Two new species of terrestrial microsnails of the genus Hypselostoma W.H. Benson, 1856 (Gastropoda, Eupulmonata, Hypselostomatidae) from northeastern Thailand

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
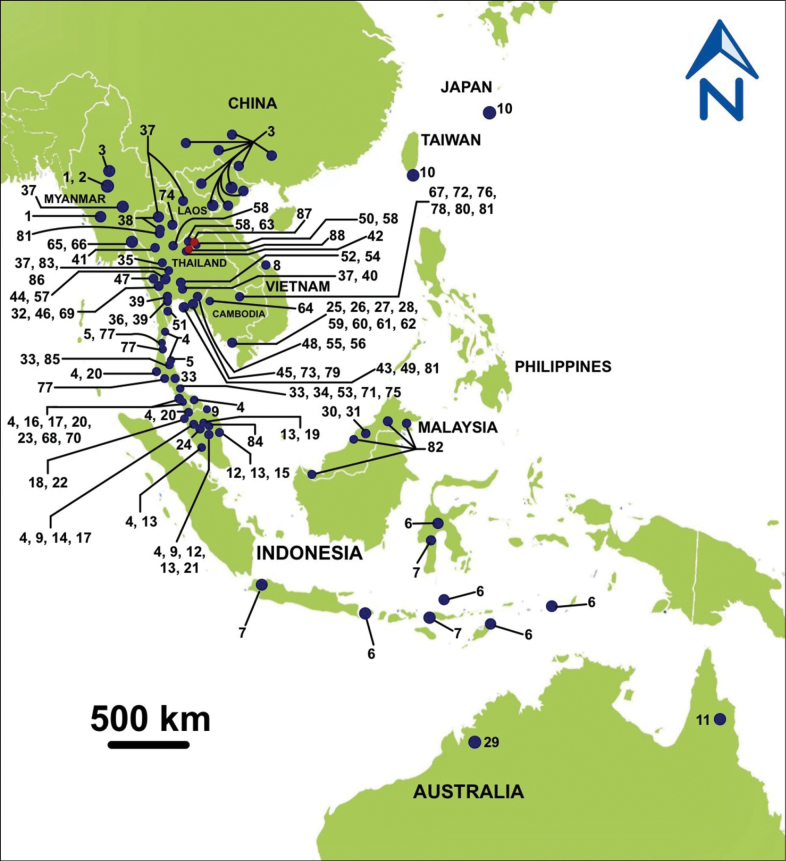 Figure 1
Figure 1 Figure 2
Figure 2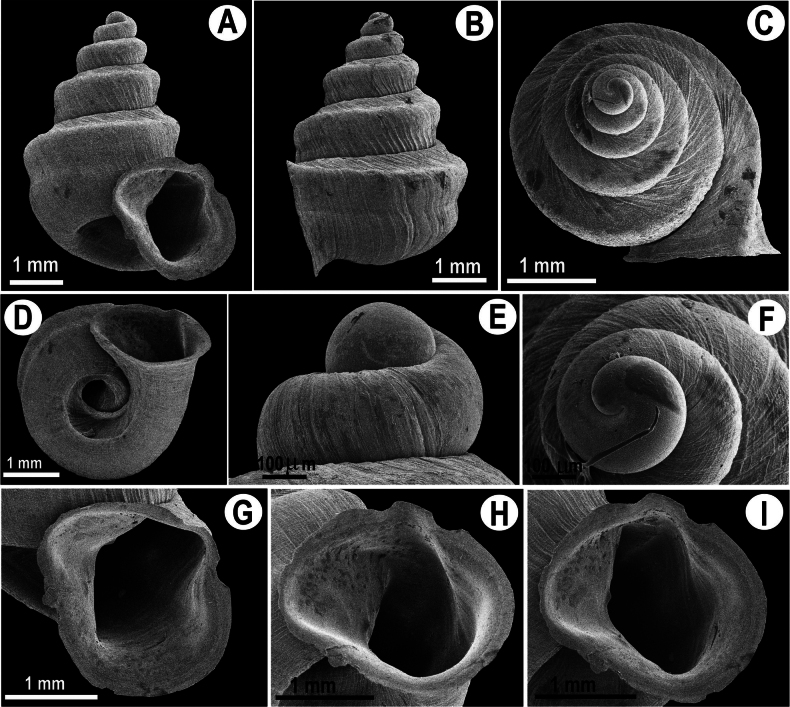 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5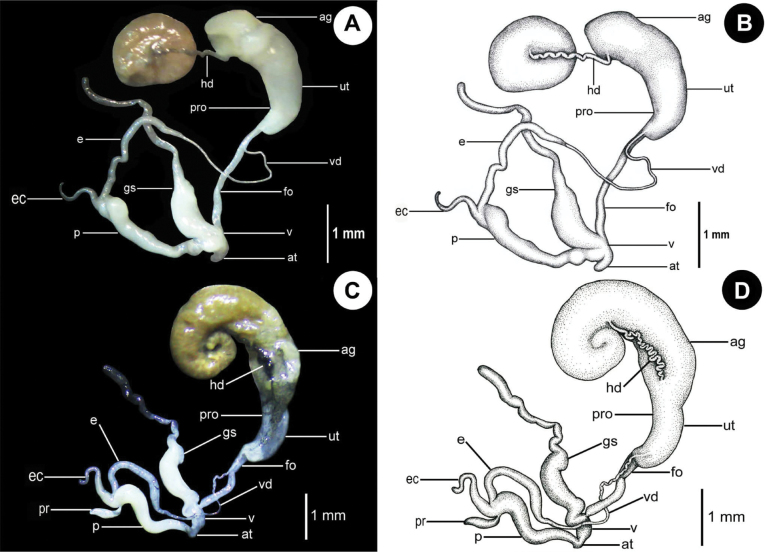 Figure 6
Figure 6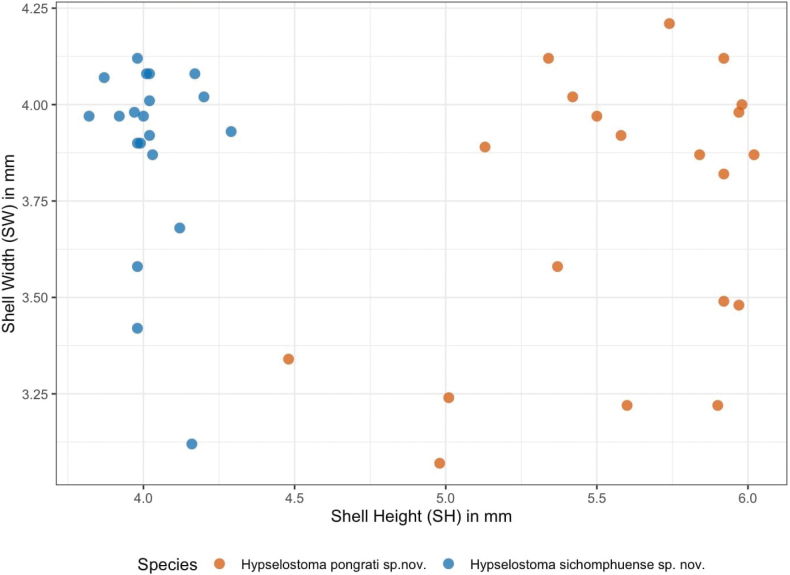 Figure 7
Figure 7| S/N | Species name | Localities | References |
|---|---|---|---|
| 1 | Thyet-Mio, Irawadi Burmanici, Myanmar | 3 | |
| 2 | Mandalay Division, Myanmar | 1, 3, 4, 18 | |
| 3 | Lang-son, Tonkin, Vietnam; Tianxingqiao Town, Zhenning Bouyeizu Miaozu Zizhixian, Guizhou, China; Nawit Village, Viengxay District, Houaphane, Laos | 3, 6, 25 | |
| 4 | Bukit Baling, Kedah; Gunong Pondok, Perak; Batu Caves, Selangor; Bukit Chintamani and Gua Bama, Pahang, Malaysia; Kuankalong Limestone Hill, Satun; Samui Islands, Thailand | 3, 7, 8, 29 | |
| 5 | Samui Islands, Surat Thani and Chumphon, Thailand | 3, 25, 26, 33 | |
| 6 | Timor Island, Kalao Island, Sulawesi, Java, Bali, Lesser Sunda Islands and Tanimbar Islands, Indonesia | 3, 21, 33 | |
| 7 | West Java Province, Djampang, Java, and Makalé to Kalossi, Sulawesi, South Celebes, Indonesia | 3, 30 | |
| 8 | Phuc-Son, Touranne, Annam, Vietnam | 3 | |
| 9 | Kelantan and Kota Tongkat, Pahang, and Kramat Pulai, Perak, Malaysia | 3 | |
| 10 | Yonakunijima, Ryuyku, Japan; Kenting Tropical Botanical Garden, Hengchun, Pingtung, Taiwan | 3 | |
| 11 | Chillagoe Caves, Queensland, Australia | 3, 22 | |
| 12 | Bukit Chintamani, Pahang, and Bukit Charas, Pahang, and Bukit Takun, Kanching, Selangor, Malaysia | 3 | |
| 13 | Bukit Chintamani; Gua Bama, Pahang, Bukit Panching, Pahang Malaysia | 3, 7, 29 | |
| 14 | Sungei Siput, Perak, Malaysia | 3 | |
| 15 | Bukit Tenggek, Pahang, Malaysia | 3 | |
| 16 | Bukit Chuping, Perlis, Malaysia | 3, 7, 28 | |
| 17 | Kota Tongkat, Pahang and Gunong Batu Kurau, Perak, Malaysia | 3 | |
| 18 | Gunong Pondok, Padang Rengas, Perak, Malaysia | 3, 7, 32 | |
| 19 | Gua Musang, Kelantan and Peninsular Siam, Malaysia | 3, 7, 32 | |
| 20 | Baling, Kedah, Malaysia; Tarutoa National Park, Satun and Krabi, Thailand | 3 | |
| 21 | Gua Bama, Padang Tengku, Pahang, Malaysia | 3, 7, 28 | |
| 22 | Gunong Pondok, Padang Rengas, Perak, and Kota Tongkat, Pahang, Malaysia | 3 | |
| 23 | Kampong Tebing Tinggi, Kangar, Perlis, Malaysia | 3, 31 | |
| 24 | Bukit Serdam, Pahang, Malaysia | 3 | |
| 25 | Kampot, Cambodia; Kien Giang, Vietnam | 3 | |
| 26 | Binh An Commune, Ba Nui Village, Mo So Cave, Kien Luong District Kien Giang, Vietnam | 3 | |
| 27 | Kien Giang, Vietnam; Kampot, Cambodia | 3 | |
