Dynamic transcriptomic landscape of myogenesis in Muscovy ducks (Cairina moschata): integrative analysis of hub genes post-hatching
Xiaofeng Li, Kewei Fan, Bing Yang

TL;DR
This study identifies key genes involved in muscle development in Muscovy ducks after hatching, offering insights for improving meat production.
Contribution
The study presents the first integrative transcriptomic analysis of post-hatching myogenesis in Muscovy ducks, identifying ten hub genes.
Findings
1,683 differentially expressed genes were identified between 1-day-old and 80-day-old Muscovy duck muscles.
Ten hub genes, including CD3E, ITK, COL4A6, and IVD, were prioritized as key regulators of myogenesis.
Functional analysis linked these genes to metabolic processes like L-leucine catabolism and fatty acid β-oxidation.
Abstract
Post-hatching myogenesis is a critical determinant of meat yield and quality, with potential regulatory roles of specific genes remaining underexplored in Muscovy ducks (Cairina moschata). This study aimed to identify hub genes governing post-hatching myogenesis through transcriptomic profiling. Three white-feathered male Muscovy ducks at 1-day-old (1D) and 80-day-old (80D) were selected, respectively. Following humane euthanasia, the left leg muscles were collected for subsequent RNA extraction, library construction, genome alignment, and transcriptome sequencing. Differential gene expression analysis was performed using DESeq2 (ver. 1.16.1), with significant differentially expressed genes (DEGs) selected under the thresholds of |log2FC|>1 and p-value<0.05. Subsequent functional characterization included enrichment analyses (Gene Ontology, KEGG, and PANTHER pathways) implemented…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
