Elucidating Circular Ribonucleic Acid Mechanisms Associated with Splicing Factor 3 Inhibition in Cervical Cancer
Amahle Nyalambisa, Babatunde Adebola Alabi, Zodwa Dlamini, Rahaba Marima

TL;DR
This study investigates how circular RNAs and a splicing factor inhibitor affect cervical cancer cells, suggesting theophylline could be a potential treatment.
Contribution
The study identifies novel circular RNA-miRNA interactions and demonstrates the cytotoxic effects of theophylline in cervical cancer cell lines.
Findings
Theophylline induces cell cycle arrest and apoptosis in cervical cancer cell lines.
hsa_circ_0001038 interacts with miR-205-5p, a tumor-suppressive miRNA, while circRNA_400029 interacts with miR-16-5p.
A ceRNA network and KEGG pathway analysis reveal key signaling pathways involved in cervical cancer progression.
Abstract
Cervical cancer (CCa) is the fourth leading cause of cancer-related deaths among women worldwide, with nearly 90% of cases in low- and middle-income countries, especially in Sub-Saharan Africa. This study explores the roles of circular ribonucleic acids (circRNAs), hsa_circ_0001038 and circRNA_400029, and the impact of the serine/arginine-rich splicing factor 3 (SRSF3) inhibitor, theophylline, in CCa cell lines. We utilized cell cycle fluorescence-activated cell sorting (FACS) and Annexin V/propidium iodide (PI) assays to evaluate theophylline’s effects on SiHa and C33A cell lines. Results showed S-phase arrest in SiHa and G2/M arrest in C33A, with significant cytotoxic effects indicated by apoptosis analysis. Using CircAtlas, we identified micro ribonucleic acids (miRNAs) binding to hsa_circ_0001038, particularly miR-205-5p, which has a tumour-suppressive role. miRTarBase identified…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
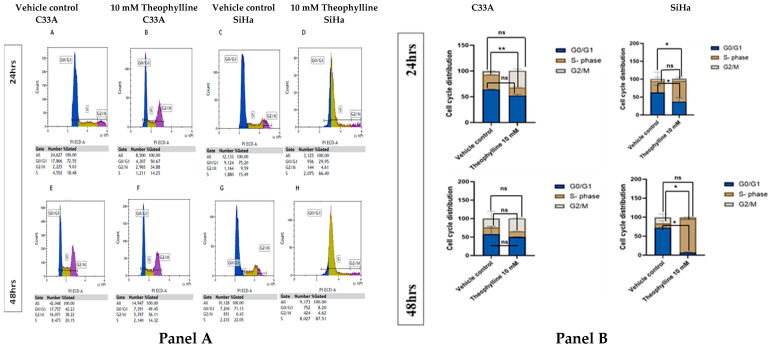 Figure 1
Figure 1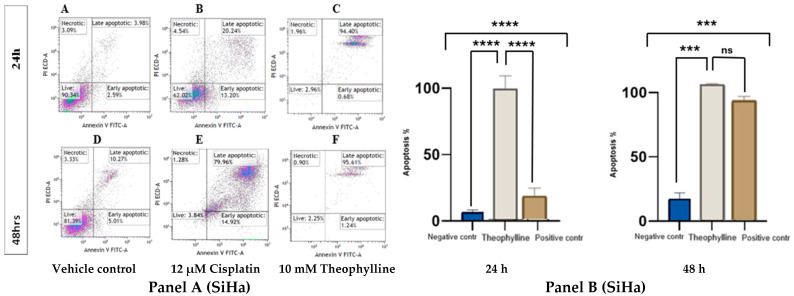 Figure 2
Figure 2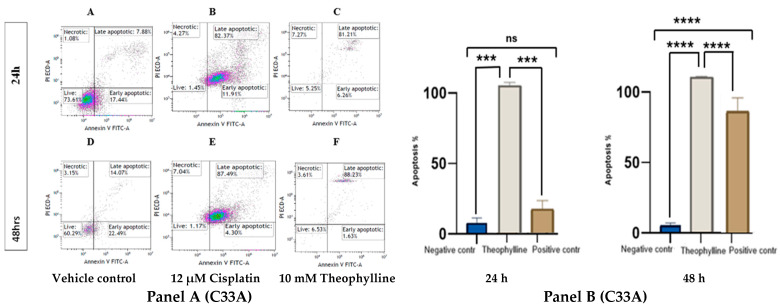 Figure 3
