Evolutionary Conservation and Regulatory Diversification of AS1 Homologs in Soybean
Dan Wang, Xuan Zhou, Dongfa Wang, Xiangtao Yang, Yexin He, Zhengjun Xia, Jianghua Chen, Weiyue Zhao

TL;DR
This study explores the evolutionary conservation and regulatory diversity of AS1 genes in soybean, revealing their role in leaf development and stress adaptation.
Contribution
The study identifies and characterizes four AS1 paralogs in soybean, showing functional conservation and regulatory diversification.
Findings
Four AS1 paralogs in soybean show functional conservation in leaf polarity regulation.
GmAS1 genes exhibit distinct tissue-specific expression patterns and promoter cis-elements.
Promoter analysis suggests subfunctionalization related to light, hormones, development, and stress.
Abstract
The establishment of dorsoventral polarity is a critical step in leaf morphogenesis, enabling the transition from radial primordia to flattened laminae. The MYB domain transcription factor ASYMMETRIC LEAVES1 (AS1) plays a central role in this process by regulating leaf polarity and developmental transitions, primarily through the repression of Class I KNOX genes. Here, four AS1 paralogs were identified in soybean (Glycine max), two of which showed collinearity with Arabidopsis thaliana and Medicago truncatula. The AS1 proteins of soybean and Arabidopsis exhibit high conservation, whereas the four GmAS1 genes in soybean display different tissue-specific expression patterns. Strikingly, each GmAS1 gene was able to fully rescue the defective phenotype of the Arabidopsis as1 mutant, indicating that GmAS1 genes are functionally conserved in leaf polarity regulation. Promoter analysis further…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
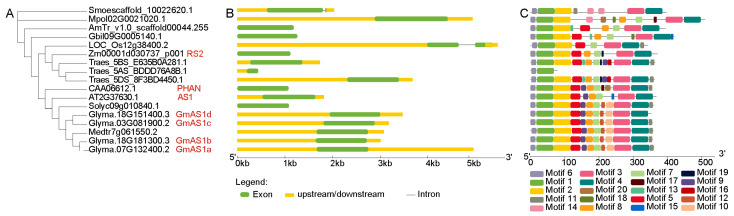 Figure 1
Figure 1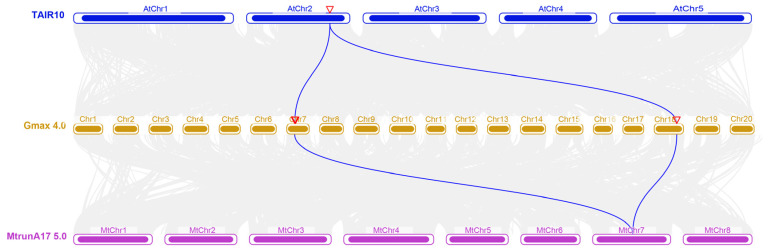 Figure 2
Figure 2 Figure 3
Figure 3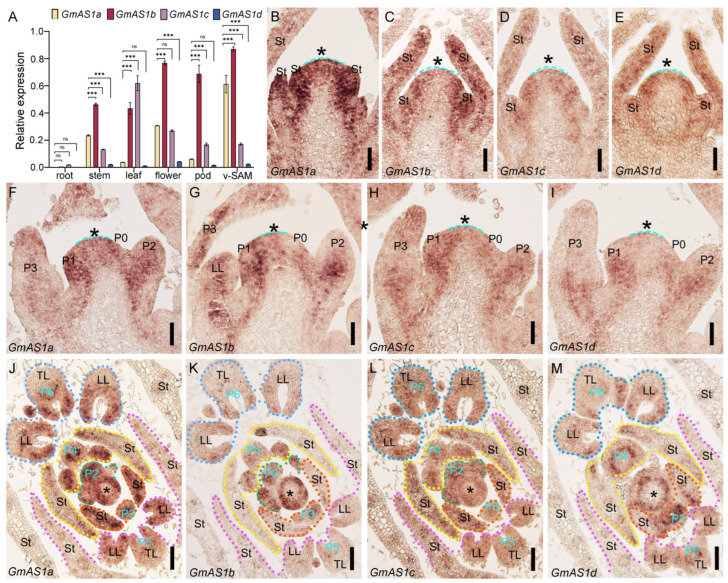 Figure 4
Figure 4 Figure 5
Figure 5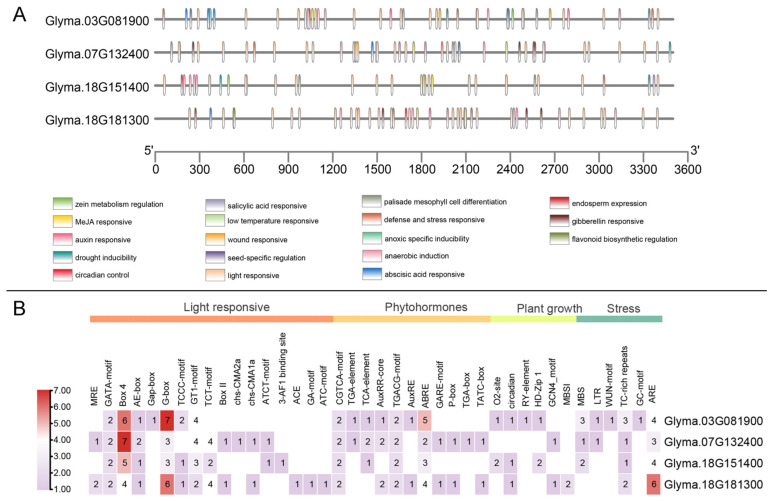 Figure 6
Figure 6- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Polysaccharides and Plant Cell Walls · Plant Reproductive Biology
1. Introduction
Leaf morphology plays an important role in plant growth and development by determining photosynthetic efficiency, transpiration dynamics, and resource allocation. Morphological variation influences biomass accumulation, stress adaptation, and source-sink relationships essential for reproductive success. Therefore, leaf morphology represents a key integrator of physiological performance and developmental