Progressive changes in coral reef communities with increasing ocean acidification
Sam H. C. Noonan, Chico Birrell, Rebecca Fisher, Katharina E. Fabricius

TL;DR
Ocean acidification is causing gradual changes in coral reef communities, with fewer calcifying organisms and more non-calcareous algae as CO2 levels rise.
Contribution
The study provides empirical evidence of progressive coral reef community shifts with decreasing aragonite saturation state, without clear tipping points.
Findings
Coral and calcareous algae cover declined by over 50% as aragonite saturation state dropped from current levels to 2.
Non-calcareous algae increased in biomass and cover with decreasing aragonite saturation.
Coral diversity, including juveniles, rapidly declined with lower aragonite saturation.
Abstract
Ocean acidification from increasing atmospheric CO2 is progressively affecting seawater chemistry, but predicting ongoing and near-future consequences for marine ecosystems is challenging without empirical field data. Here we quantify tropical coral reef benthic communities at 37 stations with varying exposure to submarine volcanic CO2 seeping, and determine the aragonite saturation state (ΩAr) where significant changes occur in situ. With declining ΩAr, reef communities displayed progressive retractions of most reef-building taxa and a proliferation in the biomass and cover of non-calcareous brown and red algae, without clear tipping points. The percent cover of all complex habitat-forming corals, crustose coralline algae (CCA) and articulate coralline Rhodophyta declined by over 50% as ΩAr levels declined from present-day to 2, and importantly, the cover of some of these groups was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