Figure 3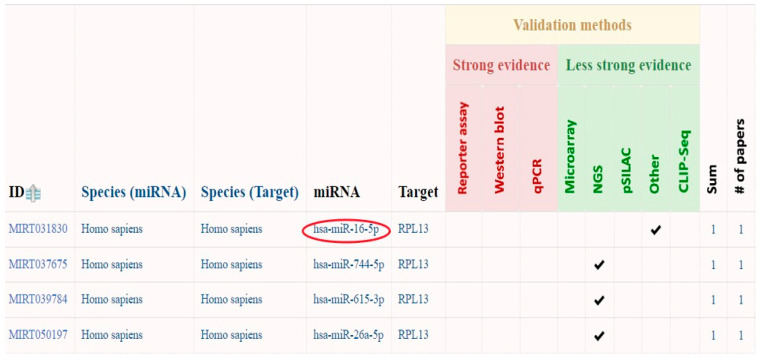 Figure 4
Figure 4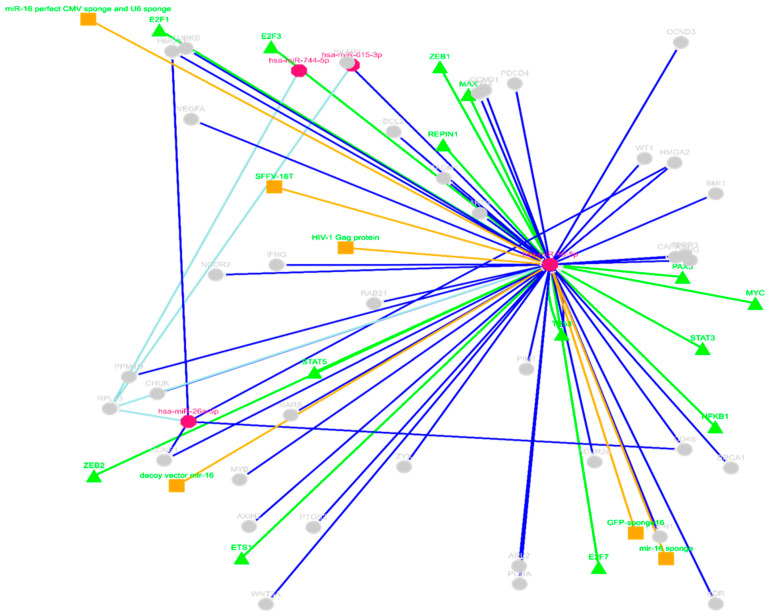 Figure 5
Figure 5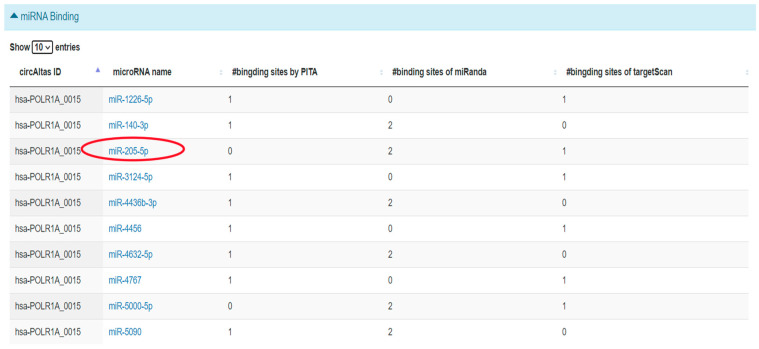 Figure 6
Figure 6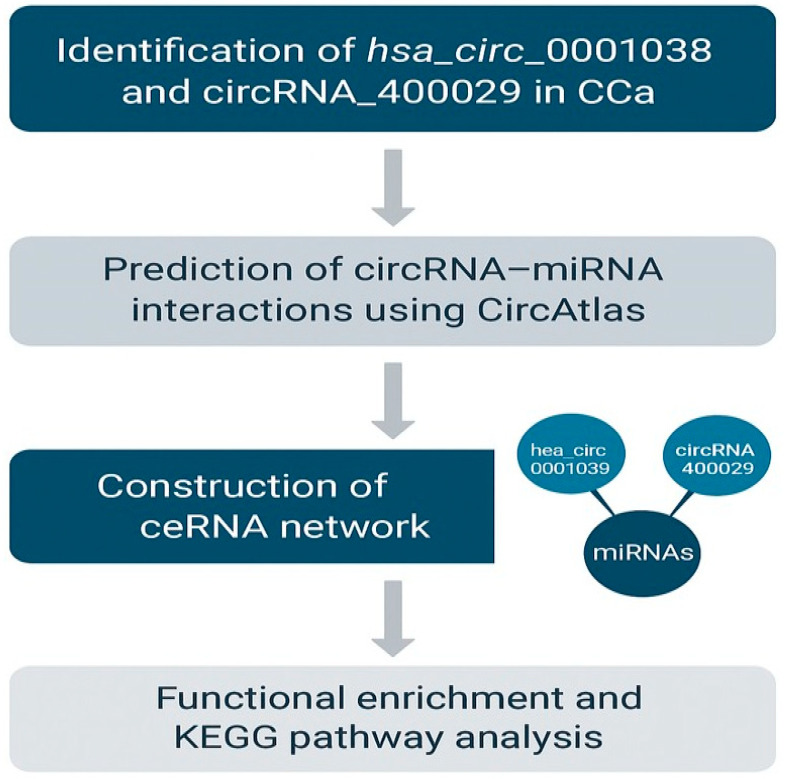 Figure 7
Figure 7- —South African Medical Research Council (SAMRC)
- —National Research Foundation (NRF)
- —University of Pretoria
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircular RNAs in diseases · RNA Research and Splicing · RNA modifications and cancer
1. Introduction
Cervical cancer (CCa) is the fourth most prevalent cause of cancer-related deaths among women worldwide [1]. Nearly ninety percent of CCa cases occur in low- and middle-income countries (LMICs), with Sub-Saharan Africa (SSA) accounting for over eighty percent of the global burden [2]. Numerous environmental and genetic risk factors are associated with CCa, including smoking, alcohol use, multiple sexual partners, unprotected intercourse, low socioeconomic status, a family history of CCa, prolonged oral contraceptive use, human immunodeficiency virus (HIV) infection, and persistent human papillomavirus (HPV) infection [3]. Persistent HPV infection, in particular, is the primary risk factor for CCa, with HPV 16 responsible for approximately 55–60% of cases, followed closely by HPV 18 [4]. Conventional treatments for CCa include surgery, radiation therapy, chemoradiation, external beam radiation therapy (EBRT), chemotherapy, and brachytherapy. However, these methods often fail to achieve the desired outcomes and can result in significant adverse effects [5,6,7]. Therefore, identifying novel diagnostic biomarkers and promising therapeutic targets is crucial for the effective detection and eradication of tumour cells in cervical tissues.