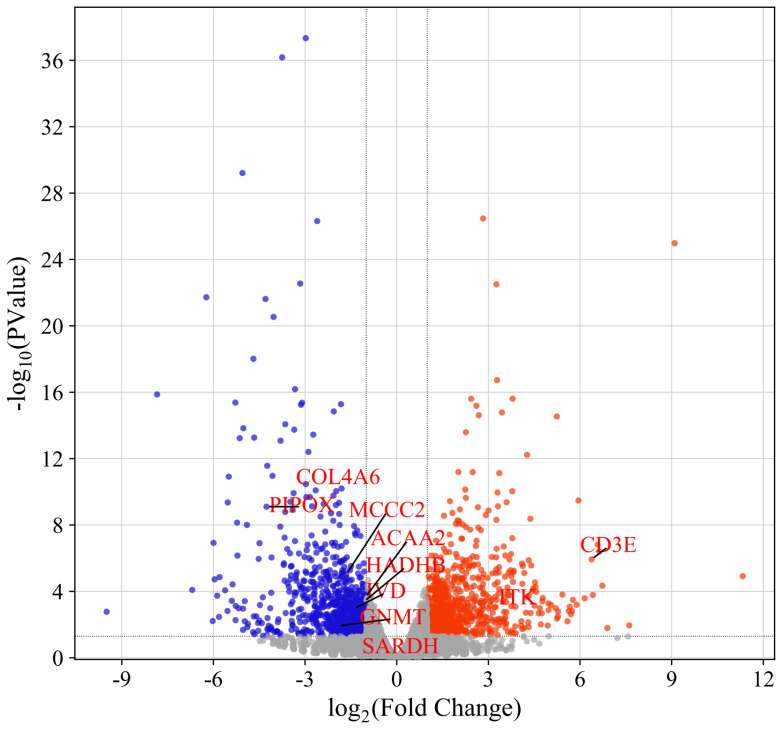 Figure 1
Figure 1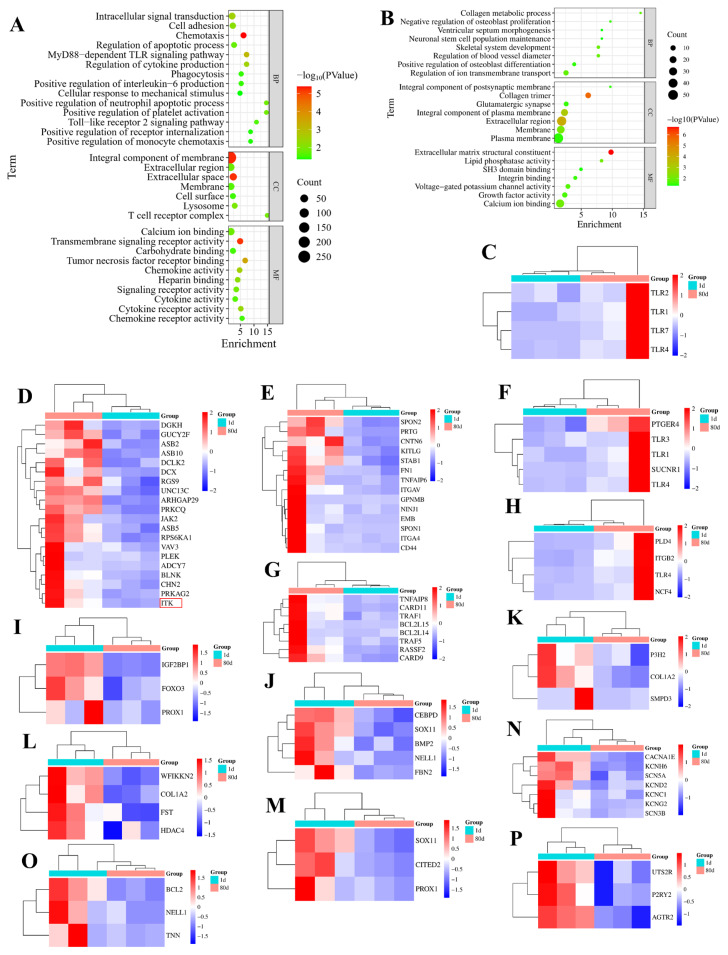 Figure 2
Figure 2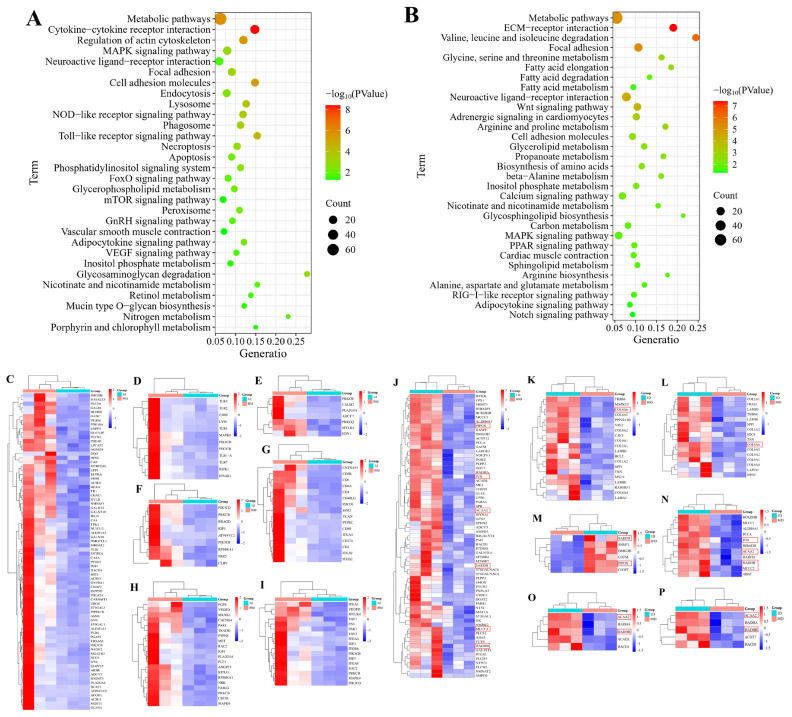 Figure 3
Figure 3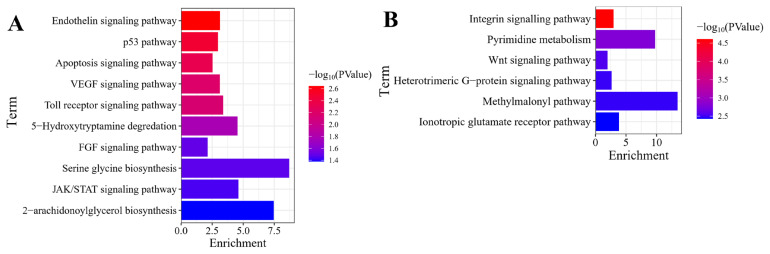 Figure 4
Figure 4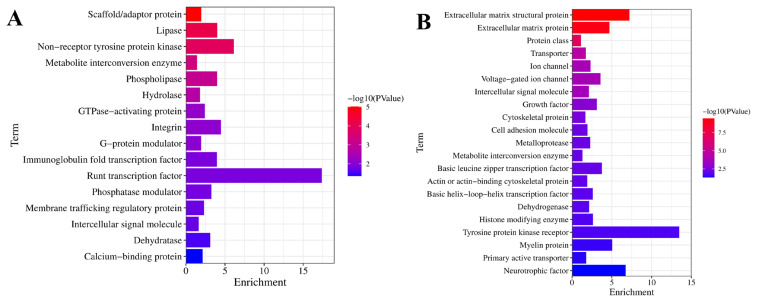 Figure 5
Figure 5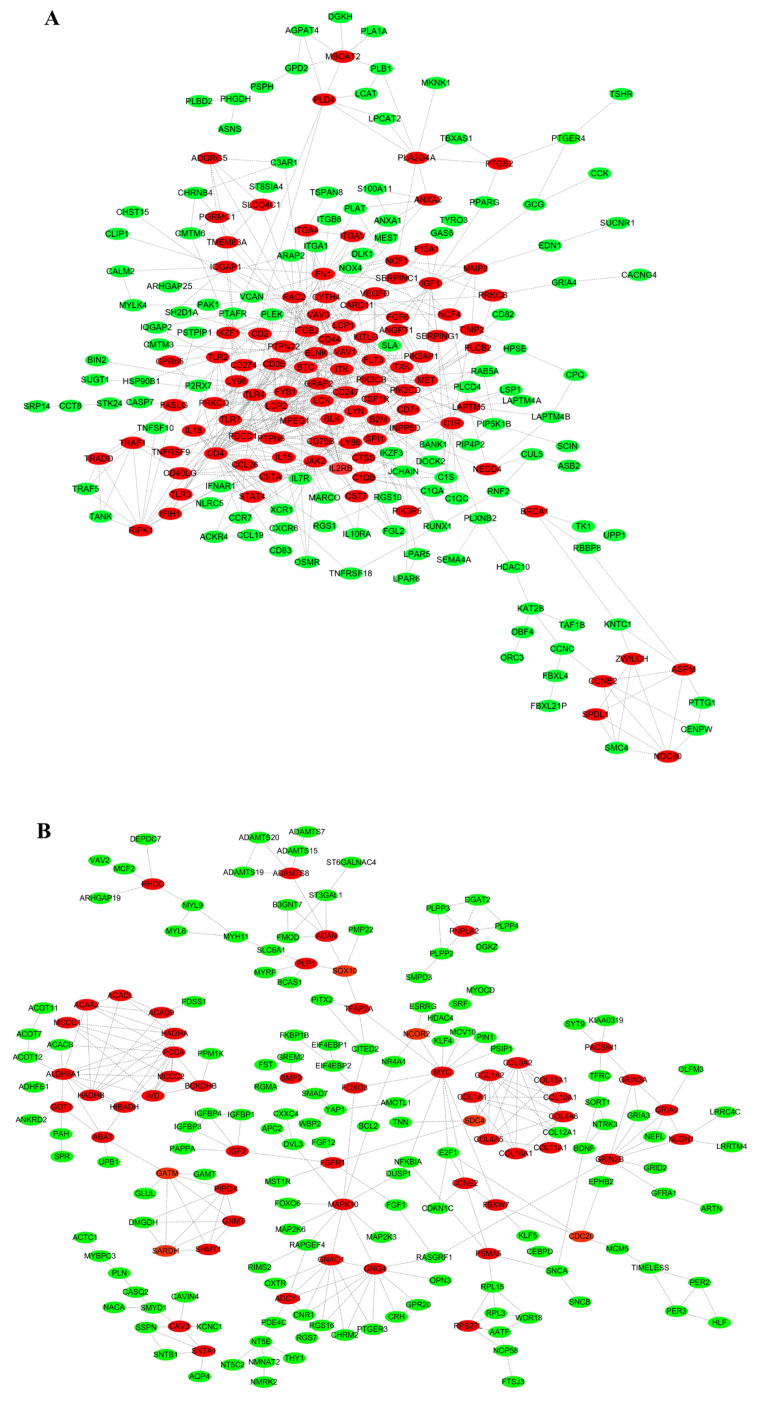 Figure 6
Figure 6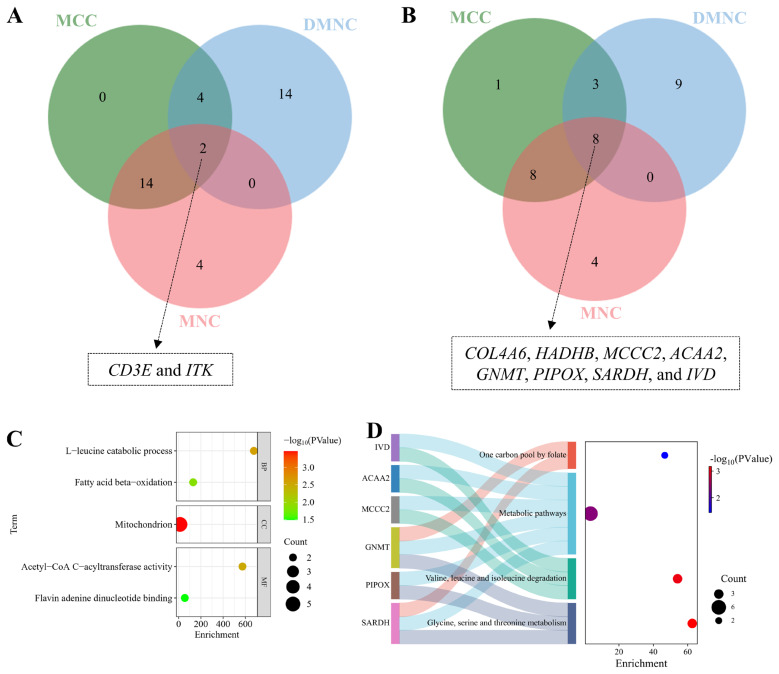 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMuscle Physiology and Disorders · Animal Genetics and Reproduction · GDF15 and Related Biomarkers
INTRODUCTION
Post-hatching skeletal myogenesis serves as a fundamental determinant of both meat yield and quality in poultry production systems, with significant implications for economic viability and consumer acceptance [1,2]. In avian species, this developmental process encompasses precisely regulated phases of myoblast proliferation, differentiation, and hypertrophic expansion - all orchestrated through complex transcriptional cascades and metabolic reprogramming. Notably, Muscovy ducks (Cairina moschata) exhibit exceptional muscular development characteristics, achieving 28%–32% leg muscle yield coupled with remarkably low fat deposition (10%–15%), thereby outperforming conventional poultry species (chickens, ducks, and geese) in leanness metrics. The functional importance of leg muscles in Muscovy ducks extends beyond meat production, as these muscles are biomechanically crucial for locomotion and weight-bearing, further underscoring their significance as key targets for genetic and