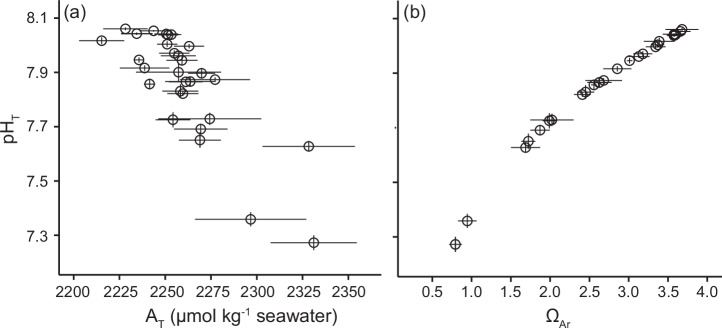 Figure 1
Figure 1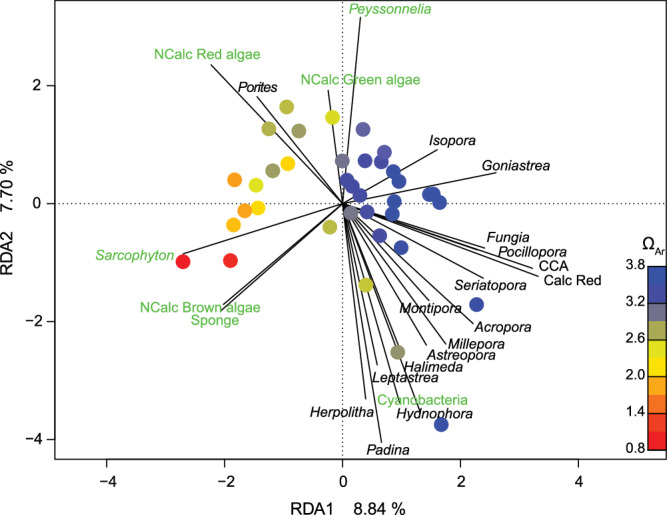 Figure 2
Figure 2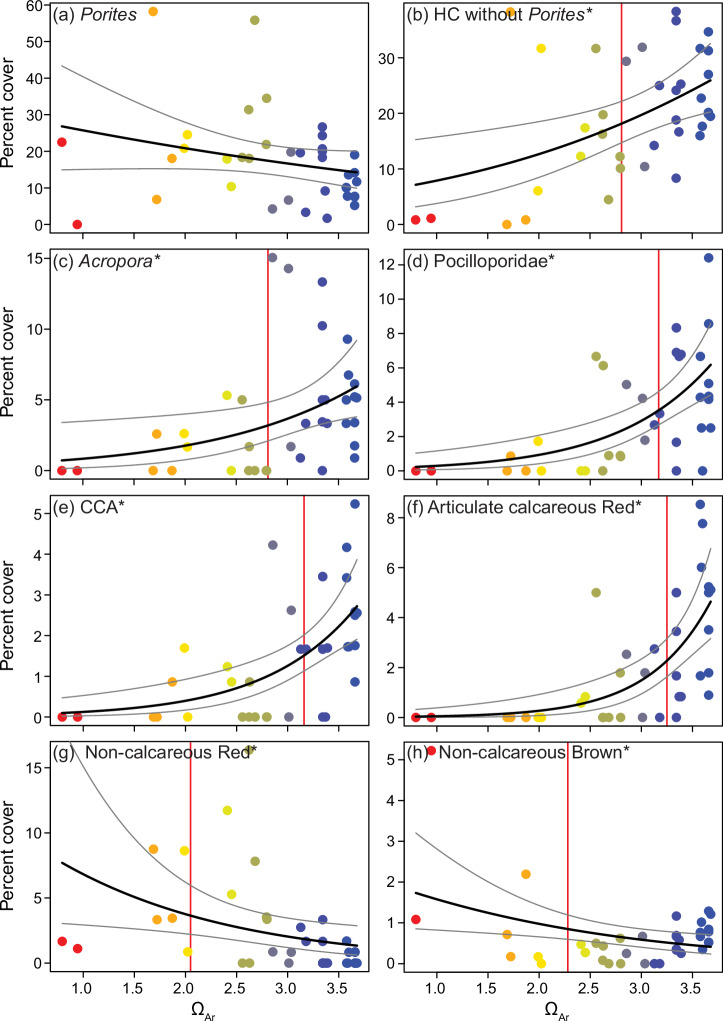 Figure 3
Figure 3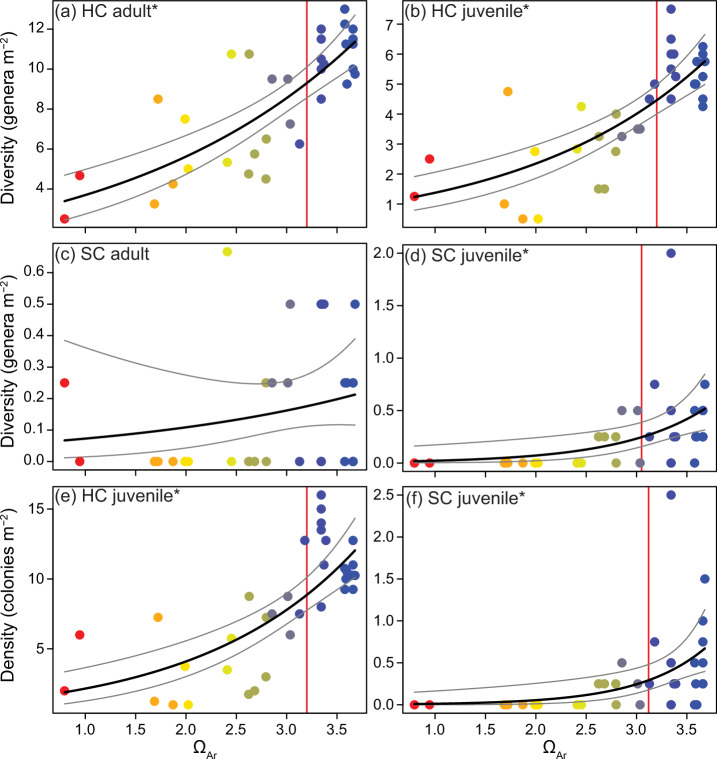 Figure 4
Figure 4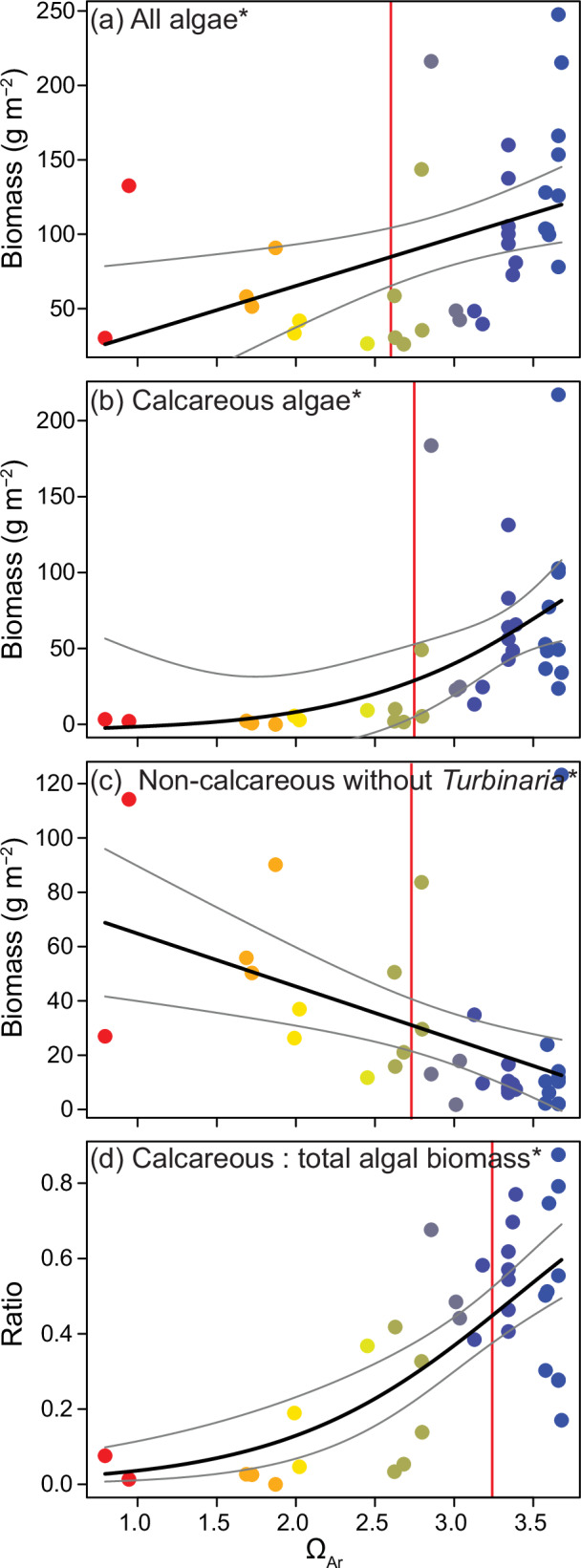 Figure 5
Figure 5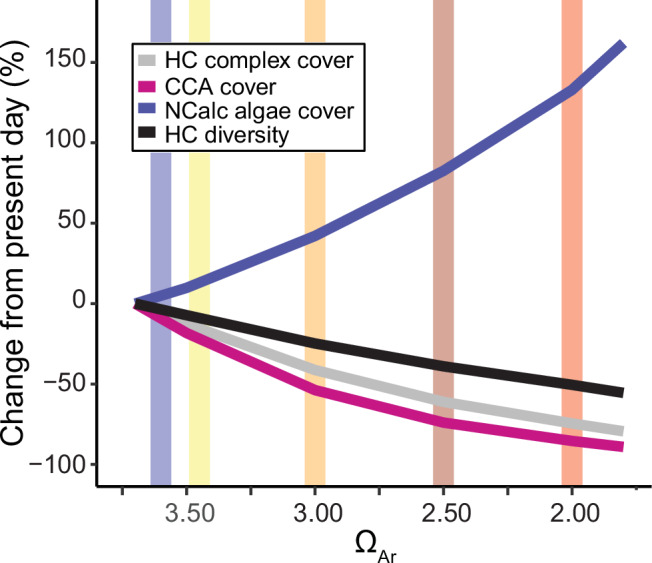 Figure 6
Figure 6- —https://doi.org/10.13039/100008344Australian Institute of Marine Science (AIMS)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCoral and Marine Ecosystems Studies · Ocean Acidification Effects and Responses · Marine and coastal ecosystems
Introduction
Human activities are increasing atmospheric carbon dioxide (CO_2_) concentrations at a rate unprecedented for at least the last 66 million years^1,2^. Approximately one quarter of this CO_2_ is absorbed by the surface waters of the world’s oceans, increasing the partial pressure of CO_2_ (pCO_2_), lowering pH and causing the chemical changes known as ocean acidification^3^. Oceanic pH is now lower than it has been for over 800,000 years, and this will likely be irreversible for millennia^4,5^. Ocean acidification is also lowering the saturation state of aragonite (Ω_Ar_), with levels declining in tropical waters at a rate of ~0.1 units per decade^6,7^. Importantly, ocean acidification is not a future-only problem, and while the ongoing chemical changes are increasingly well documented^7–9^, the present-day and near-future responses of marine communities remain less well understood.