| 28 | Phum Troung Mean, Banan District, Battambang, Cambodia | 3, 23 | |
| 29 | Yammera Gap and Napier Range, Kimberley Region, Western Australia, Australia | 3, 22 | |
| 30 | Deer Cave, Melinau Paku Valley, Fourth Division, Sarawak, Borneo, Malaysia | 3 | |
| 31 | Gunong Budah, Medalam Valley, Fifth Division, Sarawak, Borneo, Malaysia | 3 | |
| 32 | Kanchanaburi Agricultural College, Mueang District, Kanchanaburi, Thailand | 3 | |
| 33 | Ban Na San District, Surat Thani, Thailand | 17, 25 | |
| 34 | Phatthalung, Surat Thani, and Phetchaburi, Thailand | 3, 11, 17 | |
| 35 | Tebpratan Nature Reserve area, Kamphaeng Phet, Thailand | 3 | |
| 36 | Kangkrajan National Park, Phetchaburi, Thailand | 3 | |
| 37 | Takli District, Nakhon Sawan; Tepitak Tamaram Temple, Tepitak Mountain, Mueang Saraburi District, Saraburi; Muangon Cave, San Kam Pang District, Chiang Mai, Thailand | 3, 13, 17 | |
| 38 | Tamphatai National Park, Ngao District, Lampang; Ban Pa Ngae Limestone Knoll, Padaet District, Chiang Rai, Thailand | 3 | |
| 39 | Khaochongpran, Tao Pun Sub-District, Photharam District, Ratchaburi, Thailand | 3 | |
| 40 | Tepitak Mountain, Muaklek District, Saraburi, Thailand | 3, 15, 17 | |
| 41 | Chonglom Mountain, Bhumiphol Dam Reservoir, Samngao District, Tak, Thailand | 3 | |
| 42 | Phuphaman Mountain, Phetchabun, Thailand | 3 | |
| 43 | Sichang Island, Si Racha District, Chonburi, Thailand | 3, 14, 17 | |
| 44 | Tam Sua Temple, Utong District, Suphanburi, Thailand | 3 | |
| 45 | Pluangthong Mountain, Botong District, Chonburi, Thailand | 2, 3, 17 | |
| 46 | Erawan National Park, Sai Yok District, Kanchanaburi, Thailand | 3 | |
| 47 | Chongkhaokad, Sai Yok District, Kanchanaburi, Thailand | 3 | |
| 48 | Chakan Mountain, Khaochakan Temple, Khaochakan District, Sa Kaeo, Thailand | 2, 3, 17 | |
| 49 | Kohrin (Rin Island), Chonburi, Thailand | 2, 3, 17 | |
| 50 | Puttabanpot Temple, Nawang District, Nong Bua Lamphu, Thailand | 2, 3, 17 | |
| 51 | Central Area of Tamrong Temple, Mueang Phetchaburi District, Phetchaburi, Thailand | 3 | |
| 52 | Khaowongkot, Ban Mi District, Lopburi, Thailand | 3, 16, 17 | |
| 53 | Limestone Hill in Ko Si Ko Ha (Ko Na Thewada), Phatthalung, Thailand | 3 | |
| 54 | Smokon Mountain, Ban Mi District, Lopburi, Thailand | 3 | |
| 55 | Plubpluengtong Limestone Hills, Sa Kaeo, Thailand | 3, 23 | |
| 56 | Plubpluengtong Limestone Hills, Sa Kaeo, Thailand | 3, 16, 17 | |
| 57 | Tam Sua Hill, Utong District, Suphanburi, Thailand | 3 | |
| 58 | Limestone Hills in Loei, Phitsanulok, Khon Kaen and Nong Bua Lamphu, Thailand | 3 | |
| 59 | Phnom Kampong Trach, Kampot, Cambodia | 3 | |
| 60 | Kampong Trach Area, Phnom Kampong Trach, Kampot, Cambodia | 3 | |
| 61 | Phnom La’Ang Cave, Kampot Area, Kampot, Cambodia | 3 | |
| 62 | Phnom La’Ang, Kampot Area, Kampot, Cambodia | 3 | |