plasticity. The establishment of leaf morphology primarily involves development along three axes: dorsoventral (adaxial–abaxial), proximal–distal (base–tip), and mediolateral (center–margin) [1]. The establishment of adaxial–abaxial polarity is particularly critical, as it transforms the leaf primordium from a radial, stem-like structure into a flattened, bilateral organ essential for physiological functions like photosynthesis and transpiration [2]. This process is controlled by a complex regulatory network of transcription factors and signaling pathways that coordinate tissue differentiation and organ morphogenesis [3].
A key component of this network is the ARP gene, named after ASYMMETRIC LEAVES1 (AS1) from A. thaliana, ROUGH SHEATH2 (RS2) from Zea mays, and PHANTASTICA (PHAN) from Antirrhinum majus, which belongs to a highly conserved family of MYB transcription factors [4,5,6]. Loss-of-function mutations in these genes result in severe developmental abnormalities. For example, in snapdragon, phan mutants produce radially symmetrical leaves that lack adaxial identity, a phenotype linked to a failure to establish proper adaxial–abaxial polarity [7]. Similarly, in maize, rs2 mutants exhibit disorganized leaf blade–sheath boundaries and other patterning defects [5]. In A. thaliana, as1 mutant leads to asymmetric, curled, or wrinkled leaves [4]. Despite these phenotypic differences, a conserved regulatory mechanism has been established: ARP proteins, in conjunction with the LATERAL ORGAN BOUNDARIES (LOB) domain protein AS2, form a repressor complex that directly binds to the promoters of Class I KNOTTED1-like homeobox (KNOX1) genes to repress their expression [4,5,6,8,9,10,11,12,13,14]. This repression prevents the retention of meristem identity in the leaf, thereby ensuring the transition from indeterminate stem cell fate to determinate leaf fate [4]. In addition to governing dorsoventral polarity in simple leaves, ARP gene also precisely regulate compound leaf architecture**.** The spatial domain of PHAN expression defines leaflet positioning along the rachis, and its restriction diminishes the adaxial identity of the leaf primordium, resulting in a morphological transition from pinnate to palmate compound leaves in Solanum lycopersicum [15]. These findings establish as a central regulator linking meristem maintenance with lateral organ initiation. Consistent with these conserved functions, recent research has shown that mutagenesis of GmAS1/2 genes markedly affects leaf development and morphology in soybean [16].
