Two new species of Pestalotiopsis (Amphisphaeriales, Pestalotiopsidaceae) causing needle blight of Pinus massoniana in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2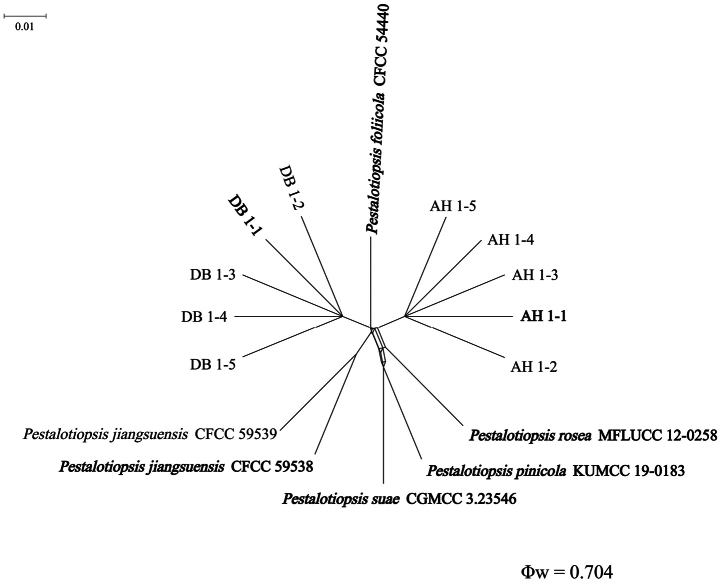 Figure 3
Figure 3 Figure 4
Figure 4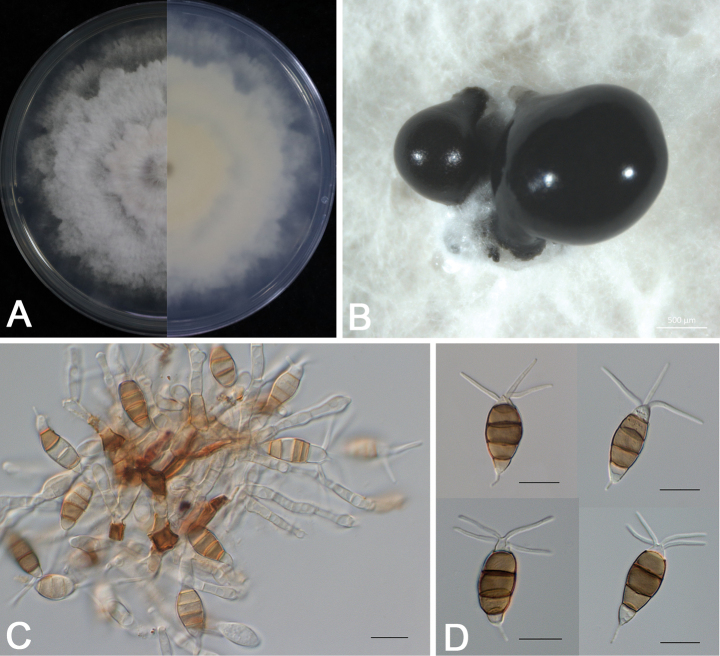 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7| Locus | PCR primers (forward/reverse) | PCR thermal cycles (annealing temperature in bold) |
|---|---|---|
|
| ITS5/ITS4 | 94 °C: 3 min, (94 °C: 45 s, 55 °C: 45 s, 72 °C: 1 min) ×35 cycles, 72 °C: 10 min |
|
| EF1-728F/EF1-986R | 94 °C: 3 min, (94 °C: 45 s, 55 °C: 45 s, 72 °C: 1 min) ×35 cycles, 72 °C: 10 min |
|
| T1/Bt-2b | 94 °C: 3 min, (94 °C: 45 s, 56 °C: 60 s, 72 °C: 1 min) ×35 cycles, 72 °C: 10 min |
| Species | Strain numberb | Host | Origin | GenBank accession numberc | ||
|---|---|---|---|---|---|---|
|
|
| TEF | ||||
|
|
| China | ||||
|
| ICMP 6088T |
| Fiji | |||
|
| LC6301T |
| China | |||
|
| HJAUP C1644.221T |
| China | |||
|
| HJAUP C1644.222 |
| China | |||
|
| IFRDCC 2397T |
| China | |||
|
|
| China | ||||
|
| SAUCC224004T |
| China | |||
|
| SAUCC224005 |
| China | |||
|
| CGMCC 3.23550T |
| China | |||
|
| CBS 331.92T |
| Singapore | |||
|
| CBS 434.65T |
| USA | |||
|
| CBS 114126T | New Zealand | ||||
|
| CBS 114193T | Australia | ||||
|
| CBS 124463T |
| Slovakia | |||
|
| CGMCC 3.18151T |
| Thailand | |||
|
| CBS 170.26T |
| New Zealand | - | ||
|
| MFLUCC 12-0277T |
| China | |||
|
| ZHKUCC23-0826T |
| China | |||
|
| ZHKUCC23-0827 |
| China | |||
|
| CSUFTCC 08T |
| China | |||
|
| HJAUP C1804.221T |
| China | |||
|
| HJAUP C1804.222 |
| China | |||
|
| CGMCC 3.23544T |
| China | |||
|
|
| China | ||||
|
| CBS 186.71T |
| Italy | |||
|
|
| China | ||||
|
|
| China | ||||
|
|
| China | ||||
|
|
| China | ||||
|
| BRIP 72970T |
| Australia | - | - | |
|
| MFLUCC 12-0273T | China | - | - | ||
|
| MFLUCC 12-0268T | China | ||||
|
| CBS 118553T |
| Colombia | |||
|
|
| China | ||||
|
| HJAUP C1724.221T |
| China | |||
|
| HJAUP C1724.222 |
| China | |||
|
| CGMCC 3.23548T |
| China | |||
|
| CBS 143421T | Australia | - | |||
|
| MFLU 14-0208T |
| New Zealand | - | ||
|
| CBS 115587T |
| China | |||
|
| CBS 143904 |
| New Zealand | |||
|
| LC3232T |
| China | |||
|
| MFLUCC12-0287T | China | ||||
|
| HGUP 4037T |
| China | |||
|
| MFLUCC 18-0913T | Thailand | ||||
|
| MFLUCC 10-0149T |
| Thailand | - | ||
|
| HMJAU 60190 |
| China | - | ||
|
| MFLUCC 18-0932T |
| Thailand | - | ||
|
| IFRDCC 2439T |
| China | |||
|
| HJAUP C1742.221T |
| China | |||
|
| HJAUP C1742.222 |
| China | |||
|
| BRIP 66615T |
| Australia | |||
|
| SAUCC230046T |
| China | |||
|
| CGMCC 3.23471 |
| China | |||
|
|
| China | ||||
|
| NTUCC 17-009T |
| China | |||
|
| MFLUCC 12-0054T |
| Thailand | |||
|
| CGMCC 3.23545T |
| China | |||
|
| HJAUP C1729.221T |
| China | |||
|
| HJAUP C1729.222 |
| China | |||
|
| IFRD 411-014T |
| China | |||
|
| NOF 3175T |
| Canada | |||
|
| E72-04 |
| Brazil | |||
|
| CBS 114127T | Australia | ||||
|
|
| China | ||||
|
|
| China | ||||
|
|
| China | ||||
|
| CBS 114491T | USA | ||||
|
| HJAUP C1638.221T |
| China | |||
|
| HJAUP C1638.222 |
| China | - | ||
|
| CBS 115391 |
| Portugal | |||
|
| CBS 265.33T |
| Netherlands | |||
|
| CBS 336.97T | Soil | Papua New Guinea | |||
|
| MFLUCC 20-0135T |
| Thailand | |||
|
| MFLUCC 12-0270T | Unidentified tree | China | |||
|
| MFLUCC 12-0259T | Unidentified tree | China | |||
|