Noncoding ribonucleic acids (ncRNAs) have been shown to play key roles in cancer progression and may serve as potential therapeutic targets to improve CCa prognosis [8,9,10]. In mammalian cells, noncoding transcripts constitute approximately 99% of total ribonucleic acid (RNA) [11,12]. Circular ribonucleic acids (circRNAs), a subtype of ncRNAs, are characterized by their covalently closed structure, which renders them resistant to exonuclease degradation [13,14]. CircRNAs are typically located in the cytoplasm of eukaryotic cells [15]. Unlike canonical splicing of linear RNA, circRNAs are produced through back-splicing, which covalently joins a downstream 3′ splice donor site to an upstream 5′ splice acceptor site, forming a closed loop [16,17]. The serine/arginine-rich splicing factor 3 (SRSF3) has been shown to play a role in this process in other cancers [18].
SRSF3 is a member of the serine/arginine-rich (SR) protein family—an important group of proteins that play a pivotal role in CCa development by influencing RNA splicing [19]. Within this family, SRSF3, the smallest member, is highly expressed in CCa and significantly contributes to tumourigenesis. However, the regulatory mechanisms involving SRSF3 and circRNAs in CCa remain poorly understood. CircRNAs have been implicated in various cancers as either oncogenic or tumour-suppressive molecules, making them potential targets for novel therapeutic strategies. For instance, circRNA_400029 is upregulated in cervical cancer and promotes cell proliferation, migration, and invasion while inhibiting apoptosis via the miR-1285-3p/TLN1 axis [20]. Similarly, hsa_circ_0001038 is overexpressed in cervical cancer and promotes proliferation, migration, and invasion by acting as a ceRNA for miR-337-3p [21]. Moreover, circRNA_101996 has been reported to accelerate cervical cancer development through regulation of the miR-1236-3p/TRIM37 axis, and circ_0084927 facilitates tumour progression by sponging miR-142-3p and upregulating ARL2 [22]. Conversely, circ_0067934 silencing induces apoptosis and cell-cycle arrest, while circ_0000515 downregulation suppresses proliferation through the miR-326/SOX11 axis [23].