Beyond its role in leaf polarity development, the AS1 gene also functions as a developmental and environmental signal integrator, regulating floral organ abscission zones, fruit patterning, flowering time, and leaf hyponasty, while also mediating defenses against fungi, bacterial pathogens and powdery mildew [17,18,19,20,21,22,23,24]. In M. truncatula, PHAN also regulates petiole and rachis length, while in S. lycopersicum, PHAN potentially modulates leaflet initiation [15,25].
While the role of the AS1 gene is well-established in diploid model systems like Arabidopsis, A. majus, and Z. mays, their evolutionary conservation and regulatory diversification in polyploid crop species remain largely unexplored. Soybean, as an ancient paleopolyploid species, is not only a cornerstone of global agriculture as the leading source of plant-derived oil and protein for food and feed, but also a key contributor to sustainable farming systems through biological nitrogen fixation. With its long history of domestication and broad adaptability, soybean holds significant economic, nutritional, and ecological value worldwide [26,27]. In addition to its agricultural significance, soybean serves as an excellent model for investigating the functional divergence of developmental regulators following gene duplication, through mechanisms such as subfunctionalization or neofunctionalization [28,29,30,31,32,33]. As leaf morphology strongly influences photosynthetic efficiency and yield, elucidating the adaptation of AS1 genes in a complex polyploid genome is essential for bridging the gap between foundational molecular genetics and agricultural trait improvement.
To address this gap, the present study identifies four GmAS1 genes and comprehensively analyzes their structural and functional conservation relative to AS1. Expression analyses showed that GmAS1a was preferentially expressed in shoot apices, GmAS1b was broadly expressed with high levels in reproductive organs and shoot apices, GmAS1c was predominantly expressed in leaves, and GmAS1d showed uniformly low expression across tissues. Functional complementation of the Arabidopsis as1 mutant by GmAS1 genes highlights their conserved biological function, while distinct expression patterns suggest regulatory divergence. Furthermore, cis-regulatory analysis of their promoters provides new evidence for regulatory diversification potentially linking developmental regulation with environmental adaptation. This work represents the first integrative characterization of the AS1 genes in a polyploid legume, offering new insights into how gene duplication maintains essential developmental functions while enabling regulatory flexibility during crop evolution.
2. Results
2.1. Phylogenetic, Gene Structure, and Conserved Motif Analysis of AS1 Genes
To elucidate the evolutionary relationships of the AS1 gene across plant lineages, twelve representative species were selected, spanning Bryophyta (Marchantia polymorpha), Pteridophyta (Selaginella moellendorffii), gymnosperms (Ginkgo biloba), basal angiosperms (Amborella trichopoda), and diverse angiosperms such as A. thaliana, A. majus, M. truncatula, G. max, S. lycopersicum, Z. mays, Triticum aestivum and Oryza sativa. Phylogenetic analysis revealed that the AS1 gene is absent in algae but present in both mosses and ferns (Figure 1A). This distribution pattern indicates that AS1 likely originated early in land plant evolution, after their divergence from algal ancestors. Its conservation across the land plant lineages suggests AS1 plays a fundamental role in the development and morphological differentiation of land plants. The retention of a single-copy AS1 gene in most of these lineages further implies strong evolutionary constraints, possibly due to its involvement in essential regulatory pathways critical for adaptation to terrestrial environments. In contrast, multiple copies of AS1 were identified in soybean and wheat, with four genes in soybean which are designated as GmAS1a (Glyma.07G132400), GmAS1b (Glyma.18G181300), GmAS1c (Glyma.03G081900), and GmAS1d (Glyma.18G151400), respectively [16].