| MFLUCC 12-0657T |
| Italy | |||
|
|
| China | ||||
|
|
| China | ||||
|
| LC4399T | China | ||||
|
| LC6636T |
| China | |||
|
| KNU-PT-1804T |
| Korea | |||
|
| NCYUCC 19-0355T |
| China | |||
|
|
|
|
|
|
|
|
|
|
|
|
| |||
|
|
|
|
| |||
|
|
|
|
| |||
|
|
|
|
| |||
|
| CBS 442.67T | Kenya | ||||
|
| CBS 114138T | New Zealand | ||||
|
| MFLUCC 16-0260T | Thailand | ||||
|
| CBS 121417T | South Africa | ||||
|
| HGUP4057T |
| China | |||
|
| MFLUCC 12-0271T | China | ||||
|
| ZHKUCC 22-0159 |
| China | |||
|
| ZHKUCC 22-0160 |
| China | |||
|
|
| China | ||||
|
|
|
|
|
|
|
|
|
|
|
|
| |||
|
|
|
|
| |||
|
|
|
|
|
|
|
|
|
|
|
|
| |||
|
| LC4344T | China | ||||
|
| HJAUP C1790.221T |
| China | |||
|
| HJAUP C1790.222 |
| China | |||
|
| CBS 102220T |
| Malaysia | |||
|
| HJAUP C1639.221T |
| China | |||
|
| HJAUP C1639.222 |
| China | |||
|
| NTUPPMCC 18-165T | China | ||||
|
| NTUPPMCC 22-012 | China | ||||
|
| CGMCC 3.18250T |
| China | |||
|
| SS1-033I |
| Canada | - | - | |
|
| CBS 144.97T |
| Netherlands | |||
|
| MFLUCC12-0279T |
| China | |||
|
| Chinese yew | China | ||||
|
| Chinese yew | China | ||||
|
| CSUFTCC 16T |
| China | |||
|
| CSUFTCC 10T |
| China | |||
|
| SAUCC232203T | - | China | |||
|
| SAUCC232204 | - | China | |||
|
| TAP1100T |
| Japan | |||
|
| NTUCC 17-011T |
| China | |||
|
| CBS 130973T |
| Australia | |||
|
| SY17A |
| China | - | ||
|
| CBS 353.69T |
| Denmark | |||
|
| MAFF 240993T |
| Japan | |||
|
| MFLUCC 16-0255T | Thailand | ||||
|
| CBS 331.96T | Coastal soil | Papua New Guinea | |||
|
| CBS 278.35 |
| Thailand | |||
|
| SAUCC230093T |
| China | |||
|
| YB28-2 | Mango | China | |||
|
| ZHKUCC 23-0873T | - | China | |||
|
| MEAN 1092T |
| Portugal | |||
|
| KUMCC 19-0183T |
| China | |||
|
| COAD 2165T |
| Brazil | |||
|
| CBS 393.48T | - | Portugal | |||
|
| ZHKUCC 23-0807T |
| China | |||
|
| ZHKUCC 23-0808 |
| China | |||
|
| SAUCC367701T |
| China | |||
|
| SAUCC367702 |
| China | |||
|
| MFLUCC 17-0416T |
| Thailand | |||
|
| IFRDCC 2399T |
| China | |||
|
|
| China | ||||
|
| CGMCC 3.23549T |
| China | |||
|
| MFLUCC 12-0258T | China | ||||
|
| CBS 176.25T | China | ||||
|
| MFLUCC 13-0399T |
| Italy | - | - | |
|
|
| China | ||||
|
| DLH 2019a |
| China | |||
|
| MFLUCC 12-0314T |
| Thailand | |||
|
| CGMCC 3.18244T |
| China | |||
|
|
| China | ||||
|
| SAUCC003804T | Soil | China | |||
|
| SAUCC003806 | Soil | China | |||
|
| SAUCC231201T |
| China | |||
|
| CBS 356.86T |
| Chile | |||
|
| CBS 144035T |
| Australia | |||
|
| CGMCC3.23546T |
| China | |||
|
| Chinese yew | China | ||||
|
| Chinese yew | China | ||||
|
| CBS 114161T | Australia | ||||
|
| CBS 141.69T | Soil | Pacific Islands | |||
|
| MFLUCC 17-1616T |
| Thailand | |||
|
| IFRDCC 2240T |
| China | |||
|
|
| China | ||||
|
| MFLUCC 12-0276T | China | - | |||
|
| MFLUCC 12-0274T | China | - | |||
|
| HHL-DG |
| China | - | ||
|
| HJHB1 |
| China | |||
|
| HJHB5 |
| China | |||
|
| LC4553T |
| China | |||
|
| ZHKUCC 23-0825T | - | China | |||
|
| CBS 114178T |
| Zimbabwe | |||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Plant chemical constituents analysis · Yeasts and Rust Fungi Studies
Introduction