Most ocean acidification studies compare biological responses between present-day control conditions and one or few levels of increased CO_2_, typically under artificial experimental conditions^10^. While these studies have formed the basis of our understanding of the effects of ocean acidification, they are unable to investigate any continual changes in the responses^11^. Biological responses to a changing environment may be gradual and smooth, or abrupt when thresholds are present and a sharp transition or change in the slope of the response curve occurs over a smaller environmental change^11–14^. With few treatment levels it is not possible to identify the minimum increase in CO_2_ that will result in a significant biological response^15^, as this may be occurring at CO_2_ levels below those typically used as experimental treatments^16^. Hence experimental outcomes and the interpretation of effects can depend on the number of experimental treatments, their concentrations and exposure times^10,14^. Furthermore, laboratory experiments can only approximate in situ environmental conditions and are largely void of species interactions, limiting their extrapolation to communities in natural environments. There is thus a need to study marine communities exposed to multiple levels of ocean acidification in situ, to better understand how this ongoing change is shaping communities today and in the near future.
Coral reefs are particularly vulnerable to ocean acidification because they are built by organisms with calcium carbonate skeletons (CaCO_3_), and lowered pH and Ω_Ar_ can inhibit calcification and accelerate CaCO_3_ dissolution^17^. Scleractinian corals have long been considered susceptible to these effects due to their aragonite skeletons^10,18,19^, and the high-Mg-calcite skeletons of crustose coralline algae (CCA) and other calcareous algae have been repeatedly shown to be particularly sensitive^20,21^. The reef matrix itself and sediments are largely made of CaCO_3_ and may be impacted sooner than many organisms as they have large surface-area to volume ratios and are not isolated from the adjacent seawater by tissue layers^22^. Ocean acidification also increases dissolved inorganic carbon (C_T_) concentrations, which can stimulate photosynthesis in certain circumstances^23,24^. Some non-calcareous photosynthetic reef organisms are predicted to benefit from ocean acidification if their photosynthesis is stimulated and calcareous competitors are simultaneously constrained^25^.
Investigations of the effects of ocean acidification on reef communities have been based on mesocosm experiments^26–28^ and observations at naturally occurring high CO_2_ analogues, e.g. volcanic CO_2_ seep sites^29–32^ or other oceanographic features affecting carbon chemistry^33–35^. While substantial variation exists between studies, they generally show consensus with the larger existing body of experimental work: that ocean acidification is likely to negatively affect many calcareous taxa (e.g. corals and CCA), while benefiting some non-calcareous groups (e.g. certain algae, sponges and seagrass). However, the magnitude and rate of change to reef communities under progressive ocean acidification remains largely unknown, including whether the changes will be linear or will occur at any abrupt tipping point^11^.
In this study we compare coral reef benthic communities at 37 stations representing a range of long-term ocean acidification conditions around volcanic CO_2_ seeps where strings of pure CO_2_ gas bubbles emerge from the sea floor. Except for contrasting levels of CO_2_ exposure, environmental conditions (e.g. levels of light, temperature, salinity and water flow) were similar across stations in this unique natural laboratory. We examined changes in the cover of common taxa, the density of young corals, and the biomass of different algal groups along the CO_2_ gradient. For the first time, these response curves allowed us to quantify the minimum CO_2_ increases that resulted in significant reef functional changes, and to project future changes to key coral reef benthos under different CO_2_ emission scenarios.
Results
Carbon chemistry
The stations were distributed across a field of patches with contrasting seeping activity, extending from areas unaffected by volcanic CO_2_ reflecting today’s mean carbon chemistry into patches with dense bubble streams representing predicted mean future conditions (Fig. 1; Supplementary Table 1). Ambient stations unaffected by the volcanic CO_2_ recorded total pH (pH_T_) values averaging 8.02 ± 0.01 SE. Fifteen stations within the seep area were within 0.2 pH units of ambient values, and the remaining seven stations recorded a mean pH_T_ below 7.8 (minimum station mean pH_T_: 7.27 ± 0.03). At ambient stations, total alkalinity (A_T_) averaged 2253 ± 1.16 µmol kg^-1^ seawater, and A_T_ increased along the gradient of seep exposure to 2331 µ mol kg^-1^ SW (Fig. 1). The calculated saturation state of aragonite (Ω_Ar_) was highly correlated with pH_T_, but increased relative to pH_T_ at pH_T_ values below ~7.7 due to their elevated A_T_, likely due to increased CaCO_3_ dissolution. Ω_Ar_ at stations unaffected by volcanic CO_2_ averaged 3.57 ± 0.02 while stations exposed to volcanic CO_2_ had Ω_Ar_ ranging from 3.39 to 0.79. Due to its significance for calcifying organisms and its high correlation to pH and other carbonate chemistry parameters, Ω_Ar_ was chosen as a proxy to characterise the ocean acidification level of each station. Temperature did not change along the carbon chemistry gradient (SeaFET logger data, GLM, p > 0.05), ranging from 27.5 – 28.8 °C across stations (Supplementary Table 1).Fig. 1. Carbon chemistry at each of the 37 stations along the CO_2_ gradient.Measured values of mean pH_Total_ (pH_T_) and total alkalinity (A_T_) (a), and pH_T_ and calculated aragonite saturation state (Ω_Ar_) (b). Error bars are one standard error.