Analysis of AS1 gene structures revealed notable variation among species, ranging from single-exon genes without UTRs to multi-exon forms with both 5′ and 3′ UTRs (Figure 1B). Despite these structural differences, AS1 proteins were highly conserved, with Motifs 6 and 1 present across all species, while a few motifs showed lineage-specific loss (Figure 1C). In conclusion, these findings indicate that although the structure and motif composition of AS1 have diversified across plant lineages, several motifs remain highly conserved, reflecting their critical role in the evolution and functional maintenance of AS1 proteins.
2.2. Collinearity Analysis of the AS1 Genes Among A. thaliana, M. truncatula, and G. max
The AS1 gene is typically present as a single copy in most plant species; but four copies were identified in G. max, likely due to two rounds of whole-genome duplication events in its evolutionary history. To trace their origins, collinearity analyses were conducted among A. thaliana, M. truncatula, and G. max. The results showed that only GmAS1a and GmAS1b retained syntenic relationships with A. thaliana and M. truncatula (Figure 2). This finding suggests that these two copies were derived from ancestral loci conserved through polyploidization, whereas the GmAS1c and GmAS1d likely originated from subsequent segmental duplications or chromosomal rearrangements, resulting in the loss of detectable synteny.
2.3. Sequence Conservation Among AS1 Proteins in Arabidopsis and Soybean
Multiple sequence alignment of Arabidopsis AS1 and the four soybean homologs revealed a high degree of amino acid conservation (Figure 3). Two distinct DNA-binding domains were clearly identified, showing near-identical sequences across all five proteins, indicating strong evolutionary constraint on these regions. In contrast, the C-terminal portions exhibited pronounced sequence divergence, suggesting potential diversification in regulatory roles or protein–protein interaction.
The sequence logo analysis further supported these observations (Figure S2). Consistent with the alignment, residues within the DNA-binding domains were highly conserved, displaying strong positional constraints and minimal substitutions, underscoring their essential function in DNA recognition and transcriptional regulation. In conclusion, these findings indicate that while the N-terminal and central domains of AS1 proteins are strongly conserved and functionally indispensable, the C-terminal region exhibits greater sequence plasticity, potentially contributing to functional diversification among soybean GmAS1 paralogs.
2.4. Expression Pattern of GmAS1 Genes
To characterize the spatial expression divergence among GmAS1 paralogs, qRT-PCR analysis was performed to quantify their transcript levels in roots, stems, leaves, flowers, pods, and vegetative shoot apices. The four GmAS1 genes exhibited distinct tissue-specific expression patterns (Figure 4A). GmAS1a was most strongly expressed in the vegetative shoot apices, with lower levels in stems and flowers. GmAS1b was broadly and highly expressed, with the highest levels in flowers, pods, and the vegetative shoot apices, moderate expression in stems and leaves, and weak expression in roots. GmAS1c was mainly expressed in leaves, with moderate levels in flowers. GmAS1d exhibited uniformly low expression across all tissues. Collectively, these patterns suggest subfunctionalization, with GmAS1a specialized in the vegetative shoot apices, GmAS1b broadly active, and GmAS1c associated with leaf development, while GmAS1d shows no strong tissue preference, possibly undergoing functional redundancy or silencing.
Consistent with these quantitative data, RNA in situ hybridization further confirmed the spatial expression pattern of the four GmAS1 genes. Their transcripts were mainly detected in the developing leaf primordia, young leaves, stipules, and stem vascular bundles (Figure 4B–I). Transverse sections further corroborated their accumulation in these tissues (Figure 4J–M).
2.5. Genetic Complementation Analysis
To evaluate the function conservation of AS1 between soybean and Arabidopsis, the coding sequences of GmAS1a, GmAS1b, GmAS1c, and GmAS1d were individually driven by the Arabidopsis AS1 promoter and introduced into the as1 mutant. PCR-based genotyping verified the mutant background and confirmed the presence of each GmAS1 transgene, and no fewer than three independent complementation lines were obtained for each construct. Phenotypic analyses showed that expression of each of the four soybean AS1 paralogs restored wild-type leaf morphology (Figure 5, Figures S3 and S4). These results demonstrate that the core developmental function of AS1 is evolutionarily conserved between soybean and Arabidopsis. Moreover, the capacity of all four soybean paralogs to complement the as1 mutant highlights their high degree of functional conservation, suggesting strong evolutionary constraints that have preserved GmAS1 function following duplication.