| 63 | Agricultural Areas in Mueang Loei District, Loei; Lampang and Chonburi, Thailand | 3, 24 | |
| 64 | Phnom Sampeov Mountain, Banan District, Battambang, Cambodia | 3, 23 | |
| 65 | Kaw Gon Cave, Hpa-An, Kayin State, Myanmar | 3, 27 | |
| 66 | Pan Do Mi Mountain, Phon Tho Village, Kamarmaung, Kayin State, Myanmar | 3 | |
| 67 | Chap Phleung Mountain, Stung Treng Mekong Bridge, Steung Treng, Cambodia | 3 | |
| 68 | Kodiang, Gua Kerbau, Kedah, Malaysia | 3 | |
| 69 | Taoist Buddhist Temple, Kanchanaburi, Thailand | 3 | |
| 70 | Tanjung Rhu Beach, Langkawi Island, Malaysia | 3 | |
| 71 | Khao Ok Thalu, Phattalung; & Songkhla, Thailand | 3 | |
| 72 | Stung Treng Mekong Bridge, Phnom Chhnok, Stung Treng , Cambodia | 3 | |
| 73 | Tham Khao Wongkot, Kaeng Hang Maeo District, Chanthaburi, Thailand | 3 | |
| 74 | Ban Pha Tun, Tham Pha Tup Forest Park, Mueang Nan District, Nan, Thailand | 3 | |
| 75 | Khao Ok Thalu, Phatthalung, Thailand | 3 | |
| 76 | Stung Treng Mekong Bridge, Chap Phleung Mountain, Steung Treng, Cambodia | 3 | |
| 77 | Krabi, Chumphon and Ranong, Thailand | 3 | |
| 78 | Stung Treng Mekong Bridge, Phnom Chhnok, Steung Treng, Cambodia | 3 | |
| 79 | Wat Tham Khao Prathun, Khao Chamao District, Rayong, Thailand | 3 | |
| 80 | Stung Treng Mekong Bridge, Chap Phleung Mountain, Steung Treng, Cambodia | 3 | |
| 81 | Stung Treng Mekong Bridge, Chap Phleung Mountain, Steung Treng, Cambodia | 3 | |
| 82 | Gunong Subis; Sarawak & Sabah in Borneo, Malaysia | 3 | |
| 83 | Nakhon Sawan, Thailand | 3 | |
| 84 | Gua Telinga, Kuala Tahan, Taman Negara, Jerantut, Pahang, Malaysia | 3 | |
| 85 | Surat Thani, Thailand | 3 | |
| 86 | Ban Non San Village, Nakhon Sawan, Thailand | 3 | |
| 87 | Si Bun Rueang District, Nong Bua Lamphu, Thailand | This study | |
| 88 | Si Chomphu District, Khon Kaen, Thailand | This study |
| Characters | Species | |||
|
|
| |||
| Shell: | ||||
| Shape | Conical | Concave-conical | Concave-conical | Conical-ovoid |
| Shell height (mm) | 4.48–6.02 | 3.82–4.29 | 2.11–2.25 | 1.87–3.17 |
| Shell width (mm) | 3.07–4.12 | 3.12–4.12 | 2.50–2.80 | 1.74–3.28 |
| Appearance of the last whorls | double keeled | double keeled | keeled at the center of the periphery | keeled above the center of the periphery |
| Apertural barrier | absent | 3 | 26 | absent |
| Radula Formula | (7-8):5:1:5:(7-8) | (9-10):7:1:7:(9-10) | 8:5:1:5:8 | (7-8):4:1:4:(7-8) |
| Genital system: | ||||
| Penial retractor muscle (pr) | absent | short tube | Bulged and curved | Short and rounded |
| Gametolytic sac | anterior portion bulge, larger than penis | anterior portion bulge, equal to penis | anterior portion bulge smaller than penis | anterior portion bulge, smaller than penis |