Coral communities—benthic cover
A redundancy analysis indicated that changes in the reef communities across the stations were strongly related to Ω_Ar_ (PERMANOVA: Ω_Ar (1, 35), F = 2.594, p = 0.001). In the ordination plot, most stations with the highest Ω_Ar clustered closely together, with positive RDA1 axis values that were associated with an increased percent cover of many calcareous taxa, including the important reef habitat-building hard corals Acropora, Pocillopora, Seriatopora, Goniastrea and Fungia spp., as well as articulate calcareous Rhodophyta and crustose coralline algae (CCA) (Fig. 2, Supplementary Table 3). Station benthic communities with mean Ω_Ar_ values < 3.0 typically recorded negative RDA1 values (Fig. 2, Supplementary Table 3). Seep site stations with lower Ω_Ar_ were typically associated with higher cover of noncalcareous taxa including brown and red macroalgae, sponges and the soft coral Sarcophyton spp., as well as the hard coral Porites (Fig. 2).Fig. 2. Redundancy analysis ordination plot of the 37 benthic reef communities with contrasting aragonite saturation states (Ω_Ar_).Points represent station means and their colours represent their station mean aragonite saturation state (Ω_Ar_). The top 50% of the most influential vectors are shown. Vector labels for noncalcareous taxa (NCalc) are written in green, for calcareous taxa in black. All vector labels and scores are listed in Supplementary Table 3.
The in situ data documented continual changes in reef benthic communities in response to near-future ocean acidification (Fig.3, Table 1). In general, abrupt thresholds were not detected, but rather we observed continual smooth changes along the Ω_Ar_ gradient. Changes were either linear or log-linear, with different slopes or susceptibilities between taxa. The percent cover of all scleractinian corals combined did not change along the Ω_Ar_ gradient and averaged 36.15 ± 1.59% across all stations (generalised linear models, Supplementary Table 2). This was largely driven by the cover of massive Porites spp., which was unaffected by Ω_Ar_. However, without massive Porites spp., the cover of all remaining hard corals combined declined compared to ambient values, by 20% at Ω_Ar_ 3.0, and by 35% at Ω_Ar_ 2.5 (Fig. 3, Table 1). The combined cover of structurally complex hard corals (i.e. branching, corymbose and tabulate growth forms), which are important as habitat for a large proportion of reef-associated organisms, was highly susceptible. Here the no-significant-effect concentration (NSEC), being the Ω_Ar_ value where the mean response first occurs outside the 99% confidence intervals of the response at ambient Ω_Ar_ values, indicated that complex coral cover became significantly lower than control values with a reduction in Ω_Ar_ of <0.4 units (NSEC Ω_Ar_ = 3.11). This decline matched patterns in the overall complexity score for each quadrat (Table 1). The cover of the genera Acropora, Seriatopora and Pocillopora were all highly susceptible and reduced by at least 30% by Ω_Ar_ 3.0. The combined cover of non-complex hard corals (i.e. massive, submassive and encrusting growth forms) was unaffected by the Ω_Ar_ gradient, again driven by massive Porites cover. However, even some robust taxa such as Goniastrea spp., and the Merulinidae and Fungiidae declined along the Ω_Ar_ gradient (Table 1, Supplementary Fig. 2). While no clear threshold was seen, virtually no other corals besides massive Porites spp. were found at stations with a mean Ω_Ar_ < 2.0 (Fig. 3). The other hard coral families recorded (Agariciidae, Diploastraeidae, Lobophyllidae and Euphylliidae) averaged <1% cover each and were not analysed individually. Soft coral cover was also low (1.28% ± 0.36) and statistically unaffected by Ω_Ar_, although practically no soft corals were found below Ω_Ar_ 2.5. Sponge cover increased along the Ω_Ar_ gradient by >150% of control values at Ω_Ar_ 2.5.Fig. 3. Changes in benthic cover of reef coral and algal communities in relation to aragonite saturation state (Ω_Ar_).The plots show percent cover of the resilient massive Porites spp. coral (a), all hard corals excluding massive Porites spp. b the important habitat-forming corals Acropora spp. c and Pocilloporidae (d), and important algal groups: crustose coralline algae (CCA) (e), articulate calcareous Rhodophyta (Red) (f), non-calcareous Rhodophyta (g), and non-calcareous Phaeophyta (Brown) (h). Point colour represents station mean Ω_Ar_ (legend as per Fig. 2). The black line is the modelled mean, and the grey lines are 95% confidence intervals (CI). The red vertical line is the no-significant-effect concentration (NSEC). * denotes statistical significance in generalised linear models at p < 0.05 (Supplementary Table 2).Table 1. Estimates of changes in coral reef benthos along the aragonite saturation state (Ω_Ar_) gradientΩ_Ar_ 3.5Ω_Ar_ 3.0Ω_Ar_ 2.5Ω_Ar_ NSECQuadrat complexity score (1–5)3.07 (0.10)2.57 (0.08)2.15 (0.09)3.26Articulate calcareous Rhodophyta cover3.45 (0.55)1.49 (0.34)0.63 (0.26)3.25Ratio calcareous : total algal biomass0.54 (0.04)0.37 (0.04)0.23 (0.04)3.24HC adult diversity10.55 (0.51)8.55 (0.36)6.93 (0.41)3.2HC juvenile diversity5.24 (0.33)4.01 (0.22)3.07 (0.24)3.2HC juvenile density10.74 (0.81)7.79 (0.54)5.65 (0.59)3.2Pocilloporidae cover5.08 (0.72)2.91 (0.46)1.65 (0.44)3.17CCA cover2.22 (0.33)1.26 (0.21)0.71 (0.20)3.16SC juvenile density0.51 (0.11)0.24 (0.07)0.12 (0.06)3.12Complex HC cover12.73 (1.48)8.57 (0.98)5.68 (1.04)3.11Seriatopora spp. cover2.37 (0.48)1.19 (0.30)0.59 (0.27)3.11Fungiidae cover0.88 (0.18)0.49 (0.11)0.27 (0.11)3.06SC juvenile diversity0.42 (0.09)0.23 (0.06)0.13 (0.05)3.05Goniastrea* spp. cover2.22 (0.38)1.46 (0.25)0.96 (0.25)2.97Pocillopora* spp. cover2.67 (0.57)1.68 (0.36)1.05 (0.37)2.91Turbinaria* spp. biomass32.50 (4.36)23.49 (3.51)14.47 (3.95)2.88HC without Porites spp. cover24.18 (2.58)19.69 (1.85)15.86 (2.06)2.81Acropora spp. cover5.24 (1.00)3.66 (0.66)2.54 (0.70)2.81Acroporidae cover10.28 (1.67)7.55 (1.13)5.51 (1.23)2.81Calcareous algae all biomass65.52 (9.41)48.52 (7.56)31.53 (8.52)2.79Non-calcareous macroalgae cover7.07 (1.09)9.14 (1.06)11.75 (1.21)2.74Non-calcareous algae biomass without Turbinaria spp. 16.07 (5.85)25.81 (4.70)35.55 (5.30)2.73Calcareous algae cover11.70 (1.60)9.40 (1.12)7.51 (1.24)2.68Total macroalgae biomass114.08 (11.32)97.82 (9.09)81.55 (10.24)2.6Merulinidae cover6.01 (1.08)4.69 (0.74)3.63 (0.81)2.59Melanamansia spp. biomass5.70 (2.29)8.76 (1.85)11.83 (2.08)2.53Non-calcareous Phaeophyta cover4.53 (1.12)5.88 (1.10)7.60 (1.27)2.28Sponge cover1.68 (0.40)2.13 (0.39)2.70 (0.45)2.19Non-calcareous Rhodophyta cover1.50 (0.52)2.04 (0.54)2.78 (0.63)2.05Hard coral cover38.60 (2.63)37.13 (2.05)35.68 (2.29)-Massive Porites spp. cover.14.86 (2.34)16.68 (1.99)18.68 (2.23)-Parameters are ranked in order of susceptibility to Ω_Ar_ changes based on no-significant-effect-concentrations (NSEC). Mean values (±SE) of the benthic cover (%) of hard (HC) and soft (SC) corals and other benthos, the density (colonies m^-2^) and diversity (genera m^-2^) of hard and soft adult and juvenile corals, and the biomass (g m^-2^) of the algal communities are shown at Ω_Ar_ 3.5, 3.0 and 2.5. All data are presented on the measured scale. *indicates a significant response to Ω_Ar_ in the generalised linear model at p < 0.05 (Supplementary Table 2).
Coral diversity and juvenile densities
Hard coral diversity, and hard and soft coral juvenile densities and diversity, all declined along the Ω_Ar_ gradient (Fig. 4, Supplementary Table 2). Adult and juvenile hard coral diversity were highly correlated (Pearson’s correlation: t = 7.91, df = 31, p < 0.001), and both reduced significantly from ambient values as Ω_Ar_ declined by as little as 0.3 units (NSEC Ω_Ar_ = 3.20). Adult soft coral diversity was low, averaging 0.04 ± 0.16 genera m^-2^ across all sites, and was statistically similar along the Ω_Ar_ gradient. Soft coral juveniles were also sparse, but more sensitive to Ω_Ar_ declines than the hard corals. The diversity and density of soft coral juveniles declined by 50% between Ω_Ar_ 3.5 to 3.0, and none were found below Ω_Ar_ ~ 2.6. This was approximately the same Ω_Ar_ value at which adult soft coral percent cover was reduced to zero.Fig. 4. Changes in the diversity and abundance of hard and soft corals in relation to aragonite saturation state (Ω_Ar_).The diversity of adult hard corals (HC) (a), juvenile hard corals (b), adult soft corals (SC) (c), juvenile soft corals (d), and the density of juvenile hard (e) and soft corals (f) in relation to station aragonite saturation state (Ω_Ar_). See Fig. 3 legend for plot description.
Macroalgal cover and biomass
The cover of algal communities also changed along the Ω_Ar_ gradient, with a general shift from calcareous to non-calcareous taxa from ambient Ω_Ar_ into the seeps (Fig. 3). The percent cover of all calcareous algae combined at control sites declined by 20% and 35% at Ω_Ar_ 3.0 and 2.5, however susceptibility differed between taxa. The abundant heavily calcified articulate Rhodophyta were the most affected, their cover was significantly reduced between ambient Ω_Ar_ and 3.25, and none were found at <2.4. Crustose coralline algae (CCA) were also highly sensitive, and cover declined by 40% between ambient and Ω_Ar_ 3.0. Both algal groups were virtually non-existent at Ω_Ar_ ≤ 2.5. In contrast, turf algae and the weakly calcified red algae Peyssonnelia spp. were unaffected by Ω_Ar_. Total non-calcareous macroalgal cover increased as Ω_Ar_ declined. This was due to non-calcareous brown and red algae, which similarly increased cover by 65% and 85% from ambient values to Ω_Ar_ 2.5. Halimeda spp. and Padina spp. (the only calcareous green and brown algae recorded), and non-calcareous green algae each had <0.5% cover and were too low to analyse individually.
The biomass (dry weight) of all collected macroalgae combined was highest at Ω_Ar_ 3.5 and declined 15% and 30% by Ω_Ar_ 3.0 and Ω_Ar_ 2.5. This was due to high calcareous macroalgal biomass at controls, which declined by 25% at Ω_Ar_ 3.0, and by 50% at Ω_Ar_ 2.5 (Fig. 5). Calcareous algae made up 50% of total macroalgal biomass at the controls, but this ratio declined to <25% at Ω_Ar_ 2.5. Total non-calcareous macroalgal biomass was unaffected by Ω_Ar_, averaging 49.41 ± 6.08 g m^-2^ across all stations. This was due to a replacement of high Turbinaria spp. at ambient CO_2_ by Melanamansia spp. as CO_2_ increased: at Ω_Ar_ 2.5 Turbinaria spp. were almost completely absent, while Melanamansia spp. more than doubled. Total non-calcareous algal biomass, without Turbinaria spp., greatly increased along the Ω_Ar_ gradient (Fig. 5).Fig. 5. Changes in macroalgal biomass in relation to aragonite saturation state (Ω_Ar_).The biomass of all macroalgae (a), calcareous macroalgae (b), total non-calcareous macroalgae without Turbinaria spp. c and the ratio of calcareous (Calc) to total macroalgal biomass (d) in relation to station aragonite saturation state (Ω_Ar_). See Fig.3 legend for plot description.