2.6. Prediction of Cis-Acting Elements
Analysis of the 3.5 kb promoter region upstream of the GmAS1 genes start codon using the PlantCARE database revealed a dense distribution of cis-acting elements associated with light response, hormone signaling, growth and development, and stress response (Figure 6). Light-responsive elements were the most abundant, with 18 core motifs identified, including MRE, GATA-motif, Box 4, AE-box, gap-box, and G-box, suggesting that the expression of GmAS1 genes is tightly regulated by light signaling networks. The promoter also contains multiple hormone-responsive elements, including jasmonic acid (CGTCA-motif, TGACG-motif), auxin (AuxRR-core, TGA-element), gibberellin (GARE-motif, P-box), and abscisic acid (ABRE) response elements, among a total of 12 types, indicating potential regulation by multiple phytohormones. In addition, six elements related to growth and development processes were identified, such as O2-site (endosperm-specific expression), circadian (circadian rhythm regulation), and RY-element (seed development regulation), indicating possible roles in specific developmental stages or tissues. Notably, the promoter contains six types of stress-responsive elements, including MBS (drought), LTR (cold), WUN-motif (wounding), TC-rich repeats (defense and stress response), GC-motif (hypoxia), and ARE (anaerobic induction). Based on the reported regulatory roles of MYB transcription factors in biotic and abiotic stress responses [34], these findings suggest that GmAS1 genes may play a key role in regulating soybean responses to abiotic stresses such as drought and low temperature.
3. Discussion
The soybean genome, shaped by multiple duplication events, provides an excellent model for investigating the evolution of developmental genes. As an ancient paleopolyploid, soybean has undergone at least two rounds of whole-genome duplication (WGD), the most recent occurring approximately 13 million years ago [32]. Despite extensive gene loss following these events, about 70–75% of soybean genes remain in duplicated copies [32,33]. This extensive genomic redundancy facilitated the expansion of the GmAS1 gene, yielding four homologs that have likely undergone subfunctionalization and regulatory diversification after duplication [35]. This trend of post-duplication divergence is not merely transcriptional but extends to the underlying regulatory mechanisms, and is heavily influenced by the mode of gene duplication. While large-scale expression profiling revealed that a majority of duplicated gene pairs display divergent expression patterns [36], further analysis shows that WGD-derived genes are significantly more prone to divergence through trans-regulatory changes, whereas small-scale duplicates (e.g., singletons, tandem) show a higher proportion of cis-regulatory divergence [37].
In this study, we systematically analyzed four GmAS1 paralogs, combining phylogenetic, structural, expression, and functional analyses to elucidate their evolutionary conservation and potential regulatory divergence. Our results reveal that, while all four GmAS1 paralogs retain highly conserved protein domains and developmental functions, they have undergone regulatory diversification reflected by distinct tissue-specific expression profiles and promoter compositions. These findings provide novel insights into how gene duplication in polyploid crops promotes the balance between evolutionary constraint and regulatory innovation.
3.1. Evolutionary Conservation of AS1 Function
Comparative phylogenetic and sequence analyses demonstrated that AS1 homologs are deeply conserved across land plants, from bryophytes to angiosperms, indicating its early establishment as a fundamental developmental regulator. The strong conservation of two MYB DNA-binding domains across soybean and Arabidopsis AS1 proteins suggests that evolutionary constraints have maintained these regions to safeguard their molecular function. Consistent with this, all four GmAS1 paralogs successfully complemented the Arabidopsis as1 mutant phenotype, restoring nearly wild-type leaf morphology. These results underscore that the core molecular function of AS1-mediating KNOX gene repression to maintain leaf determinacy-is evolutionarily stable.