Theophylline, an SRSF3 inhibitor, has been identified as a potential therapeutic agent for investigating the effects of SRSF3 suppression in CCa cells [24]. While SRSF3 has been associated with the regulation of key oncogenic pathways in CCa, its interaction with circRNAs remains largely unexplored. SRSF3 inhibition could potentially alter circRNA expression patterns, leading to changes in cell-cycle regulation, apoptosis, and cancer cell proliferation. By evaluating circRNA expression profiles and their roles in cellular pathways following SRSF3 inhibition, this study aims to provide new insights into the therapeutic potential of targeting splicing factors and circular RNAs in CCa. Understanding these molecular interactions could enhance treatment strategies and potentially support the repurposing of theophylline for CCa therapy.
2. Results
2.1. Theophylline Alters Cell Cycle Progression in CCa Cells
Theophylline treatment induces significant alterations in cell cycle distribution in both SiHa and C33A CCa cell lines (Figure 1A). In C33A, at 24 h, the G0/G1 population significantly decreased from 72.55% (Figure 1A) to 50.68% (Figure 1B) (p = 0.0053), while the G2/M population significantly increased from 9.03% (Figure 1A) to 34.88% (Figure 1B) (p = 0.0018), suggesting a G2/M phase arrest. At 48 h, the G0/G1 population modestly increased from 42.23% (Figure 1E) to 49.45% (Figure 1F), and the S-phase decreased from 20.15% (Figure 1E) to 14.32% (Figure 1F); however, these changes were not statistically significant (p = 0.147 for G0/G1; p = 0.231 for S). In the SiHa cell line at 24 h, a significant reduction in the G0/G1 phase was observed, from 75.20% (Figure 1C) to 29.95% (Figure 1D) (p < 0.0001), alongside a significant increase in the S-phase population from 15.49% (Figure 1C) to 66.40% (Figure 1C) (p < 0.0001), indicating strong S-phase arrest. By 48 h, this effect intensified, with G0/G1 phase dropping from 71.13% (Figure 1G) to 8.20% (Figure 1H) (p < 0.0001) and S-phase cells increasing from 22.05% (Figure 1G) to 87.51% (Figure 1H) (p < 0.0001).
2.2. Theophylline Induces Apoptosis in CCa Cell Line SiHa and C33A
Theophylline treatment significantly induced apoptosis in both SiHa (Figure 2, Panel A) and C33A (Figure 3, Panel A) cervical cancer cell lines. In both models, there was a marked increase in apoptotic populations compared to the control treatments. At 24 h, theophylline exhibited a notable apoptotic effect that was comparable to cisplatin (12 μM) in C33A cells but slightly less pronounced in SiHa cells. By 48 h, the apoptotic response to theophylline was markedly enhanced in both cell lines, demonstrating a clear time-dependent effect. In SiHa, theophylline treatment significantly increased late apoptotic populations compared to controls at both time points. Although the apoptotic response in SiHa cells was slightly lower than that induced by cisplatin, it remained statistically significant (p < 0.0001 vs. vehicle control; Figure 2, Panel B) and consistent over time. In C33A, theophylline exerted a significant increase in late apoptotic cells at both 24 and 48 h, exceeding levels observed in the vehicle control and surpassing even those induced by cisplatin (p < 0.0001 vs. vehicle control; p < 0.01 vs. cisplatin; Figure 3, Panel B).
Flow cytometric profiles further revealed a distinct rise in Annexin V^+^/PI^−^ cells at 24 h, indicating that theophylline initially triggers early apoptosis before progressing to the late apoptotic phase at 48 h. The increase in Annexin V^+^/PI^+^ populations at later times likely reflects secondary necrosis accompanying advanced apoptotic stages, rather than primary necrosis. These findings collectively suggest that theophylline induces a genuine apoptotic cascade that evolves, leading to sustained cytotoxic effects in both CCa cell lines.