| distal end long cylindrical and curved. | distal end long cylindrical and curved | distal end bulged | posterior portion long slender, curved | |
| References | This study | This study |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMollusks and Parasites Studies · Aquatic Invertebrate Ecology and Behavior · Invertebrate Taxonomy and Ecology
Introduction
The land pulmonate microsnails of the genus Hypselostoma W.H. Benson, 1856, were classified into the superfamily Pupilloidea W. Turton, 1831 and family Hypselostomatidae Zilch, 1959 (Gojšina et al. 2025), based on their shell morphology, especially apertural barriers (van Benthem Jutting 1950; Schileyko 1998; Panha and Burch 2008; Sutcharit et al. 2023). This genus has high-spired, conical shells with a free trumpet-shaped last whorl. Panha and Burch (2008) used the separation of the angular and parietal lamellae within the aperture as the key character to distinguish Gyliotrachela Tomlin, 1930 from Hypselostoma. Recently, Gojšina et al. (2025) treated Gyliotrachela and Antroapiculus Panha & J.B. Burch, 2002 as junior synonyms of Hypselostoma, because there are several instances of species from both genera having very similar, if not identical, shell morphologies, while the presence of parietal angular lamellae alone would not justify the separation of the two genera. In their taxonomic revision, Gojšina et al. (2025) concluded that the updated number of species of Hypselostoma is now 95 species (85 verified species + 10 unverified species from the Philippines), and that this genus is widely distributed in Southeast Asia and north and west Australia, Japan, and China (van Benthem Jutting 1950, 1962; Solem 1981; Schileyko 1998; Panha et al. 2004; Panha and Burch 2008; Tongkerd et al. 2013; Gojšina et al. 2025) (Table 1, Fig. 1).
Map of Southeast Asia and the surrounding area showing the type localities of the known Hypselostoma spp. The numbers correspond to the species numbers listed in Table 1.
The two new species of Hypselostoma described in this paper were recently discovered on a wall in limestone hills covered with mixed-deciduous forest in Nong Bua Lamphu and Khon Kaen Provinces, northeastern Thailand. The external shell morphology of the two new species is very similar to that of Hypselostoma phupaman (Panha & J.B. Burch, 2002c) from Chaiyaphum Province (Panha and Burch 2008), with which they share the double-keeled last whorl, but from which they differ by their number of apertural barriers. These three species are also similar to Hypselostoma torta Gojšina, Auffenberg & Páll-Gergely, 2025 from Nakhon Sawan Province (Gojšina et al. 2025) with which they share their shell shape, as well as the strongly shouldered penultimate and last whorls. However, H. torta differs as it has only one keel on the last whorl, instead of two.