Such evolutionary stability parallels findings from other species, including Selaginella kraussiana, Antirrhinum, Z. mays and Cardamine hirsute, where AS1 orthologs can similarly rescue as1 phenotypes despite distinct leaf morphologies across species [1,12,38]. Together, these results underscore that morphological diversity across taxa is unlikely to result from changes in AS1 protein function itself, but rather from variation in its regulatory networks. This observation sets the stage for understanding how duplicated AS1 genes in soybean maintain their conserved function while evolving divergent transcriptional regulation.
3.2. Regulatory Diversification of GmAS1 Paralogs
Despite their conserved protein domains and functional redundancy in Arabidopsis, the four GmAS1 genes display distinct expression profiles, indicating regulatory divergence following duplication. GmAS1a is preferentially expressed in vegetative shoot apices, GmAS1b exhibited broad expression across both reproductive and vegetative tissues, and GmAS1c showed leaf-enriched expression, while GmAS1d maintains low transcript levels across tissues. These complementary expression domains suggest subfunctionalization, where duplicated genes partition ancestral regulatory functions to avoid redundancy and optimize developmental precision [39,40].
Promoter analysis further supports this notion. Notably, six types of stress-responsive cis-elements were detected in the GmAS1 promoters, including MBS (drought), LTR (cold), WUN-motif (wounding), TC-rich repeats (defense and stress response), GC-motif (hypoxia), and ARE (anaerobic induction). The presence of stress-related cis-elements indicates the potential involvement of GmAS1 in defense-related signaling pathways, consistent with the reported role of AS1 in mediating defenses against fungi, bacterial pathogens, and powdery mildew. Moreover, the variation in motif composition among paralogs points to divergent cis-regulatory architectures, which may enable individual GmAS1 copies to respond to distinct physiological or environmental cues. Such diversification likely provides a flexible transcriptional framework for integrating developmental and immune signaling, thereby fine-tuning growth-defense balance in soybean.
3.3. Evolutionary and Agronomic Implications
From an evolutionary perspective, the retention of four functionally conserved yet differentially regulated AS1 paralogs in soybean exemplifies how gene duplication promotes the balance between functional constraint and regulatory innovation. Such retention likely confers developmental robustness, ensuring stable leaf polarity and morphology, while allowing flexible responses to environmental or hormonal cues. This duality reflects a broader evolutionary strategy in polyploid plants, where duplicated developmental regulators are retained not for novel biochemical roles, but for expanded regulatory capacity.
Agronomically, elucidating the regulatory diversification of AS1 paralogs offers valuable insights for soybean improvement. Leaf architecture directly determines photosynthetic efficiency and yield potential. The ability to manipulate GmAS1 expression patterns through targeted cis-regulatory modifications could enable precise control of leaf form and stress adaptability without disrupting essential developmental pathways. Given its role in coordinating morphogenetic and defense pathways, fine-tuning the expression of specific GmAS1 copies may further enhance stress resilience while maintaining optimal growth and development. Future studies integrating genome editing, chromatin accessibility profiling, and environmental transcriptomics will be critical to unravel how regulatory variation in AS1 networks contributes to phenotypic plasticity in polyploid crops.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
In this study, the as1 mutant (SALK_210101C) was obtained from AraShare (https://www.arashare.cn/index/), with Arabidopsis Col-0 and the soybean cultivar Williams 82 serving as wild-types. All plants were cultivated under long-day conditions (LD, 16 h light/8 h dark) in a temperature-controlled greenhouse maintained at 22 °C. All primer sequences are listed in Table S1.
4.2. Identification of AS1 Genes in Plant Species
To elucidate the evolutionary relationships of the AS1 gene across plant lineages, twelve representative species were selected for evolutionary analysis, encompassing M. polymorpha, S. moellendorffii, G. biloba, A. trichopoda, A. thaliana, A. majus, M. truncatula, G. max, S. lycopersicum, Z. mays, T. aestivum, and O. sativa. The Arabidopsis AS1 and maize RS2 protein sequences were used as queries in BLAST searches against Phytozome v14 (https://phytozome-next.jgi.doe.gov/), the PGCP database (https://biobigdata.nju.edu.cn/pgdatabase/home), and NCBI (https://www.ncbi.nlm.nih.gov/) to identify and retrieve homologous AS1 gene and protein sequences from these species (accessed on 12 August 2025) [41].
