Epigenetic Regulation of NKT-Cell-Related Gene Signatures and Prognostic Implications in Oropharyngeal Squamous Cell Carcinoma
Luka Minarik, Rita Khoueiry, Mirela Leskur, Vincent Cahais, Zdenko Herceg, Merica Glavina Durdov, Benjamin Benzon

TL;DR
This study explores how NKT-cell-related genes and their epigenetic regulation affect survival in oropharyngeal cancer patients.
Contribution
The study identifies ITK, ZNF683, and ATF2 as novel prognostic markers linked to immune activation and epigenetic regulation in oropharyngeal cancer.
Findings
High NKT cell differentiation scores are significantly associated with improved overall survival in OPSCC patients.
Epigenetic changes like hypermethylation of ITK and hypomethylation of ZNF683 are observed in tumor samples.
NKT-cell-related gene signatures are linked to immune microenvironment features and potential therapeutic targets.
Abstract
This study investigates the role of natural killer T (NKT)-cell-related gene signatures and their epigenetic regulation in oropharyngeal squamous cell carcinoma (OPSCC). Transcriptomic data from 81 OPSCC patients were analysed using single-sample gene set enrichment analysis (ssGSEA), revealing that high NKT cell differentiation scores were significantly associated with improved overall survival. Three genes (ITK, ZNF683, and ATF2) emerged as key prognostic markers linked to immune activation and T-cell signalling pathways. Methylation analyses from TCGA and GEO datasets indicated hypermethylation of ITK and hypomethylation of ZNF683 in tumour samples, suggesting epigenetic regulation of these immune-related genes. The immune microenvironment showed predominant T regulatory cells, macrophages, and neutrophils, alongside high expression of IL-15, IL-18, TNFα, and TGFβ. These findings…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
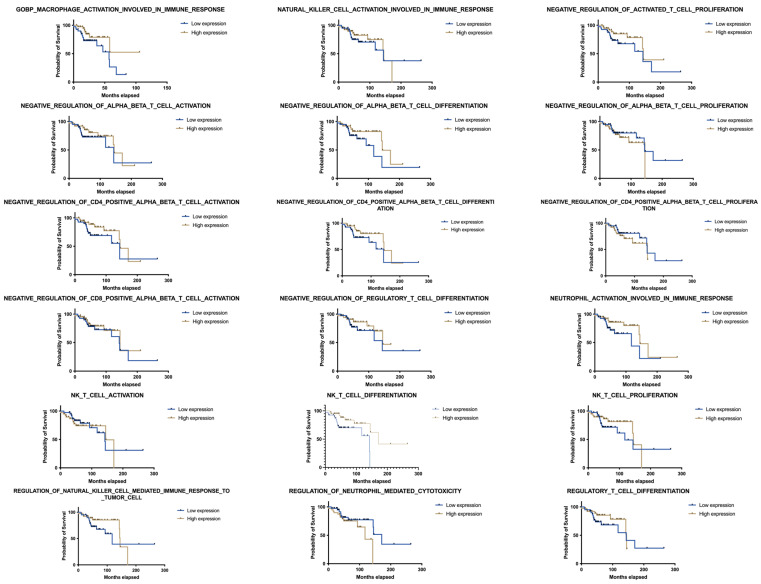 Figure 1