nutritional optimization strategies [3]. Muscovy duck muscle development progresses through four key phases: 1) neonatal (0–7 days): hyperplasia establishes basic structures; 2) rapid growth (1–4 weeks): hypertrophy begins with 3–5×protein deposition; 3) maturation (4–8 weeks): hypertrophy dominates as slow-twitch fibers increase; 4) stabilization (>8 weeks): growth slows, fibers stabilize, reaching slaughter age at ~80 days. The development of key meat quality attributes, including tenderness and intramuscular fat deposition, is intrinsically associated with the coordinated regulation of myogenic processes and metabolic pathways. Notably, branched-chain amino acid catabolism coupled with fatty acid β-oxidation serves dual roles: providing energetic substrates for muscle growth while simultaneously regulating satellite cell dynamics and proteosynthetic efficiency [4,5].
Despite advancements in mammalian models, the molecular mechanisms underlying post-hatching myogenesis in Muscovy ducks remain poorly understood, limiting the development of precision breeding strategies to enhance meat traits. Addressing this gap is urgent, as global demand for high-quality poultry protein continues to escalate amid challenges of sustainability and resource efficiency. Recent transcriptomic studies in poultry have revealed both conserved regulators of myogenesis (e.g., MYOD1, MSTN) and species-specific metabolic adaptors [6]. In Muscovy ducks, rapid post-hatching muscle growth is hypothesized to reflect evolutionary adaptations to high-energy demands and environmental stressors. Emerging evidence highlights the critical roles of genes involved in