Materials and methods
Microsnail specimens were collected during November–December 2021 and January 2022 from limestone hill in Nong Bua Lampu Province (Figs 1, 2A, B), and Khon Kaen Province (Figs 1, 2C, D), northeastern Thailand. Complete adult shells were used to count the number of whorls and measure shell height (SH), shell width (SW), aperture height (AH), and aperture width (AW) with digital vernier calipers (Electronic Digital Calliper S.H.). Shells and radulae were photographed by a scanning electron microscope (LEO 1450 VP) at the Microscope Center, Faculty of Science, Burapha University and Centre for Scientific and Technological Equipment at Suranaree University of Technology. Adult microsnails were dissected under a stereomicroscope to examine their genital system and extract their radula from the buccal mass. The radula sac was soaked in 5% NaOH solution to separate and clear the radula. Species identification was based on Pilsbry (1916–1918), van Benthem Jutting (1950), Panha and Burch (2008), and Gojšina et al. (2025). Shell characters were examined following Panha and Burch (2008) and Gojšina et al. (2025), whereas characters of genitalia and radula were studied following Dumrongrojwattana and Tanmuangpak (2020) and Tanmuangpak and Dumrongrojwattana (2022).
Living Hypselostoma spp. A, B.Hypselostoma pongrati sp. nov. paratype NHLRU015; C, D. Hypselostoma sichomphuense sp. nov. paratype NHLRU033.
The type material was deposited in the following institutions: NHLRU Natural History Museum of Loei Rajabhat University (Loei, Thailand) and NHMSU Natural History Museum, Mahasarakham University (Maha Sarakham, Thailand).
Results
Taxonomy
Family Hypselostomatidae Zilch, 1959
Hypselostoma
Taxon classificationAnimaliaEupulmonataHypselostomatidae
Genus
W.H. Benson, 1856
86B5930D-29B4-5365-B27A-C01885FAD0ED
Diagnosis.
Shell tiny, turbinate, high-spired, or discoidal. Peristome adnate or separated from the adjacent body whorl forming a tuba; edge of peristome expanded. Aperture with four or more parietal and angular apertural lamellae or plicae (barriers), except in Hypselostoma edentatum (Panha & J.B. Burch, 2002d) and H. pongrati sp. nov. which have no apertural barriers and H. sichomphuense sp. nov. which has only three barriers (Panha and Burch 2008; Gojšina et al. 2025). The genitalia of Hypselostoma possess a penis that is more swollen than the epiphallus; gametolytic sac connects to vagina very close to atrium, divided into two parts, with the proximal part swollen and larger than the penis (except in H. cultura (Tanmuangpak & Dumrongrojwattana, 2022), in which the gametolytic sac has a long duct between proximal and distal part, while there is no swollen proximal part in H. depressispira (Berry, 1963)). These anatomical characters differ from the genus Aulacospira Möllendorff, 1890, whose gametolytic sac connects to the vagina far from the atrium (Dumrongrojwattana and Tanmuangpak 2020)
Hypselostoma
pongrati
Taxon classificationAnimaliaEupulmonataHypselostomatidae
Tanmuangpak & S. Tumpeesuwan sp. nov.
095F7E0C-6B70-5DEC-8E9E-F734D112E162
https://zoobank.org/C545C51C-2464-468F-983B-9713B28FEFBC
Figs 2A, B, 3A–I, 5A–E, 6A, B; Table 2
Hypselostoma sp. – Tanmuangpak & Kaewsawang, 2025: 20, 23, 25, 27, 29–31, fig. 2q.
Material examined.
Holotype: Thailand • 1 empty shell; Nong Bua Lamphu Province, Sri Bun Rueang District, Pha Sam Yod limestone hill (17°10'02.020"N, 102°02'03.020"E) covered by mixed-deciduous forest; 20.xi.2021, leg. Tanmuangpak, K. (NHLRU014) (Fig. 3A–E). Paratypes: Thailand • 12 specimens preserved in 70% ethanol (NHLRU015–026); • five shells (NHLRU027–031); • three specimens preserved in 70% ethanol (NHMSU–00063–B); • two shells (NHMSU–00063). All paratypes collected from the same location as holotype, leg. Tanmuangpak, K. 10.xii.2021.
Table 2.: Comparison of shell, radula, and genital system of Hypselostoma pongrati sp. nov., H. sichomphuense sp. nov., H. cultura and Aulacospira vanwalleghemi.
Shell morphology of Hypselostoma pongrati sp. nov. holotype NHLRU014. A. Apertural view; B. Lateral view; C, E, F. Protoconch view; D. Umbilical view apertural detail; G–I. Apertural view in different angles.
Measurements.
Holotype: SH = 4.48 mm, SW = 3.34 mm, AH = 1.95 mm, AW = 1.92 mm. Paratypes (19 specimens measured): SH = 4.48–6.02 mm (5.58 ± 0.42 mm), SW = 3.07–4.12 mm (3.72 ± 0.36 mm), AH = 1.95–2.64 mm (2.24 ± 0.20 mm), AW = 1.68–2.42 mm (1.99 ± 0.16 mm) (Fig. 7).
Diagnosis.
This new species has a prominent keel on the upper part of the last whorl and a weaker keel on the lower part. Apertural barriers absent (Fig. 3A, G, H, L). Terminal part of the last whorl is adnated to penultimate whorl.
Description.
Shell (Fig. 3A–I) concave-conical, high spired, with 4¾ whorls. Large size for the genus, with shell height 4.48–6.02 mm, shell width 3.07–4.12 mm. The suture is deep. Protoconch smooth, with 1–1½ whorls that gradually increase in size to the teleoconch whorls. Teleoconch with 3–3¾ whorls, very fine spiral striation the body whorl widest, possesses two keels (upper one more prominent than the lower one). Apertural barriers absent. Peristome thickened and expanded. The parietal and convex angulo-palatal embayments form a sinulus, thickened, and expanded. The basal peristome is smooth and expanded, columellar side is more expand than the angulo-palatal side.