Discussion
Our empirical data from this unique natural laboratory demonstrate the drastic rates of change in coral reef benthic communities along a gradient of declining Ω_Ar_. At present-day CO_2_, these coral reefs support a diverse community of scleractinian corals and calcareous algae. These are increasingly replaced by some non-calcareous algae, massive Porites spp. corals and sponges as CO_2_ concentrations increase. High-CO_2_ communities are also increasingly less diverse and less structurally complex. Our study uniquely served to quantify near-future changes in reef communities from ocean acidification, and to define the Ω_Ar_ values that result in a significant deviation from the reefs of today. The results add to a large body of work predicting fundamental changes to coral reef communities and functions by the end of the century, with the magnitude of change dependent on the atmospheric CO_2_ emissions pathway realised. Alarmingly, these results also show that reef community changes are likely already occurring within the range of dissolved CO_2_ levels observed on contemporary reefs^16,36^.
The few studies with sufficient levels of CO_2_ to investigate continual changes to reef communities under increasing ocean acidification have shown both linear and threshold responses. Laboratory experiments on individual organisms have shown mostly linear effects^11^, while results of community-based studies or those in situ suggest tipping points may also occur^13,16^. For example, Smith et al.^16^ and Kleypas et al.^37^ both suggest that there will be a threshold in coral and algal community change between Ω_Ar_ 3.4 and 3.6, and Ω_Ar_ 3.0 has been suggested as the limit for global reef development^37,38^. We also previously deployed standardised settlement substrate along a gradient of increasing CO_2_ at this and other seep sites in Papua New Guinea, and found newly recruited CCA communities were almost absent on these surfaces at mean Ω_Ar_ < 2.5^13^. In the present study, we found linear or log-linear changes in most parameters measured along the CO_2_ gradient, beginning immediately as Ω_Ar_ declined from ambient values, and although CCA and several complex coral taxa reached very low values at 2.5 Ω_Ar_, there was little evidence of a firm tipping point or threshold for the reef communities. Here communities began to significantly change from the present-day as Ω_Ar_ values declined by as little as 0.25 units. This immediate progressive response without thresholds highlights two important considerations. Firstly, any coral reef community thresholds in response to Ω_Ar_ declines may have already been exceeded, as global Ω_Ar_ values are now 0.5 units lower than at the advent of the Industrial Revolution^9,39^. Yet we are using present-day values as our baseline and cannot infer the shape of response curves at higher Ω_Ar_ values. Secondly, the lack of thresholds indicates that even small deviations in seawater Ω_Ar_ from present-day ambient values will continue to alter coral reef communities, and the magnitude of change that communities experience into the future will likely directly depend on the amount of CO_2_ in the atmosphere.
We documented a range of contrasting susceptibilities between taxa along the Ω_Ar_ gradient. The NSEC values indicated habitat complexity was the most sensitive metric, followed by the cover of articulate calcareous red algae, the ratio of calcareous to total algal biomass, coral diversity and juvenile coral density, then the cover of several complex habitat-forming corals and CCA. The percent cover of all hard coral taxa abundant enough for analyses, except for massive Porites spp., was shown to be negatively affected by the increasing Ω_Ar_ gradient. Importantly, hard coral diversity, in both adult and juvenile life-stages, significantly declined with a Ω_Ar_ reduction of 0.3 units. At the present rate of change, these mean Ω_Ar_ values will be reached in the tropics by the year ~2060^6,7,39^. Reef communities continued to change below this Ω_Ar_ level, and a reduction of 0.5 Ω_Ar_ units from ambient values, as expected throughout the tropics by 2100 under the emissions scenario SSP2-4.5^6,39^, was associated with considerable community change. By Ω_Ar_ 3.0, the percent cover of the highly susceptible calcareous red algae (both CCA and articulate calcareous Rhodophyta) halved, and the cover of structurally complex coral species (e.g. the Acroporidae and Pocilloporidae) reduced by 30%. These more sensitive corals were then almost absent from communities by Ω_Ar_ levels of 2.5, as predicted for the end of the century under emissions scenario SSP3-7.0. The mean Ω_Ar_ of contemporary reefs in Australia’s Great Barrier Reef ranges by 0.5 units, and reefs at the lower end of this gradient similarly record reduced CCA and coral juvenile abundances^16,36^. Hence the effects of reduced Ω_Ar_ are already manifest in some present-day reefs, and these patterns will strengthen as atmospheric CO_2_ increases.
Previous studies examining changes to reef communities exposed to elevated CO_2_ have also largely shown declines in calcareous taxa and concomitant increases in the non-calcareous^29,31,32,40^. The mechanisms responsible for community changes are not fully understood, as the response of communities to high CO_2_ is often greater than the direct physiological responses seen for individual taxa. For example, metanalyses from Kornder et al.^19^ report calcification rates are reduced by an average of 18% at Ω_Ar_ < 2.5 for all corals combined. In contrast, in the present study, communities at Ω_Ar_ 2.5 exhibited a > 30% reduction in hard coral diversity and a decline of >50% cover of the more sensitive taxa compared to ambient values. Here high CO_2_ communities were dominated by massive Porites, which can be important components of the reef framework^41^, but lack the structural complexity to provide shelter for many habitat-associated species^42–44^. The observed declines in CCA and proliferation of Melanamansia spp. and other non-calcareous macroalgae at low Ω_Ar_ also likely impacted coral communities, as CCA facilitates coral recruitment while macroalgae hinders it^45,46^. The differential responses of certain algae, for example the decline in the non-calcareous Turbinaria spp. and relative robustness of the lightly calcareous Peyssonnelia spp. at high CO_2_, require further investigation. The seep communities are likely shaped not only by the direct physiological effects of elevated CO_2_ on certain taxa, but also by indirect ecological effects that can have substantial impact and are difficult to predict based on physiological studies alone^42,43,47^.