Figure 1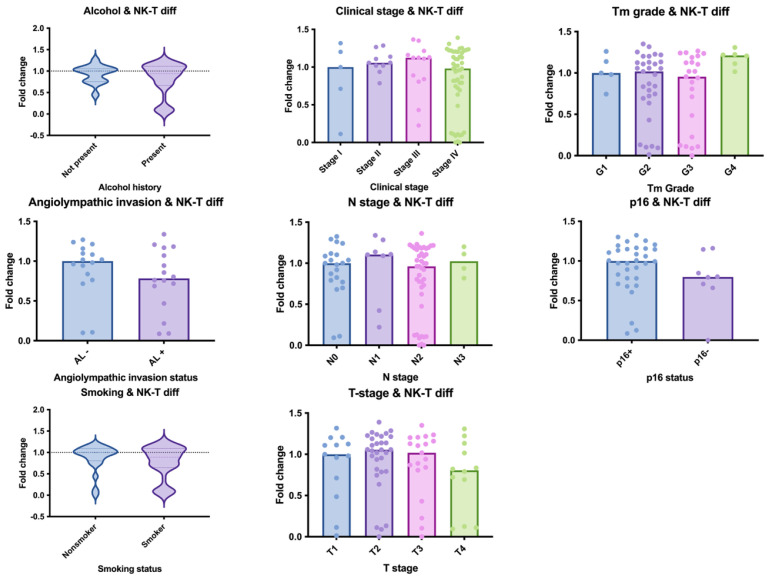 Figure 2
Figure 2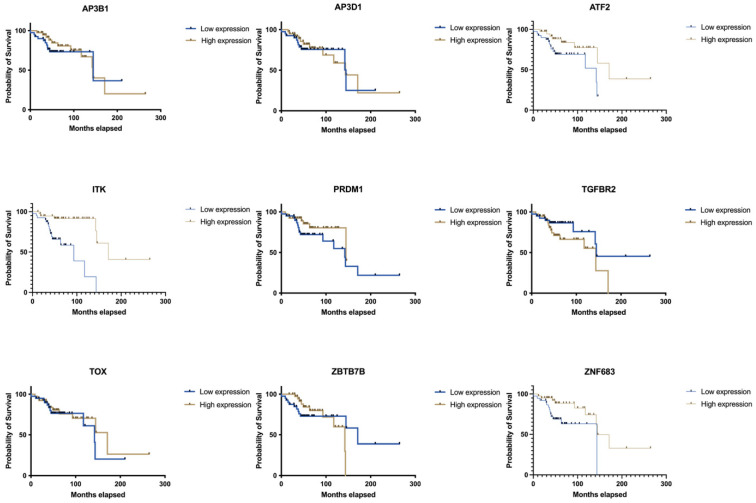 Figure 3
Figure 3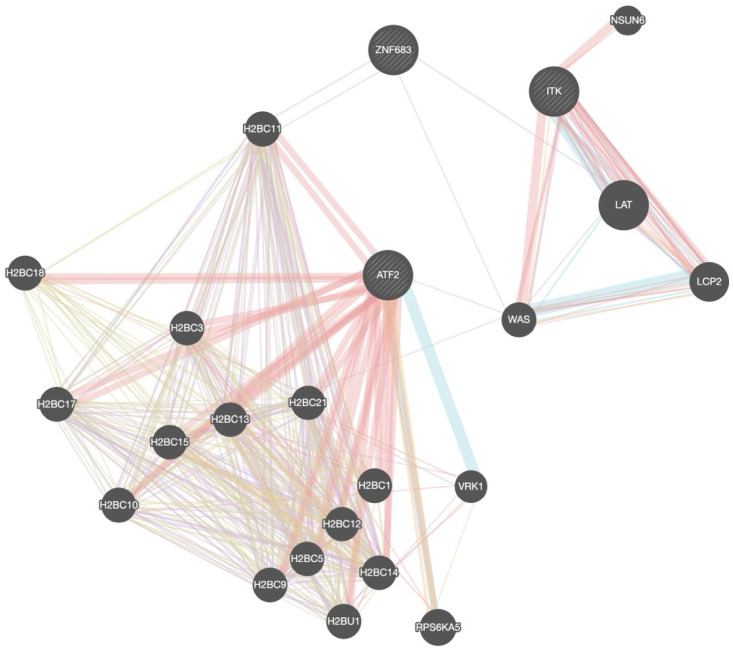 Figure 4
Figure 4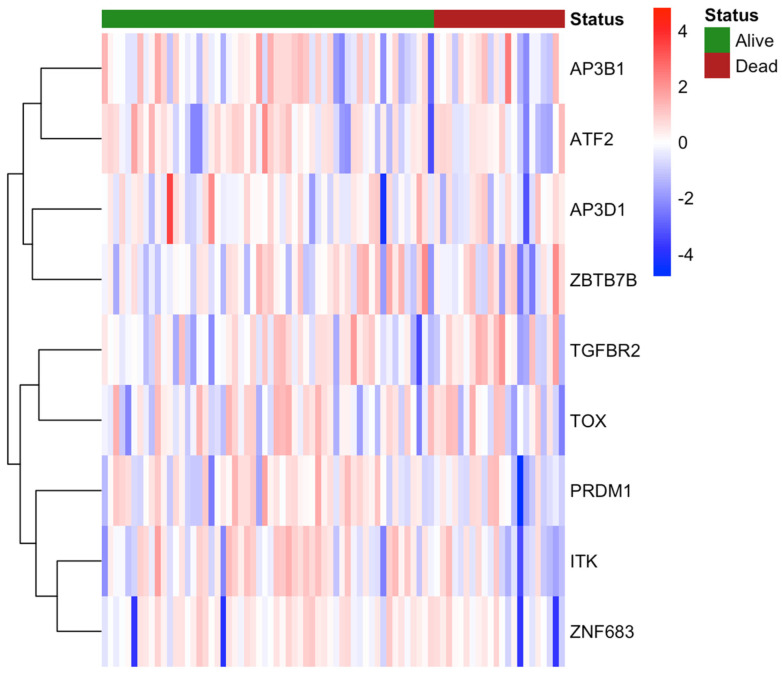 Figure 5
Figure 5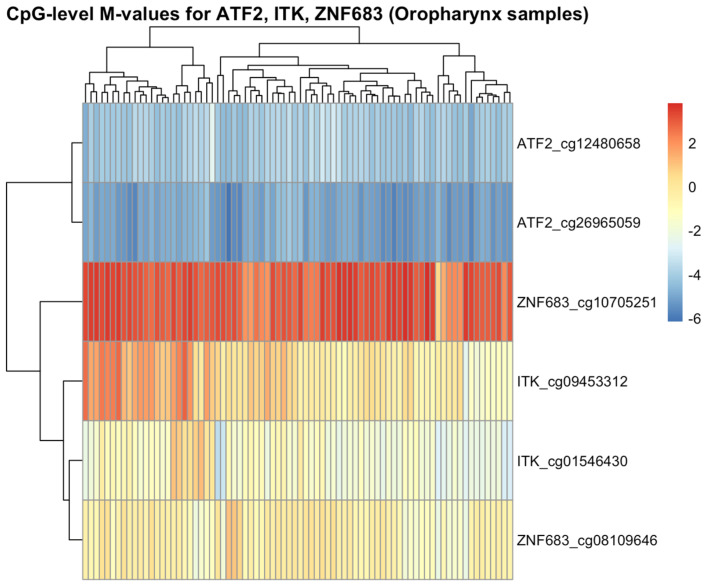 Figure 6
Figure 6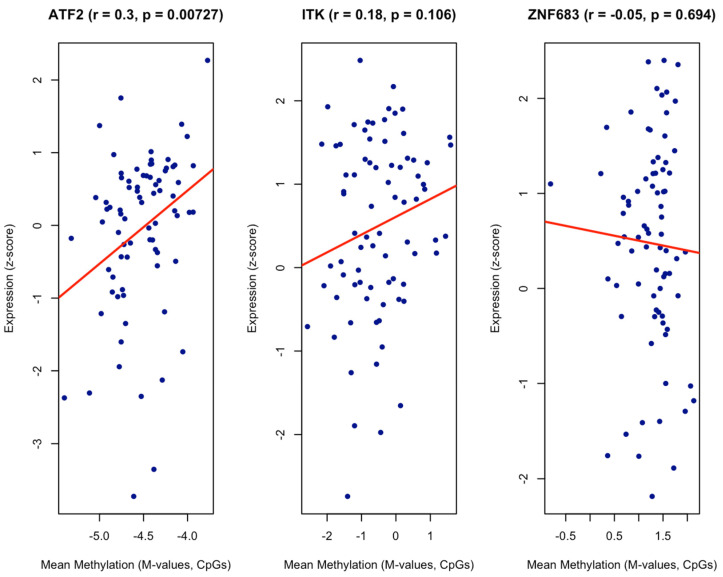 Figure 7
Figure 7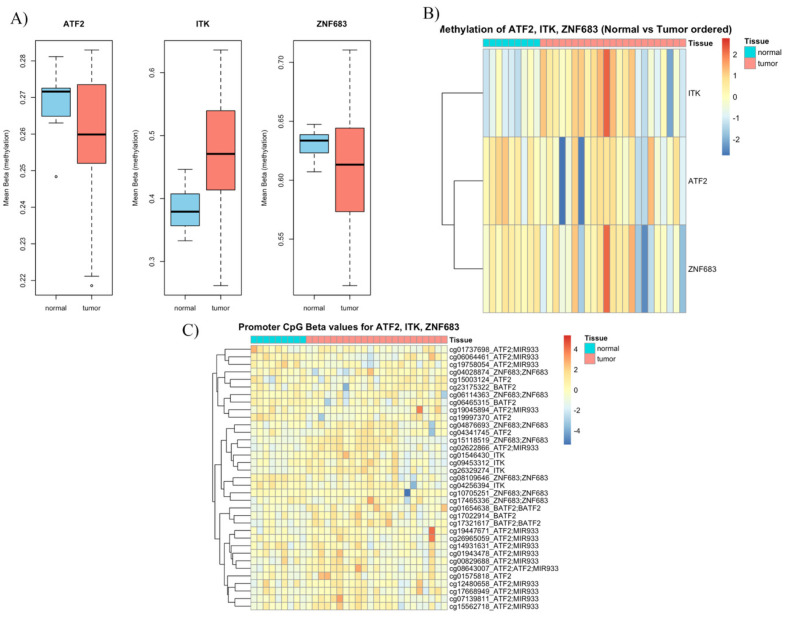 Figure 8
Figure 8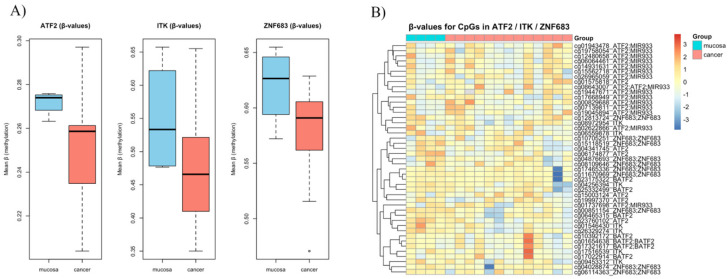 Figure 9
Figure 9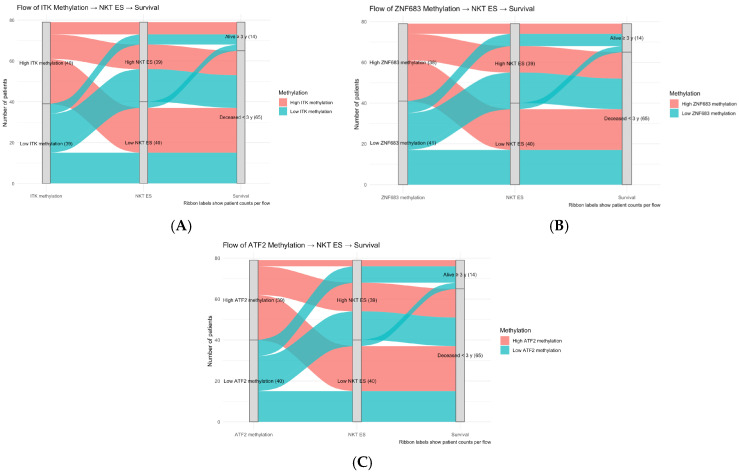 Figure 10
Figure 10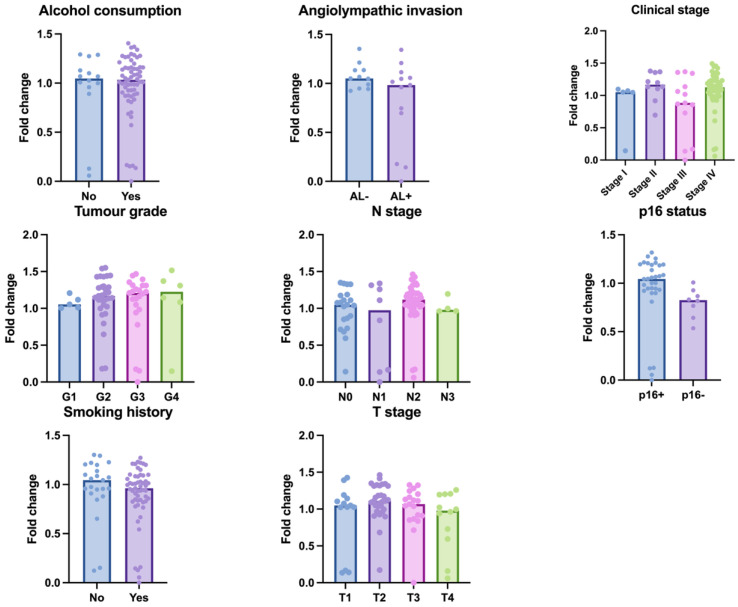 Figure 11
Figure 11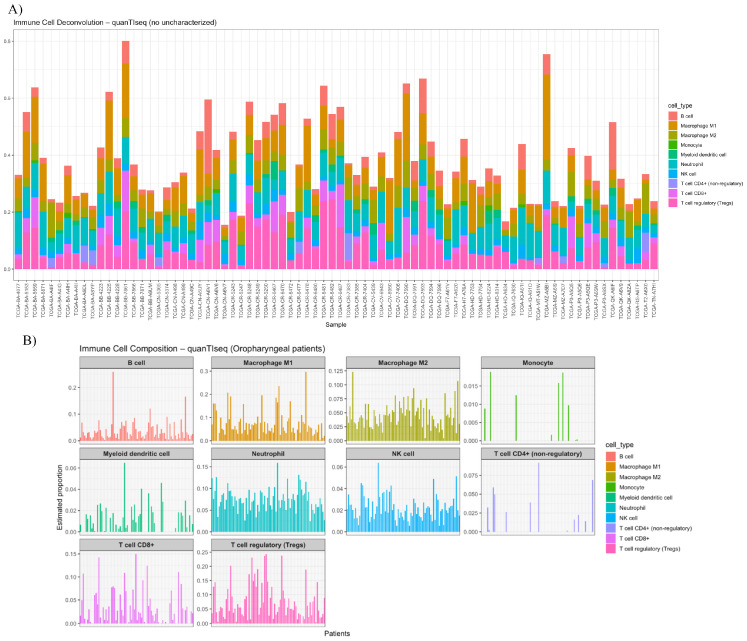 Figure 12
Figure 12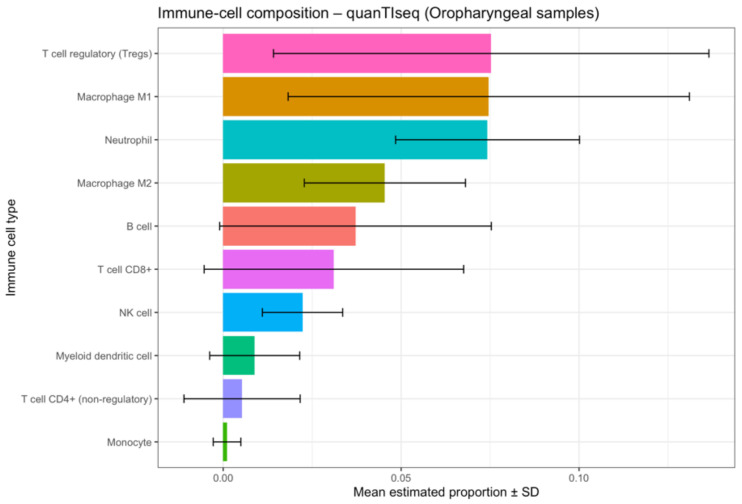 Figure 13
Figure 13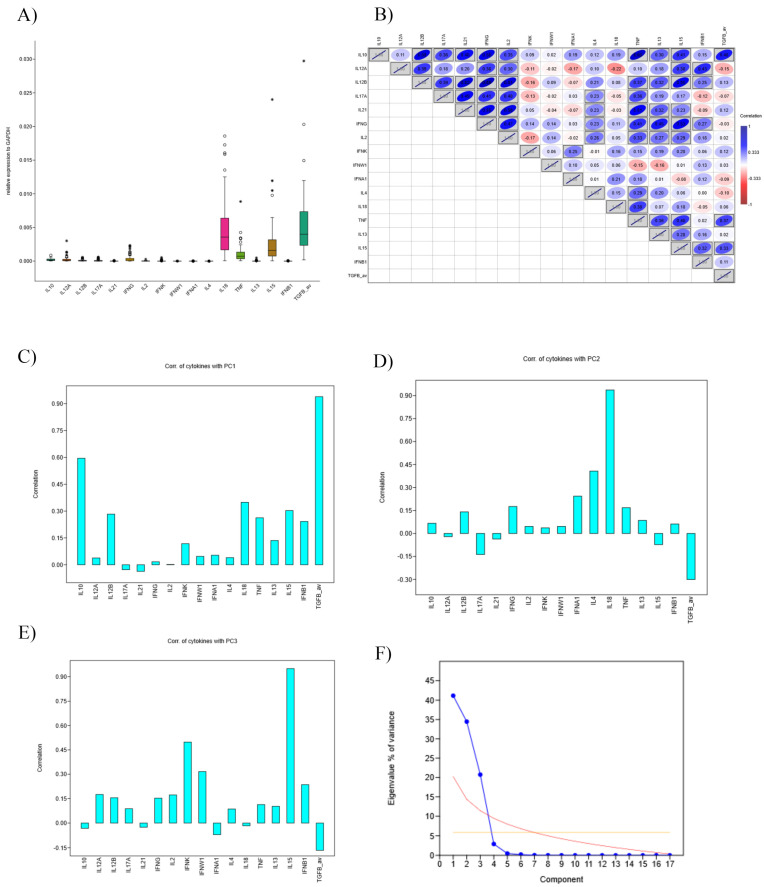 Figure 14
Figure 14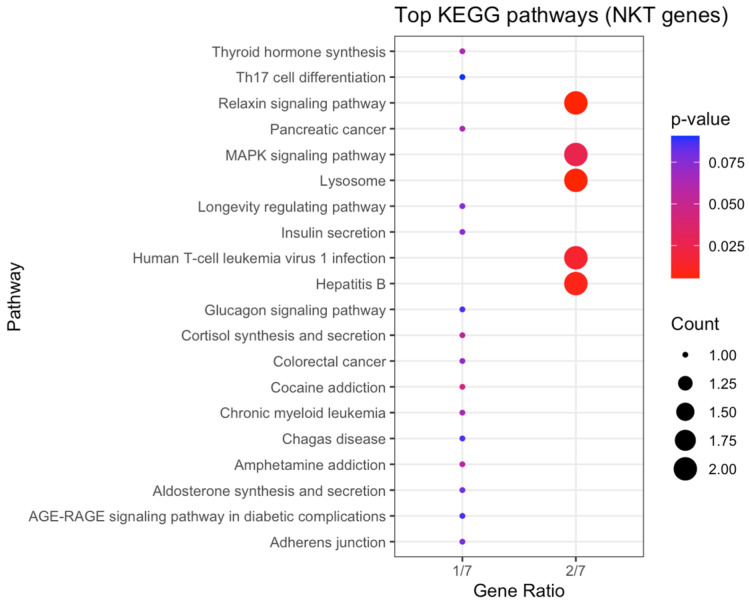 Figure 15
Figure 15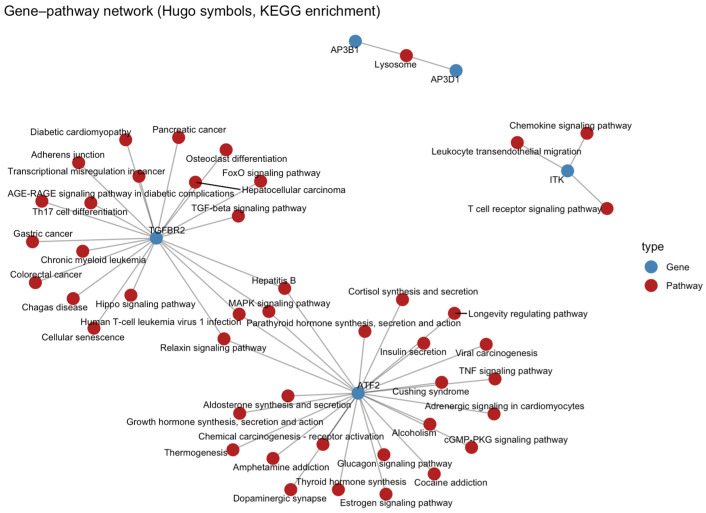 Figure 16
Figure 16Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmune Cell Function and Interaction · Cancer Immunotherapy and Biomarkers · Immune cells in cancer
1. Introduction
Head and neck cancers (HNCs) represent the fifth most common cancer worldwide, making roughly 4% of all cancer cases, with a yearly incidence of around 946,000 estimated cases [1]. Oropharyngeal cancer makes up about 23% of all HNC, with oropharyngeal squamous cancer (OPSCC) being the most common histological type [2,3]. While tobacco and alcohol consumption remain significant risk factors in many developing regions, the rising incidence of human papillomavirus (HPV) associated OPSCC in high-income countries has markedly altered both clinical presentation and prognosis [4,5]. Reflecting their clinical, genetical, and immunological differences, the American Joint Committee on Cancer (AJCC) introduced distinct staging criteria for HPV-positive and HPV-negative OPSCC in its eighth edition, recognising their differences in biological and clinical behaviours [6].