Genital system (Fig. 6A, B). Atrium shorter than the vagina. Penis shorter than the epiphallus, its anterior portion is a short and bulged tube, and the distal end of the penis bulge. Epiphallus connects to the distal end of the penis, shorter than vas deferens, anterior portion slender and cylindrical, its central portion more slender than the anterior portion, posterior portion curved, white glossy. Penial retractor muscle absent. Vas deferens long, slender, entering the epiphallus apically. The vagina and free oviduct are cylindrical, and the vagina is shorter than the free oviduct. The gametolytic sac is a long tube, with a swollen proximal part, and larger than penis, surrounded by a thin sheath and connected to the proximal part of the vagina, whose distal end is long, slender, and curved. Uterus large, with very thin prostate gland attached to it. Hermaphroditic duct loosely convoluted. The albumen gland large and yellowish. Dart apparatus absent.
Radula (n = 4) (Fig. 5A–E). Radula comprises 173–184 (179 ± 4.93) transverse rows of teeth each row containing 25–27 teeth. Radula formula (7-8)+5+1+5+(7-8). Central tooth small, unicuspid, elongated triangular. Lateral teeth bicuspid and asymmetrical, consisting of a large lanceolate endocone and smaller elongated triangular ectocone. Five lateral teeth on each side of the central tooth, the first tooth largest, and gradually smaller outwards. Marginal teeth start at 6^th^ tooth outwards from central teeth, which almost changed to tricuspids. On each outer side of the lateral teeth there are seven or eight marginal teeth.
Etymology.
This new species is dedicated to the late Pongrat Dumrongrojwattana, our highly respected senior who was an expert on microsnails and who provided us with many land snails references, knowledge, and inspiration.
Animal and ecology.
Living animals have a cream-colored body and foot, while their head and tentacles are rather black, with black eyes located at the tip of the ocular tentacle. This species was found on the limestone wall in the mixed-deciduous forest (Fig. 2A, B).
Distribution.
This species was found only on limestone wall in Pha Sam Yod, Si Bun Rueang District, Nong Bua Lamphu Province.
Remarks.
Hypselostoma pongrati sp. nov. from Si Bun Rueng District, Nong Bua Lam Phu Province, is quite different in shell shape from almost all other Hypselostoma species in Thailand. It is similar in shell shape to the geographically close H. phupaman from Chaiyaphum Province and H. sichomphuense sp. nov. from Si Chomphu, Khon Kaen Province. The unique characteristics of H. pongrati sp. nov. are the lack of apertural barriers and the last whorl adnate to the penultimate whorl, peristome thick, expanded and not reflected. It is strongly expanded at the parietal side where it leans against penultimate whorl and forms a thick crescent callus. Three species possess a double-keeled last whorl, but the lower keel is weak in both H. pongrati sp. nov. and H. sichomphuense sp. nov. Other similar species are H. torta from Nakhon Sawan Province, Thailand and H. platybasis from Steung Treng Province, Cambodia (Gojšina et al. 2025). In H. torta and H. pongrati sp. nov. all teleoconch whorls are shouldered, but in H. torta the last whorl is detached from the penultimate whorl. There are four weak apertural barriers. The columellar barriers form as a rather strong swelled part of the aperture which forms a prominent basal furrow below it. Hypselostoma platybasis differs from H. pongrati sp. nov. by possesses bluntly keeled penultimate at the center of the periphery and shouldered last whorl. Last whorl very slightly detached from the penultimate whorl and with a sharp shoulder. Apertural barriers few and relatively weak. In H. pongrati sp. nov. there is no penial retractor muscle, and the free oviduct is longer than the anterior portion of the gametolytic sac. Conversely, in H. sichomphuense sp. nov. there is a penial retractor muscle, and the free oviduct is shorter than the anterior portion of the gametolytic sac.
Hypselostoma
sichomphuense
Taxon classificationAnimaliaEupulmonataHypselostomatidae
Tanmuangpak & S. Tumpeesuwan sp. nov.
2F106A50-5E7D-525E-8B8B-6631EFACD4B5
https://zoobank.org/EAE8D792-E347-4F2C-9BDB-2291A398D89C
Figs 2C, D, 4A–I, 5F–L, 6C, D; Table 2
Material examined.
Holotype: Thailand • 1 empty shell Khon Kaen Province, Si Chomphu District, limestone wall in mixed-deciduous forest, Phu Pha Kham (16°50'22.001"N, 102°03'59.001"E), 21.xi.2021, leg. Tanmuangpak, K. (NHLRU032) (Fig. 3G–K). Paratypes: Thailand • 12 specimens preserved in 70% ethanol (NHLRU033–044) (Fig. 2C, D); • two shells (NHLRU045–046); • four specimens preserved in 70% ethanol (NHMSU–00064-B); • one shell (NHMSU–00064). All paratypes collected from the same location as holotype, leg. Tanmuangpak, K. 11.xii.2021.
Measurements.
Holotype: SH = 4.20 mm, SW = 4.02 mm, AH = 2.14 mm, AW = 2.24 mm. Paratypes: (19 specimens measured). SH = 3.82–4.29 mm (4.03 ± 0.11 mm), SW = 3.12–4.12 mm (3.88 ± 0.25 mm), AH = 1.84–2.27 mm (2.02 ± 0.12 mm), AW = 1.67–2.24 mm (2.01 ± 0.13 mm) (Fig. 7).