Our empirical data show that the magnitude of change to coral reef communities attributable to ocean acidification by 2100 will strongly depend on CO_2_ emissions. By 2050, Ω_Ar_ will likely be 0.2 units lower than at present^7^, but by 2100, levels will vary greatly depending on the SSP scenario realised^6,39^. Under the most optimistic IPCC scenario (SSP1–1.9), where CO_2_ emissions are cut to net zero by 2050 and atmospheric CO_2_ will have slightly eased, coral reefs in 2100 will not be altered greatly by ocean acidification in comparison to the present day (Fig. 6). Alternatively, the most pessimistic scenario (SSP5–8.5), where present-day annual CO_2_ emissions triple by 2100, would result in drastic changes to reef communities. It is becoming increasingly unlikely that either of these extreme scenarios will occur, with more recent estimates suggesting that SSP3-7.0 may be most appropriate in impact assessment studies^48^. Under this scenario, 2100 coral reefs would see considerable declines in hard coral diversity (40%), the percent cover of CCA (70%) and complex coral species (60%), and drastic increases in the cover of non-calcareous macroalgae (80%) (Fig. 6). If emissions are limited to the more conservative SSP2–4.5, we may still see a 50% loss in CCA cover and a 40% increase in the cover of non-calcareous macroalgae by 2100. Importantly, these predictions consider the effects of ocean acidification in isolation, and coral reefs are under increasing pressure from a range of sources. Global mass coral bleaching events from marine heat waves, also driven by anthropogenic CO_2_ emissions, and other forms of disturbances from climate change, are increasing in frequency and intensity and are increasingly impacting the world’s coral reefs^49^.Fig. 6. Observed changes in key reef benthos in relation to the saturation state of aragonite (Ω_Ar_).Shown are the percent cover of complex hard corals (HC, light grey), crustose coralline algae (CCA, magenta), non-calcareous (NCalc) macroalgae (blue), and hard coral diversity (black). The vertical lines represent the Ω_Ar_ values predicted for much of the tropics for the year 2100 by Jiang et al.^39^ for the five Shared Socioeconomic Pathways emission scenarios: SSP1–1.9 (blue), SSP1–2.6 (yellow), SSP2–4.5 (orange), SSP3–7.0 (brown) and SSP5–8.5 (red)^6,39^.
These CO_2_ seep sites are not perfect representations of future coral reefs. They are small in size and lack the co-occurring elevated temperature stress expected under climate change. Seep CO_2_ levels are also characteristically variable, especially within areas with low mean pH^29^ (Supplementary Fig. 1), and effects of this variability are largely unknown for most coral reef organisms^50^. Hence extrapolating the results seen here to the future of coral reefs globally will attract some uncertainty. However, studies in situ at CO_2_ seeps have advantages over laboratory studies (e.g. organisms grow under natural conditions of ecological interactions and have life-time acclimation) and hence provide unique insights into the effects of ongoing ocean acidification on marine communities.
Anthropogenic ocean acidification has been ongoing since the start of the Industrial Revolution, and global CO_2_ emissions continue to rise largely unabated^6^. Reliable in situ data are essential to validate observed changes from ocean acidification that have likely already occurred (but are not usually factored into models or debates), and to predict how they will continue to impact coral reefs^6,16,51^. Our data clearly show that these changes will intensify in the coming decades, with reef communities shifting away from dominance by many scleractinian corals towards less diverse and structurally complex communities characterised by fewer tolerant species and non-calcareous taxa, with a raft of flow-on effects on reef-associated organisms^42,43^. The magnitude of this change will depend on our CO_2_ emissions pathway, with more emissions resulting in a greater change to present-day coral reefs. Reef resilience and recovery following disturbances will also decline, as ocean acidification reduces rates of coral recruitment^52^. Reductions in atmospheric CO_2_ levels are urgently needed to prevent further deviations from the reefs of today.
Methods
This study was conducted at the volcanic CO_2_ seep at Upa-Upasina, Normanby Island, Milne Bay, Papua New Guinea (PNG). The CO_2_ seep site has been described in detail by Fabricius et al.^29^. It is characterised by a mosaic of nearly pure CO_2_ gas seeping through the sea floor in patches of varying intensity in the shallow waters along several hundred metres of coastline. This CO_2_ locally alters the carbon chemistry of the seawater without altering the temperature or salinity^24,44^. Seeping intensity is characteristically variable over short timeframes (minutes to hours), but mean CO_2_ levels around the seep site have remained relatively constant over 6 years of sampling (e.g. Supplementary Table 2 and Supplementary Fig. 3 from^29^ and the present study). To reach areas unaffected by the volcanic CO_2_, stations were spread well beyond the area of visible seeping, up to ~500 m to the south and north of the seep along the same island fringing reef. Sampling stations were established at ~3 m depth, each marked with a small sub-surface float (n = 37). Stations were spread widely across and along the seep seascape, to capture varying seep intensities.