2.3. In Silico Bioinformatics Analysis of Differentially Expressed circRNAs hsa_circ_0001038 & circRNA_400029 in CCa
2.3.1. Prediction of circRNA_400029 miRNA Interactions Using miRTarBase
MiRTarBase, a database of validated miRNA-target interactions, identified potential miRNAs targeting circRNA_400029. The analysis revealed that circRNA_400029 originates from the ribosomal protein L13 (RPL13) gene and interacts with four specific miRNAs (Figure 4). Among these, miR-16-5p was selected for further investigation. As a member of the miR-16 family, miR-16-5p plays a role in multiple molecular pathways linked to CCa progression. Notably, it functions as a tumour suppressor in CCa, with the potential to inhibit tumour growth and angiogenesis.
2.3.2. ceRNAs Network of circRNA_400029—miRNA Using miRTarBase
The miRTarBase was used to identify ceRNAs for circRNA_400029. The analysis identified several hundred predicted downstream gene targets of miR-16-5p. It was found that a single gene can be targeted by multiple miRNAs within the network, and vice versa (Figure 5). For instance, cyclin-dependent kinase 6 (CDK6), G1/S-specific cyclin-D1 (CCND1), High Mobility Group AT-Hook 1 (HMGA1), and High Mobility Group AT-Hook 2 (HMGA2) interact with both hsa-miR-16-5p and hsa-miR-26a-5p. On the other hand, miR-744-5p and miR-615-3p did not share any common mRNA targets with other miRNAs in the ceRNA network. The mRNA targets available in the database depend on experimentally validated data uploaded by the research community, so the absence of shared targets for miR-744-5p and miR-615-3p may be due to limited research on these miRNAs rather than a lack of actual interactions.
2.3.3. MiR-16-5p—Gene Target Pathway Analysis Using KEGG Pathway
The miR-16-5p gene target pathway analysis showed that miR-16-5p targets WNT4, a key ligand in the Wnt/β-catenin signalling pathway. KEGG pathway analysis indicated that downstream components of the Wnt/β-catenin pathway, including FZD, LRP5/6, DVL, and FRAT, are potentially upregulated in association with increased WNT4 expression. Additionally, SRSF3 was identified as a downstream target of β-catenin/TCF4 transcriptional activity. This suggests that circRNA_400029 may promote CCa progression by targeting the miR-16-5p/Wnt/β-catenin/SRSF3 axis.
2.3.4. Prediction of hsa_circ_0001038—miRNA Interactions Using circAtlas
CircAtlas predicted a total of 23 miRNAs (Figure 6) to have a binding spot for hsa_circ_0001038. MiR-205-5p was then chosen for further analysis. MiR-205-5p is a tumour-suppressive miRNA that belongs to the miR-200 family, which is known for its role in regulating epithelial to mesenchymal transition (EMT) and influencing CCa progression [25,26].
2.3.5. ceRNA Network of miR-205-5p Using miRTarBase
MiRTarBase was then used to explore the potential downstream targets of miR-205-5p. The database returned several hundred predicted gene targets. To narrow this down, only the gene targets linked to CCa pathogenesis were selected for inclusion in the ceRNA network (Figure 7).
Among the identified gene targets, DEAD-box helicase 5 (DDX5) emerged as a key gene of interest due to its known involvement in cancer-related pathways. DDX5 was predicted to be a downstream effector regulated by the circRNA hsa_circ_0001038 via the miR-205-5p/SRSF3/DDX5 axis. Interaction analyses suggested that hsa_circ_0001038 may act as a molecular sponge for miR-205-5p, a tumour-suppressive miRNA, thereby potentially modulating DDX5 expression. Additionally, SRSF3 was identified as a regulatory splicing factor influencing DDX5 isoform expression. Notably, samples with elevated SRSF3 expression also exhibited increased levels of the DDX5-L isoform, consistent with previous findings that link this variant to oncogenic signalling. These findings support a putative regulatory relationship involving hsa_circ_0001038, miR-205-5p, SRSF3, and DDX5 in CCa.