Traditionally, HNC are treated surgically with radiotherapy and platinum-based chemotherapy. However, recently, the new paradigm for cancer therapy is based on immune checkpoint inhibition, especially targeted therapy of the PD-1/PD-L1 axis and CTLA-4 [7]. This headway in cancer therapy significantly changed the way we understand OPSCC. Therapies such as pembrolizumab and nivolumab, targeting the PD-1/PD-L1 axis, have demonstrated improved survival in recurrent and metastatic HNSCC, including OPSCC, in pivotal trials like CheckMate-141, KEYNOTE-012, and KEYNOTE-040 [8,9,10]. These advances mark a paradigm shift in oncologic care, moving toward more personalised and immune-targeted approaches.
Despite initial clinical success, immunotherapy faces many challenges—one of them being that a substantial proportion of patients do not respond to immunotherapy, which may partly be explained by differences in the tumour microenvironment (TME) and biomarker expression. In addition, PD-L1 expression, tumour mutational burden (TMB), and the presence of specific T cells represent potential predictive biomarkers, but none have yet been universally accepted [11,12]. Furthermore, mechanisms of resistance are present, either innate (e.g., loss of MHC expression) or acquired during therapy [13]. This has led to an intensified search for additional indicators of immune responsiveness and tumour progression.
Given the complex interplay between tumour cells and the surrounding immune landscape, there is increasing interest in leveraging transcriptomic data to better characterise the tumour immune microenvironment (TIM) and identify immune-related gene signatures associated with disease progression and outcome [14,15]. By capturing the expression of immune-related gene signatures at the bulk or single-sample level, research can be conducted to identify patterns associated with immune infiltration, suppression, or activation. Single-sample gene set enrichment analysis (ssGSEA) is a powerful computational method that allows for the quantification of enrichment scores for specific gene sets in individual samples, enabling a more granular analysis of immune cell activity and signalling pathways [14,16]. This approach has been used to dissect the roles of various immune subsets including neutrophils, T cells, B cells, macrophages, and natural killer (NK) cells in tumour biology [16].
2. Materials and Methods
Clinicopathological, methylation and transcriptomic data from patients diagnosed with OPSCC were retrieved from the publicly available cBioPortal for Cancer Genomics database (https://www.cbioportal.org/, “Head and Neck Squamous Cell Carcinoma (TCGA, Firehose Legacy)”, accessed on the 23 December 2024). Out of a list of 530 patients diagnosed with HNC, we filtered 81 patients whose cancers were anatomically located in the oropharynx. No prior sample size calculation was performed. Gene expression profiles were analysed using ssGSEA, implemented via the GenePattern online platform (https://www.genepattern.org/), to assess the enrichment of immune-related pathways. Specifically, gene sets corresponding to activation, differentiation, and proliferation pathways of B cells, T cells, macrophages, neutrophils, NK cells, and NKT cells were evaluated. All gene sets were downloaded from the Molecular Signatures Database (MSigDB, v2024.1, https://www.gsea-msigdb.org/). The full list of immune-related pathways is available in Table 1. Survival analysis was performed for each selected immune-related pathway using Kaplan–Meier (KM) survival curves and log-rank tests. To evaluate the independent prognostic effect of NKT cell differentiation enrichment, a discrete-time survival model using a complementary log-log (cloglog) link was fitted. Bias-reduced (Firth-like) estimates were calculated to minimise small-sample bias. Covariates included HPV status (positive vs. negative) and smoking history (yes vs. no). Hazard ratios (HRs) and 95% confidence intervals (CIs) were reported on the log scale. Pathways found to be significantly associated with patient outcomes were further correlated with clinicopathological variables, including tumour grade, clinical stage, p16 status, tobacco, and alcohol consumption status. Gene expression data were downloaded as log2-transformed RNA-Seq V2 RSEM values from TCGA. Expression matrices were normalised using Z-score transformation across samples to mitigate inter-sample variability. We converted gene Z-scores into fold change values by normalising them to the median expression level of the lowest expression category. To explore the downstream molecular mechanisms associated with NKT cell-related genes, KEGG pathway enrichment analysis was performed. Genes significantly correlated with NKT differentiation ES were used as input. Pathways with an adjusted p value < 0.05 were considered significantly enriched. A gene–pathway interaction network was subsequently generated to visualise functional associations between key genes and their corresponding KEGG pathways. Searching the GEO DataSet, we collected methylation profiling data from prior studies performed on oropharyngeal cancer that uploaded data using Illumina HumanMethylation450 BeadChip (Illumina, San Diego, CA, USA). For comparing continuous variables, ANOVA or t test was used, with Welch correction if necessary. Correlations between continuous variables were performed using Spearman’s method if necessary; for reduction in dimension, principal component analysis was used. Coefficients of correlation were interpreted as suggested by Chan [17]. Statistical significance was defined as a p value less than 0.05. All statistical analyses were performed using GraphPad Prism 10 (GraphPad software, La Jolla, CA, USA, and R (v. 2024.12.1+563)).