Pinus massoniana Lamb., as a unique native tree species in China, is widely distributed in the subtropical region (Lu et al. 2022). Adapted to arid and barren soil, P. massoniana needles and roots can be used in traditional Chinese medicine (He et al. 2009); its trunk can be used for papermaking and industrial construction (Liu et al. 2015); and it has a strong regeneration ability that can promote carbon cycling (Kang et al. 2006). Therefore, it has extremely high economic and ecological value (Yang et al. 2020). Needle blight was found in Quanjiao County, Anhui Province, and Liuzhou City, Guangxi Province, which seriously endangered the health of P. massoniana needles and must be monitored.
Pine wood nematode disease, caused by Bursaphelenchus xylophilus (pinewood nematode; PWN), is one of the main diseases affecting P. massoniana (Wu et al. 2022). As early as 1975, the Guizhou Forestry Institute investigated local diseases of P. massoniana forests and found diseases such as red blight, pine blister rust, pine leaf rust, and sooty blotch. Pine rot, a world-famous disease of young P. massoniana, is caused by Cenangium ferruginosum Fr. (Fu 1989). The branches of damaged pine trees become wrinkled due to water loss, resin oozes from the trunk, and infection at the base of the trunk causes bark rot. The infection leads to the death of the whole plant. Cronartium quercuum infects the trunk, lateral branches, and bare roots of pine trees and forms galls, which seriously affect the seed-setting ability of P. massoniana (Li et al. 1995). Zhuang (2001) recorded the pathogen Diplodia sapinea (Fr.) P. Karst., which causes shoot blight of P. massoniana. Fusarium oxysporum Schltdl. can cause damping-off of P. massoniana (Luo and Yu 2020). Wilt disease, also known as damping-off disease, begins with the withering of the top needles and, over time, leads to root rot until the whole seedling dies. This pathogen has been proven to cause plant withering (Tint 1945) and root rot (Bloomberg 1971). Pseudofusicoccum kimberleyense Pavlic, T.I. Burgess & M.J. Wingf. and Pse. violaceum Mehl & Slippers can cause P. massoniana branch blight (Li et al. 2023). Needle blight of P. massoniana is caused by Pestalotiopsis funerea (Desm.) Steyaert, which makes the needles dry up, die, and defoliate early (Liang et al. 2002).
As endophytes, plant pathogens, or saprophytes, Pestalotiopsis species are widely distributed throughout the world, mainly in tropical and temperate regions, and have a wide range of host plants (Bate-Smith and Metcalfe 1957; Guba 1961; Sutton 1980; Jeewon et al. 2003; Maharachchikumbura et al. 2011). Pestalotiopsis Steyaert was segregated from Pestalotia by Steyaert (1949). Its stable characteristics include the length and width of conidia, the length and color of the median three cells, and the number and length of apical appendages (Jeewon et al. 2002). However, excessive overlap of conidia makes it difficult to identify species solely by morphological characteristics (Maharachchikumbura et al. 2011). Although some additional taxonomic features can be used as identification bases for Pestalotiopsis species—such as pigmentation of intermediate cells, an important feature that distinguishes P. funerea from Pestalotiopsis triseta (Moreau & M. Moreau) Steyaert—they still have great limitations (Griffiths and Swart 1974). In the past, Steyaert (1949) and Guba (1961) divided species with variegated conidia into two groups according to the color of intermediate cells. However, with the development of identification technology, phylogenetic analyses based on multi-gene sequences have shown that classifying species by the color of intermediate cells is unreliable (Maharachchikumbura et al. 2014).