amino acid degradation (e.g., IVD, MCCC2) and lipid metabolism (e.g., HADHB, ACAA2) in maintaining energy homeostasis during muscle hypertrophy [7–10]. Intriguingly, immune-related gene CD3E was associated with meat quality by regulating meat color, pH and muscle fat content [11,12]. Furthermore, GNMT and SARDH, implicated in folate-mediated one-carbon metabolism [13,14], could modulate epigenetic landscapes to influence myogenic transcription factor expression. Despite these insights, the functional hierarchy of hub genes coordinating metabolic reprogramming during duck myogenesis remains unresolved.
Through an integrative approach combining transcriptomic profiling (RNA-seq) and protein interactome mapping, this investigation delineates key regulatory hubs governing leg muscle development during the post-hatching phase of White Muscovy ducks. Through comparative transcriptomic profiling of male ducks at two critical developmental stages - initial post-hatching (1-day-old, ID) and mature growth phase (80-day-old, 80D) - we identify stage-specific transcriptional regulators and delineate their functional contributions to metabolic adaptation during muscle development. The findings provide dual scientific value: (1) advancing fundamental knowledge of molecular mechanisms underlying avian muscle growth, and (2) offering practical biomarkers for optimizing meat production efficiency in commercial duck farming. By establishing connections between transcriptomic dynamics and metabolic control networks, this research contributes to sustainable solutions for meeting global protein demands through precision poultry breeding approaches.