3. Results
3.1. Transcriptomic Analysis of Immune-Related Gene Sets
Transcriptomic analysis of 81 OPSCC patients revealed differential enrichment of various immune-related gene sets. Among the immune pathways analysed using ssGSEA, NKT cell differentiation exhibited the strongest association with overall survival (OS). Patients with high NKT enrichment scores (ESs) demonstrated significantly improved OS compared to those with lower scores (median survival: 5.62 vs. 4.68 years, p = 0.015; Figure 1, Table 1). KM analysis of additional immune-related gene sets showed variable survival trends, but none reached statistical significance after multiple testing correction, highlighting the unique prognostic relevance of the NKT cell signature.
In a multivariate discrete-time survival model adjusting for HPV status and smoking, high NKT differentiation remained an independent predictor of improved overall survival (HR = 0.13, 95% CI 0.03–0.49, p = 0.003, Table 2). Neither HPV positivity (HR = 1.34, 95% CI 0.43–4.17, p = 0.612) nor smoking status (HR = 0.69, 95% CI 0.29–1.66, p = 0.410) were significantly associated with survival. These results confirm that NKT cell differentiation enrichment contributes prognostic value independently of major clinical confounders.
3.2. Clinical Data and NKT Cell Differentiation
High NKT cell differentiation ESs were significantly associated with favourable clinicopathological features. Specifically, patients in the high ES group were more likely to have lower tumour grade (p = 0.0079), earlier clinical stage (p = 0.011), and negative history of alcohol consumption (p = 0.0202) (Figure 2). No statistically significant associations were observed between NKT cell ES and tobacco consumption or p16 status.
3.3. Gene Expression and Methylation
Further analysis of gene expression within the NKT differentiation gene set identified ITK, ATF2, and ZNF683 as potential drivers of the observed prognostic signal. Among these, ITK expression was most strongly correlated with improved overall survival (p < 0.01), suggesting it may play a role in the antitumour immune activity of NKT cells in OPSCC (Figure 3, Table 3).
To explore the potential interactions and functional context of these genes, we utilized the GeneMANIA network analysis tool. The interaction network (Figure 4) revealed that ITK is closely connected to canonical T-cell signalling mediators such as LAT, WAS, and LCP2, indicating its central role in immune signal transduction. Similarly, ATF2 showed strong interaction with numerous histone H2B family members (e.g., H2BC11, H2BC13, H2BC21), confirming epigenetic or transcriptional regulatory roles. ZNF683, while less interconnected, clustered in proximity with ITK and ATF2, supporting its shared involvement in immune regulatory pathways.
These findings suggest that the prognostic signal associated with NKT cell differentiation may be driven, at least in part, by the coordinated activity of transcriptional regulators (ATF2) and TCR-associated proteins (ZNF683, ITK), implicating both epigenetic control and T-cell activation in the modulation of OPSCC progression.
Because of the potential epigenetic changes, we analysed the methylation data from our samples and correlated them with transcriptomic data of our three genes of interest (Figure 5). While CpG sites annotated to ATF2 were overall hypomethylated across oropharyngeal cancer samples (Figure 6), mean methylation levels showed a significant positive correlation (r = 0.03, p = 0.007) with gene expression (Figure 7).
To further investigate the role of epigenetics in these specific genes, we analysed methylation profile data from two studies that published their methylation meta-data (GEO access: GSE178219 and GSE98807) [18,19]. When analysing the data from Soares-Lima et al., we found that ITK was hypermethylated compared to normal tissue (p = 0.001) (Figure 8A). ZNF683 promotors were hypomethylated (p = 0.08) and ATF2 promotor methylation scores were similar in both groups (p = 0.07) (Figure 8B, Table 4). We extracted the CpG promotors and their methylation status for the three genes of interest (Figure 8C).
Consistent results were observed in the study by Nakagawa T. et al., where ZNF683 was significantly hypomethylated in the cancer cohort (p = 0.02) (Figure 9A). Their data also showed hypermethylation of ITK promoters in normal tissue (p = 0.09), and no significant difference in ATF2 methylation between the two groups (Table 5). We also visualised methylation status of CpG promotors affecting the expression of the three genes of interest (Figure 9B). We observed that patients with highly methylated genes predominantly had lower NKT ES, which also might indicate an epigenetic regulatory effect (Figure 10).
3.4. Macrophage Activation and Clinical Data
We analysed macrophage activation ES with clinical data. These patients present commonly in higher T, N, and clinical stages. We observed a 1.28-fold-higher changes in the p16+ group (p = 0.0135), implying macrophage activation in HPV+ OPSCC (Figure 11).
3.5. Deconvolution and Interleukin Expression
In order to better understand the immune cell interactions, we performed quanTIseq deconvolution analysis (Figure 12), where we approximated the number of immune cells in our samples. Our study found an overall prevalence of regulatory T lymphocytes (Treg), followed by M1 macrophage and neutrophiles (Figure 13).
To further understand the potential interactions between immune cells, we also performed an analysis of cytokine expression genes in tumours; the most expressed cytokines were TGFβ, IL-18, IL-15, and TNFα (Figure 14A). Since cytokine expression correlated with each other (Figure 14B), we performed principal component analysis (Figure 14F). The first three components explained more than 90% of variance. Principal component 1 (Figure 14C) correlated very strongly with TGFβ, moderately with IL-10 and weakly with TNFα; principal component 2 (Figure 14D) correlated strongly with IL-18 and fairly with IL-4, finally the third principal component (Figure 14E) correlated strongly with IL-15 and moderately with type-I interferons (IFNK and IFNW1). In addition, components 2 and 3 inversely correlated with TGFβ. None of the principal components correlated with survival.