As a plant pathogen, needle blight caused by Pestalotiopsis is a common disease in young forests and is widespread and seriously detrimental. Li et al. (2024) discovered that Pestalotiopsis jiangsuensis is the pathogen of pinprick disease of P. massoniana. Pestalotiopsis funerea can infect Pinus tabulaeformis Carrière (Huang and He 2000), P. taeda (Huang 2018), P. massoniana (Maharachchikumbura et al. 2014), and other species, causing needle blight. Xu et al. (2017) reported that the pathogen causing needle blight of P. sylvestris L. was Parosela citrina Rydb. Needle blight not only leads to chlorosis of leaves and withering of branches but, in severe cases, can cause death of the entire plant, resulting in serious economic and ecological losses (Orlikowski et al. 2014; Hu et al. 2020; Monteiro et al. 2022).
In April and July 2023, needle blight samples of P. massoniana were collected in Quanjiao County, Anhui Province, and Liuzhou City, Guangxi Province, respectively. Over time, needle blight of P. massoniana occurred more frequently, and the pathogen spread quickly, resulting in the invasion of more P. massoniana forests. Therefore, the main purpose of this study was to determine the needle blight pathogen of P. massoniana, and its pathogenicity was verified by Koch’s postulates.
Materials and methods
Field survey and fungal isolation
In April and July 2023, pine needle lesions of P. massoniana were discovered in Liuzhou City and Quanjiao County, respectively. The entire P. massoniana forest was inspected, and samples of diseased needles from five trees were taken to the laboratory for further observation. After macroscopic and microscopic examination of the diseased needles, the healthy and diseased middle parts of the needles were cut with sterile scissors. The surface was disinfected in 70% ethanol for 30 s, in 1% NaClO for 90 s, and then washed three times in sterile water for 90 s each. The pine needle fragments were spread on sterile dry filter paper, dried, and inoculated onto potato dextrose agar (PDA) for 3 days in darkness. The hyphal tips of fungi growing from tissue blocks were cut and transferred to new PDA plates to obtain pure cultures.
Morphological identification
Colony morphology and pigment production were observed after the culture was grown at 25 °C for 7 days. During this period, the development of fungal spores was monitored daily. The shape and color of acervuli and conidial masses were observed using a Zeiss stereo microscope (SteRo Discovery V20, Oberkochen, Germany). A Zeiss Axio Imager A2m microscope (Carl Zeiss, Oberkochen, Germany) was used to examine the micromorphology of the strains, including the shape, color, and number of appendages of conidiophores, conidiogenous cells, and conidia.
Genomic DNA extraction, PCR, and sequencing
Genomic DNA of the fungi was extracted from the aerial mycelia of 5-day-old cultures using the cetyltrimethylammonium bromide (CTAB) method. Three genomic loci—internal transcribed spacer (ITS), partial translation elongation factor 1-alpha (TEF1), and partial β-tubulin (TUB2)—were amplified with the primers ITS1/ITS4 (White et al. 1990), EF1-728F/EF1-986R (Carbone and Kohn 1999), and T1/Bt-2b (Glass and Donaldson 1995; O’Donnell and Cigelnik 1997), respectively. The reaction conditions are shown in Table 1. PCR amplifications were performed in a thermal cycler with a 50-μL reaction volume. Each 50-μL PCR reaction contained 25 μL of Premix TaqTM (Takara Biomedical Technology Company Limited, Beijing, China), 2 μL of forward primer, 2 μL of reverse primer, 2 μL of DNA template, and 19 μL of sterile water. The PCR amplicons were purified and sequenced by Sangon Biotech (Shanghai, China).
Phylogenetic analyses
Based on partial comparison results from the NCBI GenBank nucleotide database and recent studies on Pestalotiopsis species, reference strains were selected. Neopestalotiopsis protearum (CBS 114178) was designated as the outgroup taxon. Concatenated multilocus data (ITS, TEF1, and TUB2) were used for phylogenetic analyses with maximum likelihood (ML) and Bayesian inference (BI). MAFFT version 7.313 (Katoh and Standley 2013) and BioEdit version 7.0.9.0 (Hall 1999) were used to manually align and edit DNA sequences. IQ-TREE version 1.6.8 (Nguyen et al. 2015) was used to conduct ML analyses on the multilocus alignments. The GTR+F+I+G4 substitution model and bootstrap method were applied, with 1,000 replications to infer phylogenetic relationships. RAxML bootstrap support values were considered significant at ML ≥ 70. Bayesian inference was performed using MrBayes version 3.2.6 (Quaedvlieg et al. 2014) under the GTR+I+G+F model (two parallel runs; 2,000,000 generations). Bayesian posterior probability values were considered significant at PP ≥ 0.90. The phylogenetic tree was constructed in FigTree version 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/).