3. Discussion
CCa remains one of the most prevalent causes of cancer-related mortality in women, particularly in LMICs, where it accounts for a disproportionate global burden [2]. Sub-Saharan Africa alone contributes over 80% of global CCa cases [3], underscoring the need for innovative diagnostic and therapeutic approaches that are both effective and accessible. In recent years, circRNAs have emerged as important regulatory molecules in various cancers, including CCa [27], particularly hsa_circ_0001038 [28] and circRNA_400029 [29]. CircRNAs are generated through back-splicing mechanisms, with SRSF3 playing a pivotal role in their biogenesis in other cancers [23]. However, the functional implications and downstream regulatory networks involving these circRNAs in CCa remain poorly understood. This study investigated the effects of splicing factor 3 inhibition using theophylline, a methylxanthine derivative known to modulate pre-mRNA splicing, on two cervical cancer (CCa) cell lines—HPV-16-positive SiHa and HPV-negative C33A. Cell cycle and apoptosis analyses revealed that theophylline exerts both cytostatic and cytotoxic effects in a cell line–dependent manner. Specifically, theophylline induced S-phase arrest in SiHa and G2/M arrest in C33A, indicating disruption of key cell cycle checkpoints and replication control. These alterations were accompanied by a significant increase in apoptotic cell populations, with effects comparable to or exceeding those of cisplatin, a standard chemotherapeutic agent. Importantly, although a marked elevation in late apoptotic (Annexin V^+^/PI^+^) populations was observed, this may partly reflect secondary necrotic events that occur following the progression of apoptosis, especially given the relatively high concentration (10 mM) of theophylline used. Therefore, the primary theophylline-induced effect is more accurately attributed to the activation of early apoptotic signalling rather than direct necrosis. The significant rise in Annexin V^+^/PI^−^ cells at 24 h further supports this interpretation, indicating that apoptosis is initially triggered before membrane disruption occurs. Previous studies have demonstrated that inhibition of splicing factors can lead to cell cycle arrest and apoptosis across various tumour models. For example, Zhang et al. [30] reported that inhibition of SF3B1 induced G2/M phase arrest and apoptotic signalling in gastric cancer cells, consistent with our findings in C33A. Similarly, Pérez-Pérez et al. [31] showed that methylxanthines such as theophylline disrupt cell cycle progression and trigger apoptosis in glioblastoma cells, supporting our observation of S-phase arrest in SiHa. Collectively, our findings highlight that theophylline induces early apoptotic responses and cell cycle dysregulation through potential interference with splicing-dependent regulators, underscoring its promise as a repurposed therapeutic candidate in CCa. Future dose–response studies using lower concentrations will be necessary to delineate the threshold between apoptotic and necrotic outcomes and to optimize its therapeutic window.
Moreover, the relationship between cytotoxicity and RNA integrity has been explored in the context of splicing factor inhibition. Previous reports by Petasny et al. [32] and Bradley et al. [33] have revealed that splicing factor disruption not only affects cell cycle progression but also impacts RNA stability, complicating downstream analyses. Our findings align with these observations, suggesting that the cytotoxic effects of theophylline may extend beyond immediate cellular responses to include detrimental impacts on RNA quality. In light of these challenges, future studies should consider alternative methodologies, such as single-cell RNA sequencing or RNA-seq, which may provide insights into gene expression changes without the constraints imposed by RNA integrity issues. Balancing therapeutic potency with cell viability to maintain molecular integrity is a common challenge in translational cancer research. However, the observed cell death underscores the therapeutic potential of theophylline in cancer.
To further explore the molecular mechanisms potentially regulated by theophylline, bioinformatics analyses revealed that MiR-16-5p targets several oncogenic mRNAs such as CDK6, CCND1, and HMGA2, which are implicated in unchecked cell proliferation and chromatin remodelling. Notably, many of these targets are components of the Wnt/β-catenin signalling pathway, a pathway commonly activated by HPV oncogenes during cervical tumorigenesis. By sponging miR-16-5p, circRNA_400029 may contribute to oncogenic signalling cascades, promoting tumour survival and progression. Similarly, hsa_circ_0001038 was predicted to sponge miR-205-5p, another tumour-suppressive miRNA involved in the inhibition of epithelial-to-mesenchymal transition (EMT) and metastasis. The downregulation of miR-205-5p has been linked to increased expression of DDX5 [34], a cofactor of SRSF3 that promotes oncogenic alternative splicing and activates the PI3K/AKT signalling pathway. This regulatory axis supports CCa cell proliferation and resistance to apoptosis. The mutual reinforcement between SRSF3, DDX5, and their downstream targets suggests a feedforward loop, potentially exacerbated by circRNA-mediated miRNA sequestration. These ceRNA networks may be central to cervical tumorigenesis and could represent novel targets for therapeutic intervention. Additionally, pathway enrichment analysis using KEGG highlighted several signalling axes associated with CCa, including the HPV E5–EGFR–RAS–ERK and HPV E6–Notch pathways, which are known to promote oncogenesis through enhanced cell proliferation, survival, and splicing deregulation. The overlap between these signalling pathways and those regulated by miR-16-5p and miR-205-5p further supports the functional relevance of circRNA_400029 and hsa_circ_0001038 in CCa biology.