Diagnosis.
This new species differs from Hypselostoma phupaman by the presence of a non-prominent lower keel on the last whorl and the absence of basal and parietal apertural barriers. Aperture has three smooth tooth-like swellings (Fig. 4A, G–I), whereas there are no apertural barriers in H. pongrati sp. nov. (Fig. 3A, G–I). Spire shorter than the spire of H. pongrati sp. nov.
Shell morphology of Hypselostoma sichomphuense sp. nov. holotype NHLRU032. A, G–I. Apertural view; B. Lateral view; C, E, F. Protoconch view; D. Umbilical view; A, G–I. Apertural view in different angles. Abbreviations for apertural teeth: UPL = upper palatal plica; C = columellar fold; IP = infraparietal plica.
Radula morphology of Hypselostoma spp. A–E.Hypselostoma pongrati sp. nov. paratype NHLRU015; F–L.Hypselostoma sichomphuense sp. nov. paratype NHLRU034. Abbreviations: C = central tooth; L = lateral teeth; M = marginal teeth.
Description.
Shell (Fig. 4A–I) concave-conical, wide-spired, with 4½ whorls. Large size for the genus, with shell height 3.82–4.29 mm, shell width 3.12–4.12 mm. The suture is deep. Protoconch pitted, without spiral pattern, with one whorl gradually increasing in size to the teleoconch whorls. The teleoconch has 3½ whorls, a very fine spiral surface, with growth lines. The body whorl widest, double keeled (prominent upper and weak lower keel). The peristome is thickened and expanded. Aperture has three smooth, tooth-like swellings, viz. an upper palatal plica, a columellar fold, and a infraparietal plica.
Genital system (Fig. 6C, D). Atrium is shorter than the vagina. Penis is longer than the epiphallus, its anterior portion is a short and gradually bulged tube, and the distal end of its posterior portion is bulged as well. Epiphallus is connected to the distal end of the penis. Epiphallus shorter than vas deferens, its anterior portion slender and cylindrical, its central portion gradually bulges, and its posterior portion is a slender tube, white and glossy. Epiphallic caecum is a short tube, attached to the anterior portion of epiphallus. The penial retractor muscle is shorter than the epiphallus, attached to the distal end of the penis. Vas deferens long, slender, entering epiphallus apically. The vagina and free oviduct are cylindrical, and the vagina is shorter than the free oviduct. The gametolytic sac is a long tube, its bulging anterior portion is surrounded by a thin sheath and connected to vagina, the distal end is a long slender and curved tube. The uterus is large, with a very thin prostate gland attached to it. The hermaphroditic duct is loosely convoluted. The albumen gland is yellowish and large. Dart apparatus absent.
Genital system of Hypselostoma spp. A, B.Hypselostoma pongrati sp. nov., paratype (NHLRU015); A. Genital system; B. Schematic drawing of the genital system; C, D.Hypselostoma sichomphuense sp. nov., paratype (NHLRU034); C. Genital system; D. Schematic drawing of the genital system. Abbreviations: ag = albumen gland; at = atrium; e = epiphallus; ec = epiphallic caecum; fo = free oviduct; hd = hermaphroditic duct; p = penis; pr = penial retractor muscle; pro = prostate; gs = gametolytic sac; ut = uterus; v = vagina; vd = vas deferens.
Shell dimension of Hypselostoma pongrati sp. nov. and Hypselostoma sichomphuense sp. nov.
Radula (n = 3) (Fig. 5F–L). Radula comprises of 154–165 (159 ± 5.56) rows of teeth, each row with 33–35 teeth. Radula formula: (9–10) +7+1+7+ (9–10). Central tooth small, unicuspid, and triangular. Lateral teeth bicuspid and asymmetrical, consisting of a large endocone and a smaller ectocone. Seven lateral teeth on each side of the central tooth; the first tooth is largest, and the other teeth are sequentially smaller. Marginal teeth are irregular, unequally tricuspid, with an endocone larger than the ectocone and gradually change to polycuspid outwards (Fig. 5I–L). There are nine or ten marginal teeth on each outer side of the lateral teeth (Fig. 5F–L).
Etymology.
The specific epithet sichomphuense refers to Si Chomphu District, Khon Kaen Province, northeastern Thailand, where this species was discovered.
Animal and ecology.
Living animals have cream-colored bodies, with pale brown head and tentacles and black eyes at the tip of the ocular tentacle. This species was found on the limestone wall covered by mixed-deciduous forest (Fig. 2C, D).
Distribution.
Limestone wall at Phu Kham, Si Chomphu District, Khon Kaen Province.
Remarks.
Hypselostoma sichomphuense sp. nov. is quite different in shell shape from all other Hypselostoma species in Thailand, except for the geographically close H. pongrati sp. nov. The unique characteristics of H. sichomphuense sp. nov. are the three tooth-like swellings in the aperture and the last whorl adnate to the penultimate whorl.
Discussion
The general shape of the radula teeth of the two new species is very similar to that of Hypselostoma cultura, but their formulae are different: (7–10):(5–7):1:(5–7):(7–10) in the new species versus 8:5:1:5:8 in H. cultura (Tanmuangpak and Dumrongrojwattana 2022).