Genealogical concordance phylogenetic species recognition analyses
SplitsTree version 4.14.6 and Genealogical Concordance Phylogenetic Species Recognition (GCPSR) analysis were used to assess recombination levels among closely related species. The pairwise homoplasy index (PHI) test was performed, and the new taxon of Pestalotiopsis and its most closely related species were independently evaluated. A PHI value (Φw < 0.05) indicates significant recombination in the dataset. Using the LogDet transform and splits decomposition options, the relationships among closely related species were displayed in a splits diagram.
Pathogenicity test
The pathogenicity of 10 representative isolates was tested on 33 healthy 2-year-old P. massoniana seedlings. The tested plants were obtained from the Gudong Seedling Base in Hechi, Guangxi, China. Healthy needles of P. massoniana were stabbed with sterile needles, each producing one wound, and conidial suspensions (10^6^ conidia·mL^−1^) were sprayed evenly on the wounds. Each isolate was inoculated onto three plants, while control plants were sprayed with sterile water. Inoculated and control seedlings were placed in a chamber (1.5 × 1.2 × 1.5 m) equipped with a humidifier (300 mL/h) to maintain relative humidity (RH) at 70%. The chamber was kept in a greenhouse at 25 ± 2 °C and observed continuously for 10 days. All experiments were conducted three times.
Results
Disease symptoms and fungal isolation
In April and July 2023, needle blight of P. massoniana was observed in Liuzhou City and Quanjiao County, showing two distinct symptom types. In Liuzhou, the disease presented as necrotic bands and dead areas, with necrotic bands encircling pine needles of varying sizes. These bands were dry and discolored, not easily broken, and the pine needles outside the necrotic zones remained green (Fig. 1B), with an incidence rate of approximately 70%. However, in Quanjiao County, the disease symptoms included dead needle tips or upper parts of needles that broke off easily, while the remaining portions of the needles stayed green (Fig. 1A), with an incidence rate of approximately 70%. The two sample sets from Liuzhou City and Quanjiao County were treated separately. Based on colony morphology on PDA and ITS sequence analysis, 183 Pestalotiopsis strains were isolated and identified from the two regions, with isolation frequencies of 91% and 92%, respectively. Ten representative isolates (DB 1-1, DB 1-2, DB 1-3, DB 1-4, DB 1-5, AH 1-1, AH 1-2, AH 1-3, AH 1-4, and AH 1-5) were selected for further study and deposited at the China Forestry Culture Collection Center (CFCC).
Symptoms of needle blight of Pinus massoniana found in Quanjiao County, Anhui Province (A), and Liuzhou City, Guangxi Province (B).
Phylogenetic analyses
The ten representative isolates, together with 163 additional isolates and Neopestalotiopsis protearum CBS 114178 as the outgroup, were included in multilocus phylogenetic analyses using concatenated ITS, TEF1, and TUB2 sequences (Table 2; Fig. 2). A total of 1,801 bp, including gaps, was obtained (ITS = 1–657, TEF1 = 658–1,273, TUB2 = 1,274–1,801). The tree topologies generated from the ML and BI analyses were consistent, with ML bootstrap support values greater than 70% and Bayesian posterior probabilities (BPP) greater than 0.90 shown at the nodes (ML/BI). In the phylogenetic analyses, the ten isolates were clustered into two independent clades (ML/BI = 100/1), which formed a larger branch with four ex-type strains with strong support (ML/BI = 100/1: Pestalotiopsis foliicolaCFCC 54440, P. pinicola KUMCC 19-0183, P. suae CGMCC3.23546, and P. rosea MFLUCC 12-0258). DB 1-1, DB 1-2, DB 1-3, DB 1-4, DB 1-5, and P. jiangsuensisCFCC 59538 (ex-type) clustered into a smaller branch with strong support (ML/BI = 100/1), while AH 1-1, AH 1-2, AH 1-3, AH 1-4, and AH 1-5 were placed in a different clade (Fig. 2). Based on the three-locus phylogenetic analyses and morphology, ten strains (DB 1-1, DB 1-2, DB 1-3, DB 1-4, DB 1-5, AH 1-1, AH 1-2, AH 1-3, AH 1-4, and AH 1-5) were identified as two different new species of Pestalotiopsis. The PHI test of the new species showed no significant recombination (Φw = 0.704) with their closely related taxa (P. foliicola, P. pinicola, P. suae, P. rosea, and P. jiangsuensis) (Fig. 3)
Phylogenetic relationships of Pestalotiopsis isolates DB 1-1, DB 1-2, DB 1-3, DB 1-4, DB 1-5, AH 1-1, AH 1-2, AH 1-3, AH 1-4, and AH 1-5, based on concatenated sequences of ITS, TEF1, and TUB2 regions. RAxML bootstrap support values (ML ≥ 70) and Bayesian posterior probability values (PP ≥ 0.90) are shown at the nodes (ML/PP). Neopestalotiopsis protearum (CBS 114178) was used as the outgroup. Scale bar = 0.04 substitutions per nucleotide position. Sequences from this study are shown in red, and ex-type strains are shown in bold.