MATERIALS AND METHODS
Animal and muscle sample collection
Six white-feathered male Muscovy ducks at 80D and 1D were purchased from Anqing Yongqiang Agricultural Technology. respectively. All animals accessed to water and feed ad libitum. Prior to muscle sample collection, the ducks were euthanized humanely via intravenous injection of pentobarbital sodium at a dose of 150 mg/kg body weight. Approximately 4 grams of muscle tissue were harvested from the left leg of each duck. The samples were then rinsed with RNase-free water and promptly frozen in liquid nitrogen for subsequent analysis.
RNA isolation and quality assessment
Muscle tissue-derived total RNA was isolated utilizing Trizol reagent (Invitrogen) according to the manufacturer’s protocol. RNA integrity assessment was performed on a Bioanalyzer 2100 system (Agilent Technologies) equipped with RNA Nano 6000 assay kit, while purity evaluation was conducted via NanoPhotometer spectrophotometry (IMPLEN). Three representative RNA samples per group meeting stringent quality criteria (OD260/280 ratio>1.8, RIN>7.0) were subsequently processed for transcriptomic analysis.
Library preparation and sequencing
RNA sequencing libraries were prepared from 1 μg qualified RNA per sample using NEBNext UltraTM RNA Library Prep Kit for Illumina (NEB), with unique barcodes assigned for sample multiplexing. Cluster generation was achieved on cBot platform (TruSeq PE Cluster Kit v3-cBotHS; Illumina), followed by 150 bp paired-end sequencing on Illumina HiSeq 2000 system (BGI).
Primary data processing
Sequencing raw data underwent initial quality control through CASAVA pipeline (ver. 1.8.2; Illumina) for FASTQ conversion. Adapter sequences, poly-N reads, and low-quality bases (Q<20) were filtered using custom Perl scripts. Data quality metrics including Q20/Q30 scores, error rates, and GC content were systematically evaluated prior to downstream analysis.
Genome alignment
Clean reads were aligned to the Muscovy duck reference genome (NCBI Assembly ID: 1498951) using HISAT2 (ver. 2.0.5) with default parameters. The reference genome index was pre-built to ensure efficient and accurate mapping of sequencing reads.
Differential expression profiling
Gene expression quantification was performed via FeatureCounts (ver. 1.5.0) with FPKM normalization. Statistical analysis of differentially expressed genes (DEGs) between developmental stages was conducted using DESeq2 (ver. 1.16.1), applying negative binomial distribution modeling with Benjamini-Hochberg correction (|log2FC|>1, p<0.05).
Functional annotation
DEGs were subjected to comprehensive functional annotation through DAVID platform (ver. 6.8), including Gene Ontology (GO) term enrichment and KEGG pathway analysis. Complementary classification was performed using PANTHER database ( http://pantherdb.org) for protein categorization.
Interaction network construction
Protein-protein interaction networks were generated from DEG sets using STRING (ver. 12.0) with high-confidence interaction scores (>0.7). Network visualization and topological analysis were implemented in Cytoscape environment (ver. 3.8.0).
Hub gene identification and functional annotation
The identification of myogenesis-related hub genes was achieved through comprehensive topological analysis of the protein-protein interaction network using CytoHubba (ver. 3.8.0), a Cytoscape plugin implementing three distinct centrality algorithms: maximal clique centrality (MCC) for identifying pivotal nodes within maximal fully-connected subnetworks, maximum neighborhood component (MNC) for detecting locally dense connectivity modules, and density of maximum neighborhood component (DMNC) quantifying network compactness through edge density ratios. A consensus set of high-confidence hub genes was derived from the intersection of these algorithmically independent predictions, ensuring methodological robustness. Functional characterization of these candidate regulators involved systematic literature mining complemented by cross-referencing authoritative databases including NCBI (https://www.ncbi.nlm.nih.gov/) and GeneCards (https://www.genecards.org/), with particular emphasis on their documented roles in skeletal muscle development processes.