3.6. KEGG Pathway Enrichment and Gene–Pathway Network Analysis
To explore downstream molecular mechanisms associated with NKT-related genes, KEGG enrichment analysis was conducted. The top enriched pathways included MAPK signalling, Th17 cell differentiation, T-cell receptor signalling, Hepatitis B, and Relaxin signalling (Figure 15).
A gene–pathway interaction network (Figure 16) revealed that ATF2 is connected to multiple signalling and metabolic pathways, including MAPK, TNF, and oestrogen signalling, suggesting its integrative role in stress and immune regulation. ITK is clustered with T-cell activation and cytokine-related pathways (chemokine signalling, T-cell receptor signalling), reinforcing its immunologic importance. TGFBR2 was connected to TGF-β and Th17 differentiation pathways, linking NKT activity with immune modulation and tumour suppression.
4. Discussion
Many studies have been carried out researching the tumour immune environment of OPSCC, primarily focusing on CD8+ and CD4+ T cell expression [20]. Other types of lymphocytes have not been as thoroughly studied. One unique population that bridges the innate and adaptive immunity are NKT cells. They are capable of direct cytotoxic activity and rapid cytokine release, playing a key role in immunosurveillance and antitumour response [21,22]. Due to their unique characteristic, NKT cells represent a specialised type of cell bridging innate and adoptive immune systems. Unlike conventional T cells, NKT cells can recognise a wide variety of antigens. Their proposed role in antitumor immunity is linked to their ability to produce IFN-γ, thereby activating NK cells and CD8+ T lymphocytes, as well as inducing IL-12 production in dendritic cells [23]. Even after adjusting for HPV status and smoking, high NKT differentiation remained an independent predictor of improved survival, highlighting its potential as a clinically meaningful biomarker beyond established prognostic factors. Given global increases in HPV-associated OPSCC and the emerging role of immune-based therapies, the findings of this study have broad relevance for biomarker development and personalised immunotherapy across diverse populations [24].
We found that NKT cell differentiation ESs were significantly associated with OS in OPSCC, suggesting that NKT activity plays a critical role in shaping patient outcomes. Supporting this finding, an in vivo study by Dhodapkar MV et al. demonstrated reversible functional defects in NKT cells among patients with progressive multiple myeloma, implicating NKT cells in the regulation of tumour growth [25]. Similarly, Molling JW et al. measured poorer overall survival outcomes among patients diagnosed with HNSCC with lower levels of circulating invariant NKT cells in 47 peripheral blood samples [26]. To our knowledge, no prior study looked at the transcriptomic expression of NKT cells in OPSCC. An immunohistochemical study conducted by Wagner S et al. described an overall better survival for patients that had a positive NK cell infiltration, compared to patients lacking NK cell infiltration [27]. Likewise, Stangl et al., in their study on HNSCC, confirmed that lower numbers of infiltrating NK cells had the highest negative predictive value on OS and disease relapse [28].
Building on these clinical and environmental findings, we described three proteins involved in the process of NKT cell differentiation significantly associated with improved OS in OPSCC: ATF2, ZNF683, and ITK.
ATF2 (Activation transcription factor 2) is a member of the basic region-leucine zipper transcription factor family and regulates numerous genes essential for cellular function. ATF2 has also been described to regulate histone H2B and H4 acetylation, regulating CRE-dependent transcription [29]. The role of ATF2 in cancer progression and outcome is controversial. For instance, Duffey et al. described that decreased expression of ATF2 in patients with HNSCC lead to an in vivo chemo resistance; interestingly, it did not affect tumour growth [30]. This is probably due to ATF2 having a role in DNA repairment. Supporting a tumour-suppressive role, Maekawa et al. demonstrated that a mouse exhibiting a heterozygote loss of Atf2 showed an increased susceptibility for mammary tumours [31]. Contrary to these findings, other studies showed an oncogene potential in solid tumours like pancreatic carcinoma and bladder cancer [32,33]. We observed that ATF2 CpG sites were globally hypomethylated in OPSCC. However, relative increases in methylation were associated with higher ATF2 expression. Since increased ATF2 expression was associated with improved overall survival, these findings suggest that methylation at ATF2 CpG sites may act as a permissive rather than repressive epigenetic mark in OPSCC.
ZNF683 (or Hobit), is a transcription factor involved in the terminal differentiation of NKT cells [34]. Its function, however, is not only restricted to NKT cells. ZNF683 is also a key regulator of tissue-resident memory T cells (Trm), a subset of peripheral CD8+ T lymphocytes that remain localised within tissues rather than recirculating through the bloodstream. Trm cells have been shown to exert potent antitumoral activity [35,36]. Consistent results were observed in the study by Nakagawa T. et al., where ZNF683 was significantly hypomethylated in the cancer cohort. These findings support our results, which suggest that ZNF683 contributes to antitumoral immune responses.
Another protein we found to have a favourable OS is ITK (IL-2-inducible tyrosine kinase), a member of the Tec family of non-receptor tyrosine kinase necessary for T-lymphocyte development, and particularly NKT cells [37]. Various studies have confirmed the role of ITK in oncogenesis, primarily in Hodgkin and non-Hodgkin lymphoma. In 2019, a case report of two siblings with biallelic ITK mutation and HPV infection resulted in epidermodysplasia verruciformis, a precancerous lesion that can lead to skin squamous cell carcinoma and Hodgkin lymphoma. Both patients exhibited a reduced number of NKT cells in their peripheral blood [38]. Our methylation data analysis of data from Soares-Lima et al. found significant hypermethylation of ITK in cancerous tissue. Similarly, we noticed a trend of hypermethylation of ITK promotor regions in OPSCC in TCGA data. However, data from Nakagawa T. et al. showed a trend towards hypermethylation in normal tissue, although not statistically significant. In contrast to our findings, Carson et al. described hypomethylation of CpG islands within ITK in melanoma, concluding that higher expression of ITK may promote carcinogenesis [39]. Similarly to the data from Nakagawa et al., Zamora-Fuentes et al. found ITK to be hypomethylated in all stages of clear cell renal cancer, classifying ITK as methylation-related oncogene; however, contrary to Carson et al., Zamora-Fuentes’ data shows that the cohort with higher expressions of ITK has a more favourable OS [40]. Furthermore, while our methylation analysis supports epigenetic regulation of ITK, ZNF683, and ATF2, the functional consequences of these modifications remain to be experimentally validated. Importantly, such DNA methylation changes are potentially reversible, which opens avenues for targeted epigenetic modulation. Recent CRISPR-based editing systems, such as catalytically inactive dCas9 fused with TET or DNMT enzymes, allow locus-specific demethylation or methylation of CpG regions [41]. Applying these tools in OPSCC models could help reverse specific epigenetic marks and directly assess their mechanistic and therapeutic impact.