Pairwise homoplasy index (PHI) test of Pestalotiopsis isolates DB 1-1, DB 1-2, DB 1-3, DB 1-4, DB 1-5, AH 1-1, AH 1-2, AH 1-3, AH 1-4, and AH 1-5, and closely related P. foliicola, P. pinicola, P. suae, P. rosea, and P. jiangsuensis, using both LogDet transformation and splits decomposition. PHI test results (Φw < 0.05) indicate significant recombination within the dataset.
Taxonomy
Pestalotiopsis
liuzhouensis
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
Li-Hua Zhu, Hui Li & D.W. Li sp. nov.
EBF4DCFB-A9B6-5BC6-9A60-861D37D86DCF
Fungal Names No: FN 572916
Etymology.
the epithet referring to the place where the holotype was collected.
Culture characteristics.
On PDA medium, the front of the colony is white, with dense aerial hyphae and complete edges. The center of the back is light yellow.
Description.
Sporadic black and gregarious conidiomata produced on PDA after 7 days under light at 25 °C, globose, semi-immersed, dark brown to black, up to 400 μm diam (Fig. 4B); Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells (7.2–)8.5–12(−13.3) × (2.5–)3.1–5.2(−5.9) µm (11.2 ± 1.3 × 3.9 ± 0.7 µm, n = 30), hyaline, ampulliform or cylindrical, and sometimes slightly wide at the base (Fig. 4C). Conidia phragmospores, (19.7–)21.1–23.4(–24.8) × (7.8–) 8.4–9.2 (–9.8) µm (22.4 ± 1.4 × 8.9 ± 0.6 µm, n = 30), fusoid, ellipsoid, straight to slightly curved, 4-septate (Fig. 4D); basal cell hyaline, obconic, thin-walled, 3.7–6.1 μm long; three median cells doliiform, wall rugose, concolorous, brown, septa darker than the rest of the cell (second cell from the base 3.7–5.6 μm long; third cell 4.1–5.8 μm long; fourth cell 4.0–6.2 μm long); apical cell hyaline, smooth-walled, conic or trapezoid, tapering toward the apex, 2.2–4.4 μm long, with 2–4 tubular apical appendages (mostly 2 and rarely 4), arising from the apical crest, unbranched, filiform, 8.7–23.4 μm long; basal appendage single, tubular, unbranched, centric, 1.5–5.1 μm long.
Pestalotiopsis liuzhouensis (DB 1-1). A. Colony on PDA after 5 days at 25 °C in the dark; B. Conidiomata and conidial masses; C. Conidiophores, conidiogenous cells, and conidia; D. Conidia. Scale bars: 500 μm (B); 20 μm (C, D).
Holotype.
China • Guangxi province, Liuzhou city, Shatang Town, Junwu Park, 24°21'45"N, 109°24'07"E (DMS), isolated from needles of Pinus massoniana, 23 July 2023, Hui Li, holotype CFCC 70485. Holotype is a living specimen being maintained via lyophilization at the China Forestry Culture Collection Center (CFCC), Chinese Academy of Forestry, Beijing, China, and ex-type DB 1-1 is stored at the Forest Pathology Laboratory, Nanjing Forestry University.
Habitat and host.
On needles of Pinus massoniana with needle blight.
Known distribution.
Liuzhou, Guangxi Province, China.
Additional specimens examined.
China Guangxi province, Liuzhou city, Shatang Town, Junwu Park, 24°21'45"N, 109°24'07"E (DMS), isolated from needles of Pinus massoniana, 23 July 2023, Hui Li, cultures: CFCC 70482 (= DB 1-2), CFCC 70479 (= DB 1-3), CFCC 70474 (= DB 1-4), and CFCC 70478 (= DB 1-5).
Notes.
Compared with P. jiangsuensis, DB 1-1 has wider conidia, 22.4 ± 1.4 × 8.9 ± 0.6 µm vs 23.4 ± 1.8 × 7.5 ± 0.5 μm. The number of apical appendages is 2–4, while there are 1–4 tubular apical appendages in P. jiangsuensis.
Pestalotiopsis
kendrickii
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
Li-Hua Zhu, Hui Li & D.W. Li sp. nov.
1CD55EBD-75E0-5085-87F1-D513E8C29B50
Fungal Names No: FN 572917
Etymology.
The epithet is named after Dr. Bryce Kendrick to commemorate his contribution to mycology.
Culture characteristics.
On PDA medium, the front of the colony is white, with dense aerial hyphae, and the colony slightly bulges in an irregular circle within 1.5 cm from the center, with wavy edges. The center of the back is light yellow, and the color gradually becomes lighter around.
Description.
Sporadic black and gregarious conidiomata produced on PDA after 8 days under light at 25 °C, globose, semi-immersed, black, up to 1.5 mm diam (Fig. 5B); Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells (5.3–)7.6–11.3(−13.6) × (2.3–)2.8–5.7(−6.2) µm (10.3 ± 2.3 × 4.6 ± 0.5 µm, n = 30), hyaline, ampulliform or pear shape, smooth-walled, and sometimes slightly wider at the base (Fig. 5C). Conidia phragmospores, (18.7–)19.8–22.3(–23.4) × (6.5–)7.0–7.8(–8.3) µm (21.1 ± 1.1 × 7.3 ± 0.5 µm, n = 30), fusoid, ellipsoid, straight to slightly curved, 4-septate (Fig. 5D); basal cell obconic, hyaline, thin-walled, 3.7–5.5 μm long; three median cells doliiform, wall rugose, light brown, concolorous, septa darker than the rest of the cell (second cell from the base 3.8–5.3 μm long; third cell 3.3–4.6 μm long; fourth cell 3.8–5.3 μm long); apical cell hyaline, smooth-walled, conic or trapezoid, tapering toward the apex, 2.7–3.8 μm long, with 3–4 tubular apical appendages (mostly 3), arising from the apical crest, unbranched, filiform, inserted at the top or side of the apical cell, 10.1–18.7 μm long; basal appendage single, tubular, unbranched, centric, 2.9–6.1 μm long.