RESULTS
Transcriptomic profiling of muscovy duck muscle development
High-throughput sequencing of leg muscle tissues from 80D and 1D Muscovy ducks yielded comprehensive transcriptomic data, with 15,271 transcripts and 12,202 protein-coding genes successfully annotated. Comparative transcriptome analysis revealed 1,683 DEGs between developmental stages, comprising 865 upregulated and 818 downregulated genes in 80D samples (Supplement 1 and Figure 1). Notably, the top 30 most significantly upregulated genes (e.g., JCHAIN, METTL21C, and ALDH1A1) and downregulated genes (e.g., BD2, CHRM2, and DBX2) were systematically cataloged in Tables 1 and 2 respectively, providing crucial candidates for subsequent functional investigations into avian muscle development.
Functional characterization of differentially expressed genes in Muscovy duck myogenesis
Based on GO terms meeting statistical significance (p<0.05) and prior published evidence, we visualized GO enrichment for significantly up- and down-regulated genes in 80D muscle tissue. Upregulated DEGs showed predominant involvement in intracellular signal transduction, cell adhesion, chemotaxis, phagocytosis, apoptotic process regulation, and cytokine production regulation (Figure 2A and Supplement 2). In contrast, downregulated genes were significantly enriched in developmental processes including skeletogenesis, extracellular matrix remodeling, and neural stem cell maintenance, along with regulatory functions in ion homeostasis and vascular tone modulation, while demonstrating inhibitory effects on osteoblast proliferation and stimulatory roles in osteogenic differentiation (Figure 2B and Supplement 3).
Additionally, Figures 2C–2P reveals the expression outline for the DEGs between 80D and 1D in MyD88-dependent TLR, intracellular signal transduction, cell adhesion; the positive regulation of inflammatory response; apoptotic process regulation, phagocytosis, neuronal stem cell population maintenance, the positive regulation of osteoblast differentiation; collagen metabolic process, skeletal system development, ventricular septum morphogenesis, ion transmembrane transport regulation, the negative regulation of osteoblast proliferation; and blood vessel diameter regulation, respectively.
KEGG enrichment of the differentially expressed genes
Utilizing KEGG pathways demonstrating statistical significance (p<0.05) and supported by existing literature evidence, we conducted pathway enrichment visualization of DEGs (up- and down-regulated) in 80D muscle tissue samples. As shown in Figure 3A and Supplement 4, the upregulated DEGs in 80D leg muscle were involved in many signaling pathways, including MAPK, VEGF, mTOR, lysosome, FoxO, endocytosis, peroxisome, adipocytokine, nitrogen metabolism, metabolic pathways, cell adhesion molecules, and vascular smooth muscle contraction signaling pathways. Moreover, the downregulated genes in 80D muscle were involved in signaling pathways, such as MAPK, PPAR, Notch, Wnt, calcium, adipocytokine, metabolic pathways; amino acids biosynthesis; fatty acid degradation; and arginine biosynthesis signaling pathways (Figure 3B and Supplement 5). In addition, the upregulated DEGs in metabolic, Toll-like receptor, vascular smooth muscle contraction, mTOR, cell adhesion molecules, MAPK, focal adhesion signaling pathways were visualized in Figures 3C–3I using the heatmaps, respectively. Also, the downregulated DEGs in metabolic; focal adhesion; ECM-receptor interaction; glycine, serine and threonine metabolism; valine, leucine and isoleucine degradation; fatty acid metabolism; and fatty acid metabolism were showed in Figures 3J–3P, respectively.
PANTHER pathway analysis of the differentially expressed genes
As illustrated in Figure 4A, the upregulated in 80D muscle were related to diverse pathways, including VEGF, Jak/STAT, FGF, Toll receptor, PDGF, p53, endothelin, apoptosis, interleukin, inflammation, T cell activation, B cell activation, and 2-arachidonoylglycerol biosynthesis signaling pathways. Furthermore, the downregulated in 80D muscle were associated with multiple pathways, such as integrin, pyrimidine metabolism, Wnt, Heterotrimeric G-protein, and ionotropic glutamate receptor signaling pathways. (Figure 4B).
Protein classification for the differentially expressed genes
As illustrated in Figure 5A, the upregulated in 80D muscle were related to diverse proteins, such as hydrolase, phospholipase, integrin, lipase, dehydratase, phosphatase modulator, scaffold/adaptor protein, G-protein modulator, and metabolite interconversion enzyme. Moreover, the downregulated in 80D muscle were related to a massive number of proteins, including transporter, ion channel, growth factor, cytoskeletal protein, intercellular signal molecule, extracellular matrix structural protein, cell adhesion molecule, metabolite interconversion enzyme, extracellular matrix protein, histone modifying enzyme, basic leucine zipper transcription factor, and tyrosine protein kinase receptor (Figure 5B).
