controls.30 Expression of miR-4651 was found to be decreased in HCC tissues compared to adjacent normal tissues, and patients with higher expression had a better survival rate. It suppresses the growth of HCC by targeting the expression of the transcription factor FOXP4.31 High tissue levels of miR-4651 were predictive of a better response to transarterial chemoembolization in HCC patients,32 possibly linked to expression of its target CYP2W1, a member of the cytochrome P450 superfamily. Taken together, follow-up studies are warranted to better define the role of this miRNA in liver diseases and its potential as a biomarker.
MiR-335-5p Levels are Decreased in HCC
Serum levels of miR-335-5p were significantly lower in HCC patients, regardless of cirrhosis status, compared to healthy controls (Figures 1, A2). In a prediction model, miR-335-5p performed well in discriminating between HCC and healthy controls (AUC: 0.86, Figure 2). In line with our results, Elfert et al. reported decreased serum levels of miR-335 in patients with hepatitis C virus (HCV) (compared to normal controls), especially those with HCV-related HCC. Indeed, this miRNA had a high prognostic power in distinguishing HCC patients from nonmalignant HCV patients or healthy controls.33 Also Cui et al. found serum levels of miR-335-5p to be reduced in HCC patients compared to healthy controls and hepatitis patients; higher miR-335 concentrations were associated with a better treatment response to transarterial chemoembolization and with a better prognosis of HCC patients.34 Decreased levels of this miRNA were observed in extracellular vesicles isolated from sera of patients with severe liver injury-chronic hepatitis B and decompensated cirrhosis.35
MiR-335-5p has tumor suppressive functions in several types of cancer, including HCC (see36 for a review), inhibiting expression of proteins like ROCK1 or MAPK1, implied in the regulation of cellular migration and/or proliferation.37^,^38 M**iR-335 is located within an intron of MEST, a gene presenting hypermethylation of its promoter in HCC,39 which may explain the lower expression of miR-335-5p in tumor tissue compared to the surrounding nontumor tissue.37, 38, 39 Overexpression of miR-335-5p or application of miR-335-5p via exosomes decreased proliferation rates of hepatoma cells, both in vitro and in vivo.40 Indeed, an exosome-based anticancer drug using miR-335 (next to tumor necrosis factor-related apoptosis inducing ligand) was started to be developed for treatment of HCC.36
Results of Other MiRNAs in the Context of Previous Studies
We included into our study miR-122-5p, the most abundant miRNA in the liver, as well as the inflammation-associated miR-21-5p, both well-studied miRNAs in the context of liver diseases and robustly detectable in serum (eg5). In HCC samples, we observe only a trend toward higher levels of both miRNAs than in healthy controls (Figure 1). Several studies, however, described significantly higher levels in the blood of HCC patients, of miR-122 (eg26^,^27^,^33) or/and of miR-21-5p (eg26^,^41), whereas no differences were noted in other studies.27^,^42 (For miR-122-5p, we found higher levels in MASLD-NC-MASH samples compared to those from MASLD-NC-Stea and MASLD-Cirr samples. As this miRNA was previously found to be decreased in high grades of fibrosis (eg5^,^43), it was stated that this miRNA could “be postulated as a biomarker for nonalcoholic steatohepatitis, but it does not seem useful as a biomarker for advanced fibrosis”.5 Higher concentrations of circulating miR-122-5p in MASH samples compared to simple steatosis samples were also reported by others (eg25^,^44^,^45). However, in contrast to several other (eg25^,^44, 45, 46, 47) but not all48^,^49 studies, we did not detect significant differences between (all) MASLD and healthy samples in our cohort (Figure A2). Of note, our MASLD cohort is dominated by individuals with simple steatosis (40, ie, 57%, vs 9 with MASH and 21 with cirrhosis). Interestingly, Auguet et al. (2014) and Pirola et al. (201544; validation cohort) found no differences between healthy and simple steatosis, whereas miR-122 levels were significantly higher in sera from patients with steatohepatitis.44^,^50
MASLD-NC-MASH samples contained significantly higher levels also of miR-21-5p compared to the MASLD-NC-Stea and MASLD-Cirr samples (Figure 1). Moreover, we also observed a higher concentration in “MASLD-complication” samples compared to simple steatosis (Figure A3A). Higher serum levels of miR-21-5p in MASH samples compared to simple steatosis have also been noticed by others.25^,^51 It should be noted, however, that the overall levels of miR-21-5p were lower in MASLD samples compared to the healthy samples of our cohort (Figure A2). This is in line with a study by Sun et al.52 whereas other studies noted no difference46^,^47 or higher levels in MASLD (eg25^,^26).
MiR-873-5p and miR-526a2 (sequence identity with miR-518d-5p) were included in the study because of their relevance to liver diseases: miR-873-5p is found to be upregulated in liver tissues of MASLD patients, an important target of miR-873-5p being the mitochondrial glycine-N-methyltransferase, thus affecting the methionine cycle and the activity of the electron transport chain.23 MiR-526a2 was described to be upregulated in the tissues and sera of HCC patients and to exert antiapoptotic effects in HCC cells.24 While we only see a trend towards higher levels of serum miR-873-5p during MASLD progression (Figures 1, A2, A3A), miR-526a is higher expressed in “complicated MASLD” compared to simple steatosis (Figure A3A). MiR-873-5p is present at lower levels in sera of HCC patients compared to healthy controls while in our study no significant difference was found for miR-526a2 (Figures 1, A2).