Pestalotiopsis kendrickii (AH 1-1). A. Colony on PDA after 5 days at 25 °C in darkness; B. Conidiomata and conidial masses; C. Conidiophores, conidiogenous cells, and conidia; D. Conidia. Scale bars: 500 μm (B); 20 μm (C, D).
Holotype.
China • Anhui province, Quanjiao city, Washan forest farm, 32°05'06"N, 118°16'22"E (DMS), isolated from needles of Pinus massoniana, 26 April 2023, Hui Li, holotype CFCC 70475. Holotype is a living specimen being maintained via lyophilization at the China Forestry Culture Collection Center (CFCC), Chinese Academy of Forestry, Beijing, China, and ex-type AH 1-1 is stored at the Forest Pathology Laboratory, Nanjing Forestry University.
Habitat and host.
On needles of Pinus massoniana with needle blight.
Known distribution.
Quanjiao, Anhui Province, China.
Additional specimens examined.
China Anhui province, Quanjiao city, Washan forest farm, 32°05'06"N, 118°16'22"E (DMS), isolated from needles of Pinus massoniana, 26 April 2023, Hui Li, cultures: CFCC 70487 (= AH 1-2), CFCC 70495 (= AH 1-3), CFCC 70494 (= AH 1-4), and CFCC 70483 (= AH 1-5).
Notes.
Pestalotiopsis foliicola, P. pinicola, and P. suae have 2–3 apical appendages, whereas P. rosea has 1–3 tubular apical appendages, some of which are branched. The number of apical appendages in P. kendrickii is 3 or 4, and the appendages are unbranched, while P. jiangsuensis has 1–4 apical appendages.
Pathogenicity test.
Five days after inoculation, needles inoculated with strains AH 1-1, AH 1-2, AH 1-3, AH 1-4, and AH 1-5 exhibited obvious gray spots (Fig. 6), while those inoculated with strains DB 1-1, DB 1-2, DB 1-3, DB 1-4, and DB 1-5 showed milder yellow–brown spots (Fig. 7). Ten days after inoculation, the lesions expanded into necrotic bands. The necrotic symptoms on needles inoculated with strains AH 1-1, AH 1-2, AH 1-3, AH 1-4, and AH 1-5 were pronounced, and the lesions had almost spread throughout the needles, forming necrotic bands with an incidence rate of 100%. The spots on needles inoculated with strains DB 1-1, DB 1-2, DB 1-3, DB 1-4, and DB 1-5 turned reddish brown and developed into necrotic bands, also with an incidence rate of 100%. No symptoms developed on the control needles. Pestalotiopsis liuzhouensis and P. kendrickii were successfully re-isolated from 100% of the inoculated plants, as identified by morphological characteristics and phylogenetic analysis of ITS, thereby fulfilling Koch’s postulates.
Pathogenicity of representative isolates of Pestalotiopsis kendrickii (AH 1-1, AH 1-2, and AH 1-3) on Pinus massoniana. A. No symptoms were observed on control pine needles treated with sterile water after 5 days; B–D. Symptoms on pine needles inoculated with conidial suspensions of AH 1-1, AH 1-2, and AH 1-3 after 5 days, respectively; E. No symptoms were observed on control pine needles treated with sterile water after 10 days; F–H. Symptoms on pine needles inoculated with conidial suspensions of AH 1-1, AH 1-2, and AH 1-3 after 10 days.
Pathogenicity of representative isolates of Pestalotiopsis liuzhouensis (DB 1-1, DB 1-2, and DB 1-3) on Pinus massoniana. A. No symptoms were observed on control pine needles treated with sterile water after 5 days; B–D. Symptoms on pine needles inoculated with conidial suspensions of DB 1-1, DB 1-2, and DB 1-3 after 5 days, respectively; E. No symptoms were observed on control pine needles treated with sterile water after 10 days; F–H. Symptoms on pine needles inoculated with conidial suspensions of DB 1-1, DB 1-2, and DB 1-3 after 10 days.