Patients with higher macrophage activation ES had a higher OS and correlated with p16 positivity. In their immunohistochemical study, Tosi et al. noticed a different effect of tumour-associated macrophage dependent on HPV status, where HPV+ cancers had a favourable effect on survival, unlike the HPV cohort [42]. Snietura et al. also found that CD68+ and CD163+ cells were more expressed in tumour, with the CD163+ phenotype exhibiting negative characteristics like lower OS and loco-regional control, especially in HPV-OPSCC [43]. Our deconvolution analysis also highlights the pivotal role macrophages have on the TME. In addition to this, cytokines involved with innate immunity and Treg response seem to be the most expressed ones. Given the heterogeneity of head and neck squamous cell carcinomas, where some subtypes display markedly more aggressive behaviour while others have more favourable survival [44], further epigenetic research should be extended to other oropharyngeal and head and neck cancer subtypes to better understand how epigenetic regulation contributes to these clinical differences. Given the predominance of innate immune cells like NK/NKT cells and macrophages, we believe that further research should be focused on their interaction in the TME, and how this communication could be used as a novel therapeutic strategy or for employing therapies that are already used in other types of cancer such as type-I interferons.
The main limitation of our study is the relatively small sample size and its retrospective design. The retrospective nature of the dataset introduces potential bias in patient selection and data completeness. Despite standardised TCGA curation, residual confounding by unmeasured clinical variables cannot be excluded. While ssGSEA enables cell-type-specific pathway quantification at the individual sample level, it cannot distinguish tumour versus stromal cell origin or account for cellular heterogeneity within bulk transcriptomic data. Functional validation through in vitro and in vivo assays will be necessary to confirm the biological role of ITK, ZNF683, and ATF2 in NKT cell differentiation and OPSCC progression. Future functional experiments (e.g., CRISPR-Cas9 knockdown or overexpression assays) are warranted to clarify how these genes interact with immune cell subsets and influence tumour progression.
5. Conclusions
In conclusion, our results suggest that NKT-related gene expression signatures, particularly ITK, ZNF683, and ATF2, may serve as prognostic biomarkers in OPSCC and could represent novel therapeutic targets. We also concluded that epigenetic regulation is an important mechanism in OPSCC carcinogenesis. Given that epigenetic alterations are reversible, this raises the possibility that pharmacologic reprogramming of immune-related genes could augment antitumour immunity by activating innate cells and improve responses to immunotherapy in OPSCC. However, larger and functionally oriented studies describing these mechanisms are needed to validate these findings and clarify the contributions of NKT cells in OPSCC pathogenesis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bray F. Laversanne M. Sung H. Ferlay J. Siegel R.L. Soerjomataram I. Jemal A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries CA Cancer J. Clin.20247422926310.3322/caac.2183438572751 · doi ↗ · pubmed ↗
- 2Chaturvedi A.K. Epidemiology and Clinical Aspects of HPV in Head and Neck Cancers Head Neck Pathol.20126162410.1007/s 12105-012-0377-022782220 PMC 3394159 · doi ↗ · pubmed ↗
- 3Maschio F. Lejuste P. Ilankovan V. Evolution in the Management of Oropharyngeal Squamous Cell Carcinoma: Systematic Review of Outcomes over the Last 25 Years Br. J. Oral Maxillofac. Surg.20195710111510.1016/j.bjoms.2018.12.00630665664 · doi ↗ · pubmed ↗
- 4Chaturvedi A.K. Engels E.A. Pfeiffer R.M. Hernandez B.Y. Xiao W. Kim E. Jiang B. Goodman M.T. Sibug-Saber M. Cozen W. Human Papillomavirus and Rising Oropharyngeal Cancer Incidence in the United States J. Clin. Oncol.2011294294430110.1200/JCO.2011.36.459621969503 PMC 3221528 · doi ↗ · pubmed ↗
- 5Maluf F.V. Andricopulo A.D. Oliva G. Guido R.V. A Pharmacophore-Based Virtual Screening Approach for the Discovery of Trypanosoma cruzi GAPDH Inhibitors Future Med. Chem.201352019203510.4155/fmc.13.16624215344 · doi ↗ · pubmed ↗
- 6Amin M.B. Greene F.L. Edge S.B. Compton C.C. Gershenwald J.E. Brookland R.K. Meyer L. Gress D.M. Byrd D.R. Winchester D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to Build a Bridge from a Population-based to a More “Personalized” Approach to Cancer Staging CA Cancer J. Clin.201767939910.3322/caac.2138828094848 · doi ↗ · pubmed ↗
- 7Pardoll D.M. The Blockade of Immune Checkpoints in Cancer Immunotherapy Nat. Rev. Cancer 20121225226410.1038/nrc 323922437870 PMC 4856023 · doi ↗ · pubmed ↗
- 8Gillison M.L. Blumenschein G. Fayette J. Guigay J. Colevas A.D. Licitra L. Harrington K.J. Kasper S. Vokes E.E. Even C. Check Mate 141: 1-Year Update and Subgroup Analysis of Nivolumab as First-Line Therapy in Patients with Recurrent/Metastatic Head and Neck Cancer Oncologist 2018231079108210.1634/theoncologist.2017-067429866947 PMC 6221824 · doi ↗ · pubmed ↗