PPI network analysis
To elucidate key regulatory elements in Muscovy duck myogenesis, PPI networks were constructed for DEGs. Network topology analysis identified many critical myogenesis-related genes, with upregulated candidates in 80-day-old (80D) muscle tissue including immunoregulatory components (CD247, LCP2, LCK, CD3E), lysosomal protease (CTSS), and macrophage colony-stimulating factor receptor (CSF1R) (Figure 6A). Conversely, downregulated genes predominantly encoded extracellular matrix components (COL9A2, COL1A2) and metabolic enzymes (ALDH6A1, ACADL) (Figure 6B), suggesting distinct functional shifts during late-stage muscle development.
Identification and functional characterization of hub genes
Hub gene analysis was performed using CytoHubba with three topological algorithms (MCC, DMNC, and MNC). The consensus analysis revealed immune-related hub genes (CD3E, ITK) among upregulated candidates, while downregulated hubs comprised metabolic regulators including collagen type IV (COL4A6), mitochondrial enzymes (HADHB, MCCC2), and amino acid metabolism components (GNMT, PIPOX) (Figures 7A, 7B). Functional enrichment analyses demonstrated these hub genes’ involvement in leucine catabolism and fatty acid β-oxidation, along with participation in essential metabolic pathways: amino acid degradation, one-carbon metabolism, and folate cycle (Figure 7C). Notably, ITK was functionally annotated as a critical modulator of adaptive immunity, orchestrating T-cell and NKT-cell development and differentiation (Table 3), highlighting the intricate interplay between immune regulation and muscle development.
DISCUSSION
Muscle development of Muscovy ducks represents a complex and biologically significant phenomenon, influenced by multiple determinants including but not limited to genetic breed, ontogenetic stage, sexual dimorphism, and nutritional parameters [15–18]. Females develop significantly larger but less dense myofibers than males by postnatal day 42, a disparity that progressively intensifies until day 70 [16]. This phenotypic variation correlates with distinct endocrine profiles—males maintain higher serum testosterone levels associated with superior growth metrics, while females show elevated estradiol concomitant with upregulated lipid metabolism markers (CD36, CPT1A) and suppressed AMPK phosphorylation [17]. Transcriptomic analyses reveal 1,118 DEGs, including core myogenic regulators (MYLK4, KIT) and sex-specific modulators (TPM2, VCP) that orchestrate muscle hypertrophy through AMPK signaling, focal adhesion, and calcium-dependent pathways [16]. Notably, nutritional interventions such as 2.5% earthworm hydrolysate supplementation demonstrate the plasticity of this system, significantly enhancing muscle yield independent of sexual dimorphism [18]. Our investigation systematically resolves this knowledge gap through transcriptomic profiling, revealing 1,683 DEGs and computationally prioritizing ten hub genes (CD3E, ITK, COL4A6, HADHB, MCCC2, ACAA2, GNMT, PIPOX, SARDH, and IVD) via integrated RNA-seq and protein-protein interaction network analyses. The functional convergence of these hub genes on L-leucine degradation and fatty acid β-oxidation pathways [19] underscores metabolic reprogramming as an evolutionary adaptation for rapid muscle growth - a finding that corroborates emerging paradigms in avian energy metabolism.
Our study reveals an unexpected role of CD3E and ITK in post-hatching muscle development, challenging existing paradigms in muscle biology. Three lines of evidence demonstrate their unconventional functions: First, CD3E appears to activate duck myoblasts through conserved inflammatory pathways similar to those documented in mammalian muscle repair [20–22]. Second, ITK’s well-known lymphocyte differentiation function [12] seems evolutionarily repurposed to regulate myofiber maturation during crucial developmental stages. Most notably, their persistent high expression in 80-day post-hatching muscles indicates these molecules may have adapted to sustain muscle hypertrophy under continuous metabolic stress. These findings significantly expand our understanding of myogenic regulation beyond traditional pathways. These findings fundamentally expand the conceptual framework of muscle development by demonstrating that immune-related genes constitute essential components of the growth regulatory network in commercial poultry breeds [23–25].