Discussion
The species nomenclature of Pestalotiopsis was originally established and distinguished based on the first observation of their hosts (Jeewon et al. 2002; Lee et al. 2006). With the advancement of research, the genus has been classified using the morphological characteristics of conidia, such as the length and width of conidia, the length and color of intermediate cells, and the length and number of appendages, which have significantly improved the accuracy of species identification within the genus (Jeewon et al. 2003; Hu et al. 2007). Molecular technology has also been widely applied in fungal taxonomy and in the identification of plant fungal pathogens. Pestalotiopsis sensu lato was divided into three genera by Maharachchikumbura et al. (2014): Pestalotiopsis sensu stricto, Neopestalotiopsis, and Pseudopestalotiopsis. These three branches correspond to three conidial types—conidia with light brown or olivaceous concolorous median cells (Pestalotiopsis sensu stricto), conidia with versicolorous median cells (Neopestalotiopsis), and conidia with dark-colored concolorous median cells (Pseudopestalotiopsis). This classification further improves the accuracy of species identification and benefits studies on the species diversity of the genus (Liu et al. 2010). However, with the increasing number of new Pestalotiopsis species, additional challenges have emerged, such as conidial overlap and complexity and the absence of gene fragments in some species (Jeewon et al. 2002; Tejesvi et al. 2009). The separation of Pestalotiopsis species therefore remains a major challenge.
Pestalotiopsis is widely distributed worldwide and has numerous plant hosts (Maharachchikumbura et al. 2014). It has been reported and studied as a plant pathogen, saprophyte, and endophyte (Barr 1975; Maharachchikumbura et al. 2011). As a plant pathogen, Pestalotiopsis can cause a variety of diseases, including branch blight, leaf spots, canker, root rot, leaf chlorosis, branch withering, and fruit rot, and in severe cases can even lead to plant death, resulting in significant economic and ecological losses (Crous et al. 2011; Maharachchikumbura et al. 2014). Zhang et al. (2012) found that Pestalotiopsis camelliae can infect Camellia japonica, causing leaf withering; Karaca and Erper (2001) found that Pestalotiopsis guepinii can infect the branches of Corylus heterophylla and Juglans regia, causing the branches to turn brown and wither. Espinoza et al. (2008) found that Pestalotiopsis clavispora, P. neglecta, and P. angustata are pathogenic to the fruits of apple, kiwifruit, and blueberry, and that P. clavispora is also pathogenic to blueberry twigs. Li et al. (2024) found that P. jiangsuensis can infect P. massoniana, causing pine foliage to turn gray and wither. As an endophyte, Pestalotiopsis is typically nonpathogenic or weakly pathogenic to plant hosts (Watanabe et al. 2010).
Several reported endophytic Pestalotiopsis species can produce secondary metabolites with great potential applications in medicine, agriculture, and industry (Xu et al. 2010; Xu et al. 2014). For example, Strobel et al. (1996) isolated a strain of P. microspora from the bark of Taxus chinensis, which produces paclitaxel, an anticancer drug. Pestalotiopsis microspora has also been shown to produce another secondary metabolite with antifungal and antioxidant activities (Strobel et al. 2002). Shimada et al. (2001) isolated two plant growth regulators from the culture filtrate of P. theae. Yang and Li (2013) isolated the endophyte P. foedan from Nelumbo nucifera, whose fermentation broth produces a compound with moderate activity against tumor cell lines. Species of Pestalotiopsis are recognized as a rich source of diverse bioactive compounds and continue to show great potential for future development (Monden et al. 2013).
Interestingly, the pathogen causing needle blight of P. massoniana is not static. In 1980, Qiu et al. (1980) first discovered that the pathogen of P. massoniana needle blight was P. funerea, but Li et al. (2024) later identified another pathogen responsible for the same disease. Currently, two additional pathogens have been identified, indicating that pathogens of the same genus can exhibit diversity even on the same host. We suspect that the pathogen responsible for pinprick blight of P. massoniana may vary across regions, possibly due to geographical and environmental factors. Future research should expand the scope of investigation to better understand the relationships among these pathogens.
Supplementary Material
XML Treatment for Pestalotiopsis liuzhouensis
XML Treatment for Pestalotiopsis kendrickii
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barr ME (1975) The genus Ostreichnion.Mycotaxon 3: 81–88. 10.5962/p.413961 · doi ↗
- 2Bate-Smith EC Metcalfe CR (1957) Leucanthocyanins. 3. The nature and systematic distribution of tannin in dicotyledonous plants. The Journal of the Linnean Society.Botany 55(362): 669–705. 10.1111/j.1095-8339.1957.tb 00030.x · doi ↗
- 3Bloomberg WJ (1971) Diseases of Douglas-fir seedlings caused by Fusarium oxysporum.Phytopathology 61: 467–470. 10.1094/Phyto-61-467 · doi ↗
- 4Carbone I Kohn LM (1999) A method for designing primer sets for speciation studies in filamentous ascomycetes.Mycologia 91(3): 553–556. 10.2307/3761358 · doi ↗
- 5Crous PW Summerell BA Swart L (2011) Fungal pathogens of Proteaceae.Persoonia 27: 20–45. 10.3767/003158511 X 60623922403475 PMC 3251321 · doi ↗ · pubmed ↗
- 6Espinoza JG Briceño EX Keith LM Latorre BA (2008) Canker and twig dieback of blueberry caused by Pestalotiopsis spp. and a Truncatella sp. in Chile.Plant Disease 92(10): 1407–1414. 10.1094/PDIS-92-10-140730769572 · doi ↗ · pubmed ↗
- 7Fu LY (1989) A newly discovered disease of Pinus massoniana in this province. Hunan Forestry Science and Technology (04): 34–35.
- 8Glass NL Donaldson GC (1995) Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes.Applied and Environmental Microbiology 61(4): 1323–1330. 10.1002/bit.2604601127747954 PMC 167388 · doi ↗ · pubmed ↗