4. Materials and Methods
4.1. Cell Culture and Drug Treatment
CCa cell lines SiHa (HPV 16+) and C33A (HPV^−^) were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). SRSF3 inhibitor (Theophylline) was purchased from MedChemExpress (Monmouth Junction, NJ, USA). Cisplatin was purchased from Sigma-Aldrich^®^ Solutions (St. Louis, MO, USA). The human CCa cell lines SiHa (HPV 16+) and C33A (HPV^−^) were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM: F12) (Thermo Fisher Scientific, Waltham, MA, USA). This medium was enriched with 10% (v/v) fetal bovine serum (FBS) from Gibco (Thermo Fisher Scientific, Waltham, MA, USA) and supplemented with 1% streptomycin (Thermo Fisher Scientific, Waltham, MA, USA). The cells were maintained at 37 °C in a humidified incubator with 5% CO_2_ until they reached 70% confluency. Before theophylline treatment, the cells were subjected to serum starvation for 24 h to synchronize the cell cycle.
4.2. Propidium Iodide (PI) Cell Cycle FACS Analysis
The C33A and SiHa cells were treated with 10 mM theophylline for 24 and 48 h, with 0.1% DMSO as a control. Cells were cultured at a density of 5 × 10^5^ cells per well in 6-well plates and harvested by centrifugation at 500× g for 5 min. The supernatant was discarded, and cells were washed twice with ice-cold 1× PBS. The cells were fixed by adding 7 mL of ice-cold 70% ethanol while vortexing and stored at −20 °C for 24 h. After fixation, cells were centrifuged again, washed with PBS, and resuspended. Cells were stained with a solution containing propidium iodide (PI) and RNase A, achieving final concentrations of 1 µg/mL of PI and 0.2 mg/mL of RNase A. The staining solution was incubated for 30 min at 37 °C, then placed on ice. The samples were analyzed by flow cytometry (Beckman Coulter CytoFLEX cytometer) (Beckman Coulter, Brea, CA, USA) using Kaluza analysis software version 2.1 (Beckman Coulter, Brea, CA, USA)within an hour, and the results were presented as histograms.
4.3. Apoptosis Assay by FACS
C33A and SiHa cells were cultured separately and treated with 10 mM theophylline for 24 and 48 h [24]. Cisplatin (12 μM) served as a positive control, while DMSO was used as a negative control. After treatment, cells were digested with EDTA-free trypsin and centrifuged at 300× g for 5 min at 4 °C. The cells were then incubated under optimal culture conditions for 30 min. Cells were washed twice with ice-cold PBS, centrifuged again at 300× g for 5 min at 4 °C, and resuspended in 100 μL of 1× binding buffer at a density of 1–5 × 10^5^ cells. To each tube, 4–5 μL of CL488-Annexin V and 5 μL of PI working solution were added. Compensation adjustments were made using additional tubes with single dyes (either CL488-Annexin V or PI). Samples were incubated for 10–15 min at room temperature in the dark. After incubation, 400 μL of 1× binding buffer (or 1× PBS) was added to each tube. The samples were analyzed using the Beckman Coulter CytoFlex flow cytometer within an hour, and the results were presented as histograms.