4.3. Phylogenetic, Gene Structure, and Conserved Motif Analysis of AS1 Homologs
Multiple sequence alignment of identified AS1 protein homologs was conducted using MAFFT with default parameters [42]. Phylogenetic trees were subsequently constructed using the maximum likelihood (ML) method implemented in IQ-TREE [43]. The resulting trees were subsequently refined and visualized using iTOL [44]. Gene structures were illustrated using GSDS 2.0 (https://gsds.gao-lab.org/). Conserved motifs were identified with MEME (https://meme-suite.org/meme/tools/meme (accessed on 12 August 2025)), and the resulting file was visualized by TBtools (v2.327) [45].
4.4. Collinearity Analysis of the AS1 Genes
The genome sequences (fasta) and annotation features (gff) of A. thaliana (TAIR 10), Glycine max (v4.0) and M. truncatula (A17 r5.0) were obtained from NCBI collinearity analyses between A. thaliana and G. max, as well as between G. max and M. sativa, were conducted using One Step MCScanX module in TBtools, and these results were subsequently integrated with the Text Merge for MCScanX function in TBtools.
4.5. Mutiple Sequence Alignment of AS1 Homologs
To investigate the sequence conservation of AS1 proteins between A. thaliana and G. max, protein sequences of AtAS1, GmAS1a, GmAS1b, GmAS1c, and GmAS1d were aligned using Clustal Omega (https://www.ebi.ac.uk/jdispatcher/msa/clustalo?stype=protein (accessed on 15 February 2025)) with default parameters. The resulting alignment was visualized and formatted with ESPript 3.0 (https://espript.ibcp.fr/ESPript/ESPript/ (accessed on 15 February 2025)). To further assess residue conservation across the five proteins, WebLogo 3.7.9 (https://weblogo.threeplusone.com/) was employed to generate sequence logos, providing a graphical representation of positional amino acid frequencies. Conserved domains and functional motifs were identified using the SMART database (https://smart.embl.de/), which guided domain annotation in the alignment and sequence logo analyses.
4.6. RNA Extraction and Quantitative RT-PCR (qRT-PCR) Analysis
Vegetative shoot apices, roots, stems, and leaves of Williams 82 at 21 days after germination, as well as flowers and pods at the reproductive stage, were collected for total RNA extraction. First-strand cDNA was synthesized from 2 μg total RNA using a HiScriptII 1st Strand cDNA Synthesis Kit (Vazyme Biotech, Nanjing, China). qRT-PCR was performed using a Roche LightCycler 480 system (Roche Diagnostics International Ltd., Rotkreuz, Switzerland) in 10 μL reaction volumes containing 5 μL of 2× Quantfast Green qPCR SuperMix (Maibo Biotech, Hangzhou, China), 0.3 μL of specific primers, 0.2 μL of 50× Rox Dye, 1 μL of 1:5 dilution of reverse transcription product, and 3.2 μL of dd H_2_O. The relative expression levels of each gene were calculated by the 2^−ΔCT^ method using the housekeeping gene GmACTIN-11 (Glyma.18G290800) as the reference gene [46,47].
4.7. RNA in Situ Hybridization
Full-length CDS with 5′ UTR of GmAS1a, GmAS1b, GmAS1c, GmAS1d served as templates to synthesize digoxigenin-labeled antisense RNA probes. Three-week-old seedling vegetative shoot apices were sectioned into eight 8-μm-thick serial sections and hybridized with the probes as previously described [48]. The signals were observed using an Olympus BX63 microscope (Olympus, Tokyo, Japan) with differential interference contrast (DIC) imaging.