Compared with the genital systems of H. cultura (see Tanmuangpak and Dumrongrojwattana 2022) and Aulacospira spp. (see Dumrongrojwattana and Tanmuangpak 2020), these two new species have a large cylindrically shaped penis, which is relatively long and whose distal end bulges. The epiphallus is more elongated with a cylindrical shape but shorter than the penis. The gametolytic sac is very long, the anterior to central portion bulged, equal to larger than penis and connected to the vagina, and the central to posterior portion is slenderer and long, with the distal end curved.
Phylogenetic analysis of DNA sequence data of some Asian land microsnails showed the incongruence between shell-based taxonomy and DNA-based phylogenetic relationships (Chiba 1999; Tongkerd et al. 2004a; Hirano et al. 2014). For example, DNA-based phylogenic analysis of Thai microsnails revealed that Gyliotrachela, Hypselostoma and Anauchen are paraphyletic, e.g., Aulacospira smaesarnensis Panha & Burch, 2001 nested within Gyliotrachela (Tongkerd et al. 2004a). In this phylogenetic tree, the leaf litter inhabiting H. panhai, with its last whorl adnated to the penultimate whorl, whereas the limestone-dwelling H. erawan as sister group has the last part of last whorl detached from the penultimate whorl. Tongkerd et al. (2004a) suggested that the loss of this ancestral shell character state may have been driven by selective pressures in the new leaf litter habitat. These results suggest that the use of apertural dentition is not suitable for diagnostic generic characters because of ecological transitions can lead to morphological change. In this study, the species with a double-keeled last whorl seem to have fewer apertural barriers, ranging from six in H. phupaman (Panha & Burch, 2008) to none in H. pongrati sp. nov. The reduction of apertural barriers may be due to decrease in predation pressure by specific predators (Páll-Gergely et al. 2022), which is an interesting point for further research.
In the period 2002–2023, many new species of land snails were described from the isolated limestone hills in Chaiyaphum-Khon Kaen-Nong Bua Lamphu-Loei Areas, for example, Hypselostoma phupaman (Panha & J.B. Burch, 2002); H. surakiti (Panha & J.B. Burch, 2003); H. loei Panha & Prateespasen, 2005; Phuphania costata C. Tumpeesuwan & S. Tumpeesuwan, 2014; Sinoennea loeiensis Tanmuangpak & S. Tumpeesuwan, 2015 in Tanmuangpak et al. (2015); Sesara triodon Tanmuangpak & Tumpeesuwan, 2016 in Tanmuangpak et al. (2016); Aenigmatoconcha clivicola C. Tumpeesuwan & S. Tumpeesuwan, 2017; Landouria strobiloides C. Tumpeesuwan & S. Tumpeesuwan, 2019; Landouria circinataNahok et al., 2021; L. tuberculataNahok et al., 2021; L. trochomorphoidesNahok et al., 2021; L. chloritoidesNahok et al., 2021; and L. elegansNahok et al., 2021; H. cultura Tanmuangpak & Dumrongrojwattana, 2022; and Rhiostoma ebenozostera Tongkerd & Panha, 2023 in Tongkerd et al. (2023). These new species described from this limestone area confirm that this area is very important for biodiversity conservation and needs continuous studies and protection.
Supplementary Material
XML Treatment for Hypselostoma
XML Treatment for Hypselostoma pongrati
XML Treatment for Hypselostoma sichomphuense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Benson WH (1856 h) Description of Tanystoma tubiferum, a Burmese form related to the genus Anostoma of Lamarck.Annals & Magazine of Natural History 17(98): 129–131. 10.1080/00222935608697483 · doi ↗
- 2Benson WH (1856 i) Remarks on the genera Tanystoma, Nematura, and Anaulus.Annals & Magazine of Natural History 17(100): 342–343. 10.1080/00222935608697520 · doi ↗
- 3Berry AJ (1963) The genital system of the Malayan hill snail Gyliotrachela depressispira, with notes on breeding.Proceedings of the Zoological Society of London 141(2): 361–369. 10.1111/j.1469-7998.1963.tb 01616.x · doi ↗
- 4Blanford WT (1863) Contributions to Indian malacology. No. IV. Descriptions of new land shells from Ava, and other parts of Burma.Journal of the Asiatic Society of Bengal 32: 319–327.
- 5Burch JB Panha S Tongkerd P (2003) New taxa of Pupillidae (Pulmonata: Stylommatophora) from Thailand. Walkerana 13(29/30): 129–187.
- 6Chiba S (1999) Accelerated evolution of land snals Mandarina in the oceanic Bonin Islands: Evidence from mitochondrial DNA sequences.Evolution International Journal of Organic Evolution 53(2): 460–471. 10.1111/j.1558-5646.1999.tb 03781.x 28565404 · doi ↗ · pubmed ↗
- 7Dumrongrojwattana P Tanmuangpak K (2020) The terrestrial microsnail genus Aulacospira Möllendorff, 1890 (Eupulmonata, Stylommatophora, Hypselostomatidae) in Thailand with key to Thai species.Zoo Keys 980: 23–42. 10.3897/zookeys.980.5410033192136 PMC 7642177 · doi ↗ · pubmed ↗
- 8Gojšina V Hunyadi A Sutcharit C Tongkerd P Auffenberg K Grego J Vermeulen JJ Reischütz APáll-Gergely B (2025) A new start? Revision of the genera Anauchen, Bensonella, Gyliotrachela and Hypselostoma (Gastropoda, Eupulmonata, Hypselostomatidae) of Southeast Asia with description of 46 new species.Zoo Keys 1235: 1–338. 10.3897/zookeys.1235.14528140321358 PMC 12044344 · doi ↗ · pubmed ↗