In our previous microarray analysis, miR-4530 and miR-4651 were among the miRNAs that were upregulated in primary human hepatocytes upon stimulation with hyper-IL-6,22 a finding that we could confirm by qPCR validation, including further primary hepatocyte samples (Figure A1). Interestingly, miR-4530 is present at higher concentration in the sera of MASLD patients “with complication” compared to those with simple steatosis (Figure A3A). Overall, we observe lower levels miR-4530 in MASLD patients compared to healthy controls (Figure A2A), which is driven by a lower concentration in patients with simple steatosis (Figure 1). HCC sera did not present different levels of miR-4530 compared to healthy control samples. Of note, in a study with 86 HCC patients, those with a higher serum level of miR-4530 had better survival rates compared to those with lower levels.53
MASLD and HCC patients with cirrhosis had reduced levels of serum miR-146a-5p (Figure 1). Interestingly, this microRNA was also observed to be reduced in hepatic fibrosis due to schistosomiasis, with lower serum levels in higher grades of fibrosis.54 Also Blaya et al. found the circulating levels of miR-146a-5p to be lower in patients with cirrhosis compared to healthy controls, with a further decrease in patients with decompensated cirrhosis or acute-on-chronic liver failure and correlating with patient outcomes.55
No Major Impact of the PNPLA3 p.I148M Risk Genotype on the Serum Levels of the 10 MiRNAs
The PNPLA3 p.I148M allele is one of the strongest genetic risk factors for MASLD and for other liver diseases. Interestingly, we did not observe significant differences between carriers of the risk allele and those with homozygosity of the WT allele (exception: lower levels of miR-21-5p in the sera of MASLD-NC-Stea patients with the risk allele, Figure 5). Our study therefore does not confirm findings by Ezaz et al.,10 who found the GC and GG variants at rs738409 of PNPLA3 to be robustly associated with higher levels of circulating miR-122-5p in a cohort of 132 MASLD patients. Of note, carriers of the PNPLA3 p.I148M risk allele had consistently lower median values regarding the serum level of miR-4530 compared to those carrying only the WT allele (Figure 5).
Conclusion
It would be important to validate our findings in an independent study cohort with a larger number of enrolled individuals, including a larger healthy control group (ours comprised 14 participants only), to increase statistical power. Moreover, the suitability of miR-4651 as biomarker in the diagnosis of simple steatosis should be followed up. A limited number of molecules have been tested for their diagnostic potential to detect simple steatosis: Soluyanova et al. identified a set of 5 serum miRNAs which could predict liver fat content with high sensitivity and specificity.56 Moreover, serum concentrations of the liver-produced glycoprotein afamin correlate with hepatic fat contents and could therefore serve as an early marker of metabolic syndrome.57 For prediction of MASH and MASLD-cirrhosis, more powerful clinical markers, as single parameters, outperformed the miRNAs of our panel. Yet, we provide evidence that (selected) miRNAs may be valuable analytes for inclusion in multiparameter prediction models, to complement imaging techniques and serum markers as well-established noninvasive diagnostic tools for MASLD.1^,^58
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Huang D.Q.Wong V.W.S.Rinella M.E.Metabolic dysfunction-associated steatotic liver disease in adults Nat Rev Dis Primers 112025144005036210.1038/s 41572-025-00599-1 · doi ↗ · pubmed ↗
- 2Singal A.G.Kanwal F.Llovet J.M.Global trends in hepatocellular carcinoma epidemiology: implications for screening, prevention and therapy Nat Rev Clin Oncol 2020238648843788473610.1038/s 41571-023-00825-3 · doi ↗ · pubmed ↗
- 3Llovet J.M.Kelley R.K.Villanueva A.Hepatocellular carcinoma Nat Rev Dis Primers 7202163347922410.1038/s 41572-020-00240-3 · doi ↗ · pubmed ↗
- 4Harrison S.A.Ratziu V.Magnanensi J.NIS 2+, an optimisation of the blood-based biomarker NIS 4(R) technology for the detection of at-risk NASH: a prospective derivation and validation study J Hepatol 7920237587673722492310.1016/j.jhep.2023.04.031 · doi ↗ · pubmed ↗
- 5Lopez-Riera M.Conde I.Tolosa L.New micro RNA biomarkers for drug-induced steatosis and their potential to predict the contribution of drugs to non-alcoholic fatty liver disease Front Pharmacol 8201732817988310.3389/fphar.2017.00003 PMC 5263149 · doi ↗ · pubmed ↗
- 6Yamamoto Y.Kondo S.Matsuzaki J.Highly sensitive circulating Micro RNA panel for accurate detection of hepatocellular carcinoma in patients with liver disease Hepatol Commun 420202842973202561110.1002/hep 4.1451 PMC 6996324 · doi ↗ · pubmed ↗
- 7Wu X.M.Xi Z.F.Liao P.Diagnostic and prognostic potential of serum micro RNA-4651 for patients with hepatocellular carcinoma related to aflatoxin B 1Oncotarget 8201781235812492911338310.18632/oncotarget.16027 PMC 5655278 · doi ↗ · pubmed ↗
- 8Baselli G.A.Dongiovanni P.Rametta R.Liver transcriptomics highlights interleukin-32 as novel NAFLD-Related cytokine and candidate biomarker Gut 692020185518663200155410.1136/gutjnl-2019-319226 PMC 7497582 · doi ↗ · pubmed ↗