4.4. Bioinformatics Analysis
To determine the interactions of the hsa_circ_0001038 and circRNA_400029, bioinformatics tools including CircAtlas, miRTarBase, and KEGG pathway were used. CircAtlas was used to predict miRNA binding sites of hsa_circ_0001038. However, circRNA_400029 could not be found in CircAtlas. This could be because this database relies on experimental data uploaded by the research community, and this circRNA may not have been included in the available datasets. Therefore, to explore miRNA interactions for circRNA_400029, MiRTarBase was used. MiRTarBase was also used to visualize the ceRNA network of the two circRNAs associated with miRNAs and mRNAs, which were identified using circAtlas for hsa_circ_0001038 and miRTarBase for circRNA_400029. KEGG pathway was used to map the miRNA-gene target pathway.
4.5. Statistical Analysis
Data acquired from at least 3 independent experiments, each in at least triplicate, was analyzed using GraphPad Prism version 9.0 statistical software. Statistical significance was performed using ONE-WAY ANOVA followed by Tukey’s post hoc test for multiple comparisons. The significance level was deemed at p < 0.05. The results are presented as mean ± standard error of the mean (SEM).
5. Conclusions and Future Perspective
This study demonstrated that SRSF3 inhibition using theophylline exerts potent cytostatic and pro-apoptotic effects in CCa cells by interfering with cell cycle progression and inducing apoptosis. Bioinformatics analysis further implicates circRNA_400029 and hsa_circ_0001038 as key regulators in CCa through their interaction with tumour-suppressive miRNAs and downstream ceRNA oncogenic pathways. Future research should include dose–response studies with lower theophylline concentrations to confirm its selective apoptotic effects and minimize necrosis. Functional validation of circRNA_400029–miR-16-5p and hsa_circ_0001038–miR-205-5p interactions through knockdown or overexpression experiments will help clarify their regulatory roles. Integrating RNA sequencing and proteomic analyses could further elucidate how theophylline modulates SRSF3-dependent pathways and downstream signalling networks. Finally, in vivo and organoid-based studies are essential to evaluate the therapeutic relevance of theophylline and the biomarker potential of these circRNAs in cervical cancer diagnosis and treatment response.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Castle P.E. Einstein M.H. Sahasrabuddhe V.V. Cervical cancer prevention and control in women living with human immunodeficiency virus CA Cancer J. Clin.20217150552610.3322/caac.2169634499351 PMC 10054840 · doi ↗ · pubmed ↗
- 2Zhang J. Qiu H. Qing Y. Hu Y. Deng H. A commentary on ‘Local excision as a viable alternative to hysterectomy for early-stage cervical cancer in women of reproductive age: A population-based cohort study’Int. J. Surg.20241103120312110.1097/JS 9.000000000000121838377050 PMC 11093455 · doi ↗ · pubmed ↗
- 3Zhang S. Xu H. Zhang L. Qiao Y. Cervical cancer: Epidemiology, risk factors and screening Chin. J. Cancer Res.20203272010.21147/j.issn.1000-9604.2020.06.0533446995 PMC 7797226 · doi ↗ · pubmed ↗
- 4Yousaf S. Shehzadi A. Ahmad M. Asrar A. Ahmed I. Iqbal H.M. Bule M.H. Recent advances in HPV biotechnology: Understanding host-virus interactions and cancer progression—A review Int. J. Surg.20241108025803610.1097/JS 9.000000000000211739806745 PMC 11634192 · doi ↗ · pubmed ↗
- 5Kumar L. Harish P. Malik P.S. Khurana S. Chemotherapy and targeted therapy in the management of cervical cancer Curr. Probl. Cancer 20184212012810.1016/j.currproblcancer.2018.01.01629530393 · doi ↗ · pubmed ↗
- 6Chargari C. Peignaux K. Escande A. Renard S. Lafond C. Petit A. Kee D.L.C. Durdux C. Haie-Méder C. Radiotherapy of cervical cancer Cancer Radiothérapie 20222629830810.1016/j.canrad.2021.11.00934955418 · doi ↗ · pubmed ↗
- 7Guo L. Hua K. Cervical cancer: Emerging immune landscape and treatment Onco Targets Ther.2020138037804710.2147/OTT.S 264312 PMC 743451832884290 · doi ↗ · pubmed ↗
- 8Movahedpour A. Khatami S.H. Khorsand M. Salehi M. Savardashtaki A. Mirmajidi S.H. Negahdari B. Khanjani N. Naeli P. Vakili O. Exosomal noncoding RN As: Key players in glioblastoma drug resistance Mol. Cell. Biochem.20214764081409210.1007/s 11010-021-04221-234273059 · doi ↗ · pubmed ↗