In addition, we identified several downregulated hub genes (HADHB, ACAA2, GNMT, SARDH, IVD, MCCC2, PIPOX, and COL4A6) that are functionally linked to critical metabolic pathways, including glycine/serine/threonine metabolism, branched-chain amino acid (BCAA) degradation, and folate-mediated one-carbon metabolism (Figure 7D). Notably, HADHB and ACAA2—key regulators of mitochondrial fatty acid β-oxidation—showed reduced expression in 80D muscles, consistent with a metabolic shift from lipid breakdown to anabolic processes during hypertrophy. This parallels observations in broiler chickens, where suppressed fatty acid oxidation accompanies rapid muscle growth [26]. Similarly, GNMT and SARDH, which modulate folate-dependent one-carbon metabolism, may influence epigenetic regulation of myogenic factors like AKT and S6K1 to promote muscle development [27]. The decreased expression of IVD and MCCC2—enzymes involved in BCAA catabolism (Figures 3J, 3N)—further supports a preference for protein synthesis over amino acid degradation in mature muscle. Collectively, these metabolic adaptations align with the heightened anabolic demands of hypertrophic growth, facilitating sustained myofiber expansion [28].
The downregulation of PIPOX in 80D muscle (Figures 3J, 7D) was functionally linked to glycine, serine, and threonine metabolism. This aligns with established evidence that these pathways regulate skeletal muscle growth, fatty acid metabolism, and intramuscular fat deposition [29]. Concurrently, reduced COL4A6 (collagen IV) expression in focal adhesion and ECM-receptor interaction pathways (Figures 3K, 3N) appears to regulate myoblast differentiation, muscle fiber formation, and muscle size [16,30–32]. Focal adhesion dynamics correlate with distinct developmental stages during myogenesis—including differentiation, fiber formation, and hypertrophic responses to mechanical loading—playing critical roles in muscle cell homeostasis [30]. Supporting this, Zhang et al [16] identified focal adhesion and ECM-receptor interaction pathway genes as determinants of breast muscle hypertrophy in Muscovy ducks, while Wu et al demonstrated their conserved role in driving skeletal muscle development across avian species (ducks and quails) [31,32]. These findings collectively highlight coordinated metabolic and structural adaptations underlying muscle development.
CONCLUSION
While this study provides novel insights, several limitations warrant attention. First, the functional roles of prioritized hub genes (e.g., CD3E, ITK) in myogenesis remain hypothetical and require experimental validation through knockout or overexpression models. Second, the interplay between metabolic pathways needs mechanistic exploration to clarify their synergistic effects on myogenesis. Third, the study focused on transcriptomic changes, overlooking post-translational modifications or epigenetic regulation that may further modulate myogenesis. Future work should integrate multi-omics approaches (proteomics, metabolomics) and in vivo interventions to validate these targets and refine strategies for enhancing poultry meat quality.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Khalil KM El-Shater S Rizk H Khalifa E Embryonic thermal manipulation of poultry birds: lucrative and deleterious effects J Anim Physiol Anim Nutr 2024 108 346 56 10.1111/jpn.13896 37885333 · doi ↗ · pubmed ↗
- 2Zhuang X Xie F Lin Z Effect of mi R-493-5p on proliferation and differentiation of myoblast by targeting ANKRD 17 Cell Tissue Res 2023 393 119 32 10.1007/s 00441-023-03777-3 37178193 · doi ↗ · pubmed ↗
- 3Lin R Li H Lai L Analysis of genetic structure and identification of important genes associated with muscle growth in Fujian Muscovy duck Poult Sci 2024 103 104445 10.1016/j.psj.2024.104445 39504826 PMC 11570716 · doi ↗ · pubmed ↗
- 4Plotkin DL Delcastillo K Van Every DW Tipton KD Aragon AA Schoenfeld BJ Isolated leucine and branched-chain amino acid supplementation for enhancing muscular strength and hypertrophy: a narrative review Int J Sport Nutr Exerc Metab 2021 31 292 301 10.1123/ijsnem.2020-0356 33741748 · doi ↗ · pubmed ↗
- 5Shamim B Hawley JA Camera DM Protein availability and satellite cell dynamics in skeletal muscle Sports Med 2018 48 1329 43 29557519 10.1007/s 40279-018-0883-7 · doi ↗ · pubmed ↗
- 6Chen MM Zhao Y Yu K A MSTN Del 73C mutation with FGF 5 knockout sheep by CRISPR/Cas 9 promotes skeletal muscle myofiber hyperplasia e Life 2024 12 RP 86827 10.7554/e Life.86827.3 39365728 PMC 11452178 · doi ↗ · pubmed ↗
- 7Schlune A Riederer A Mayatepek E Ensenauer R Aspects of newborn screening in isovaleric acidemia Int J Neonatal Screen 2018 4 7 10.3390/ijns 4010007 33072933 PMC 7548899 · doi ↗ · pubmed ↗
- 8Järvinen E Ismail K Muniandy M Biotin-dependent functions in adiposity: a study of monozygotic twin pairs Int J Obes 2016 40 788 95 10.1038/ijo.2015.237 26601567 · doi ↗ · pubmed ↗