4.8. Genetic Complementation Analysis
The promoter region of AtAS1 (proAtAS, 2000 bp) and the full-length coding sequences (CDS) of GmAS1a (1086 bp), GmAS1b (1074 bp), GmAS1c (1074 bp), and GmAS1d (1059 bp) were amplified by PCR with Phanta Super-Fidelity DNA Polymerase (Vazyme Biotech, Nanjing, China). The CDS of GmAS1a, GmAS1b, GmAS1c and GmAS1d were individually fused by overlapping PCR to the proAtAS1, and subsequently constructed into the pCAMBIA3301-NM vector digested with BamH I and Pml I endonucleases. The recombinant plasmids were verified by Sanger sequencing to ensure sequence integrity and correct insertion orientation. Subsequently, the validated constructs were introduced into Agrobacterium tumefaciens strain EHA105 cells. Following bacterial colony PCR verification, the transformed Agrobacterium cultures were used for floral dip transformation of Arabidopsis thaliana as1 mutant plants [49]. Transformed seeds were sown in nutrient soil and were selected by three sequential spray applications of 0.002% Basta solution at the first true-leaf stage. Herbicide-surviving plants with wild-type morphology were genotyped by PCR to confirm transgene integration.
4.9. Prediction of Cis-Acting Elements
Putative promoter regions (3.5 kb upstream of the start codon) of GmAS1 genes were retrieved from the Phytozome v14 database. These sequences were subsequently analyzed for cis-regulatory elements using PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 15 February 2025)). Motif identification and visualization were subsequently conducted in TBtools.
5. Conclusions
Collectively, this study demonstrates that duplicated AS1 genes in soybean maintain conserved protein function but diversified regulation, illustrating how polyploidy drives regulatory evolution while safeguarding core developmental mechanisms. This work provides a framework for linking gene duplication to morphological innovation and environmental adaptation in complex crop genomes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hay A. Tsiantis M. The genetic basis for differences in leaf form between Arabidopsis thaliana and its wild relative Cardamine hirsuta Nat. Genet.20063894294710.1038/ng 183516823378 · doi ↗ · pubmed ↗
- 2Nicotra A.B. Leigh A. Boyce C.K. Jones C.S. Niklas K.J. Royer D.L. Tsukaya H. The evolution and functional significance of leaf shape in the angiosperms Funct. Plant Biol.20113853510.1071/FP 1105732480907 · doi ↗ · pubmed ↗
- 3Nakayama H. Leichty A.R. Sinha N.R. Molecular mechanisms underlying leaf development, morphological diversification, and beyond Plant Cell 2022342534254810.1093/plcell/koac 11835441681 PMC 9252486 · doi ↗ · pubmed ↗
- 4Byrne M.E. Barley R. Curtis M. Arroyo J.M. Dunham M. Hudson A. Martienssen R.A. Asymmetric Leaves 1 mediates leaf patterning and stem cell function in Arabidopsis Nature 200040896797110.1038/3505009111140682 · doi ↗ · pubmed ↗
- 5Timmermans M.C. Hudson A. Becraft P.W. Nelson T. ROUGH SHEATH 2: A myb protein that represses Knox homeobox genes in maize lateral organ primordia Science 199928415115310.1126/science.284.5411.15110102816 · doi ↗ · pubmed ↗
- 6Waites R. Selvadurai H.R. Oliver I.R. Hudson A. The PHANTASTICA gene encodes a MYB transcription factor involved in growth and dorsoventrality of lateral organs in Antirrhinum Cell 19989377978910.1016/S 0092-8674(00)81439-79630222 · doi ↗ · pubmed ↗
- 7Waites R. Hudson A. Phantastica: A gene required for dorsoventrality of leaves in Antirrhinum majus Development 19951212143215410.1242/dev.121.7.2143 · doi ↗
- 8Guo M. Thomas J. Collins G. Timmermans M.C. Direct repression of KNOX loci by the ASYMMETRIC LEAVES 1 complex of Arabidopsis Plant Cell 200820485810.1105/tpc.107.05612718203921 PMC 2254922 · doi ↗ · pubmed ↗