The carbon chemistry of each of the 37 stations was characterised in two ways. Firstly, bottle samples were collected ~twice-daily for 2 weeks at each station for pH and total alkalinity (A_T_). Where CO_2_ was elevated, one sample per station was taken during each sample period (pH n = 17–18, A_T_ n = 3–4 per station). Of the fifteen stations unaffected by volcanic CO_2_, six were investigated for their carbon chemistry (pH n = 11 – 17, A_T_ = 3–4), and the remaining nine stations were given the average carbon chemistry values of the adjacent ambient stations in further analyses. All pH samples were measured within 1 h of collection using a benchtop pH electrode (Eutech) and meter (Oakton, USA), with mV converted to pH_Total_ (hereafter pH_T_) by comparison to readings from a certified TRIS pH buffer (A.G. Dickson lab, Scripps Institution of Oceanography, USA) following standard procedures^53^. Samples for A_T_ were preserved with saturated HgCl_2_ and processed at the laboratories of the Australian Institute of Marine Science (AIMS) within 1 month of collection using open cell potentiometric titration (855 robotic titrosampler, Metrohm) following Dickson et al.^53^. A_T_ values were calculated using a Gran function and comparisons to certified reference materials were <5 µmol kg^-1^ (CRM batch 141, A.G. Dickson lab, Scripps Institution of Oceanography, USA). Secondly, pH loggers (SeaFET, Seabird USA) were deployed for ~24 h at each station, sampling every 10 min. The two pH loggers rotated through all but three of the CO_2_ affected stations (n = 19), and two of the stations unaffected by volcanic CO_2_. The pH loggers were also deployed side-by-side away from the seep in ambient seawater three times throughout the sampling period, and an offset was applied to one instrument to account for differences between loggers (~0.07 pH_T_) after comparisons with parallel bottle pH samples (*n *= 3). Estimates of station pH derived from the bottle measurements and SeaFET loggers were highly correlated (Supplementary Fig. 1) and subsequently combined for further analyses. To do so, pH data were averaged for each method after being standardised by the square root of their sample size. Here sample size was either the number of bottle samples, or the number of 10 min readings the SeaFET took over the ~24 h deployment at each station. This weighted approach avoided either dataset obscuring the other. Measured pH_T_ and A_T_ values were used to calculate the remaining carbon chemistry variables for each station using the CO2SYS macro^54^ with the constants of Dickson and Milero^55^ (Supplementary Table 1).
At each station, a series of 1 m^2^ quadrats were sampled on SCUBA to characterise benthic communities. Four quadrats were sampled at each station, however five stations unaffected by volcanic CO_2_ had only two quadrats sampled. Quadrats were placed immediately around the sub-surface floats, to ensure the measured carbon chemistry parameters from the floats were representative of the quadrat area. Quadrat placement avoided areas completely occupied by single large coral colonies, likely underrepresenting large table Acropora corals commonly found at the sites unaffected by volcanic CO_2_, as well as massive Porites spp. which dominate the seep^29^. Photographs were taken of each quadrat for analysis of benthic cover and community composition^56^. To do so photographs were overlaid with 28 evenly spaced points and the benthos underneath each was recorded to the highest taxonomic level possible (a total of 4004 observations). This was genus level for hard corals and soft corals, while macroalgae and other taxa were classified into phyla. Organisms were also categorised as being calcareous or not; calcareous being those with solid calcium carbonate supporting structures (e.g. the scleractinian corals and CCA), while non-calcareous lack these heavy deposits but includes some organisms that contain small amounts of CaCO_3_ (e.g. Peyssonnelia and Padina spp. algae and Sarcophyton soft corals). The morphology of hard corals was also recorded (e.g. branching Acropora spp., tabular Acropora spp. etc.). Visual in situ surveys were conducted in each quadrat for juvenile (<5 cm diameter) hard and soft coral density and diversity^56^. Each quadrat was further visually classified for structural complexity on a scale of 1–5, with 1 being least complex and 5 being highly complex^43^. Each type of survey was conducted by a single observer to minimise bias.
All macroalgae occurring within one quarter (i.e. 0.25 m^2^) of two of the quadrats per station were hand-collected by divers using a scraping tool. Larger areas were not sampled due to time constraints in the field. Much of the macroalgae were cryptic, occurring within the crevices of the reef where they were protected from grazing, and hence unlikely to be documented in the photo-surveys. Collections did not sample encrusting algal taxa, such as crustose coralline algae (CCA) and Peyssonnelia spp., nor turfs. Macroalgae samples were sundried, transported to AIMS, sorted into calcareous and non-calcareous taxa, and after 3 days in a drying oven at 60°C, weighed to the nearest 0.1 mg (Shimadzu AW220, Japan). The two most dominant genera (Melanamansia and Turbinaria) were also separated and weighed.
Statistics and reproducibility
All statistics were conducted in R (version 4.4.0)^57^. Ω_Ar_ values calculated from pH_T_ and A_T_ were chosen as predictors as they can strongly influence the physiology of corals and other calcareous reef taxa and were highly correlated with other carbon chemistry variables (Fig. 1). All benthic data (i.e. benthos percent cover, coral juvenile abundance and diversity, algal biomass etc.) were averaged across quadrats by station, prior to statistical analyses, as each station was represented by a single Ω_Ar_ value in models. Changes in the cover of the communities were first examined via redundancy analysis (RDA). This allowed the visualisation of multi-dimensional community data in two-dimensional space, and to test the significance of mean station Ω_Ar_ (continuous variable) on community changes via PERMANOVA.
Generalised linear models (GLM) were used to examine the changes in the percent cover of different taxa from the photo surveys, the density and diversity of hard and soft coral adult and juvenile corals, and biomass of the algal communities in relation to mean Ω_Ar_ from each of the stations. Different link functions were used in models depending on data type (Supplementary Table 2): quasibinomial for percent cover, quasipoisson for juvenile abundance and diversity counts, and gaussian for algal biomass weights. Algal weights were square-root transformed to better fit the assumed distribution, with the final model chosen based on lowest Akaike information criterion (AIC). The “Predict” function in R was used to estimate response values at Ω_Ar_ 3.5, 3.0 and 2.5, approximately representing Ω_Ar_ ambient values away from the seeps, and what is projected for tropical coral reefs in the region by the year 2100 under the Intergovernmental Panel on Climate Change’s (IPCC) shared socio-economic pathways SSP2-4.5 and SSP3-7.0^6,39^. No-significant-effect concentration (NSEC) values were also calculated for each GLM following Fisher and Fox^15^. Here the NSEC is the Ω_Ar_ where the predicted response first significantly deviates above or below (depending on the shape of the response curve) the response at an ambient Ω_Ar_ of 3.5.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Supplementary information
Transparent Peer Review file Supplementary information Description of Additional Supplementary Files Supplementary Data 1 - Algal biomass Supplementary Data 2 - Benthic percent cover Supplementary Data 3 - Adult coral diversity Supplementary Data 4 - Juvenile coral diversity Reporting summary
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lan, X., Tans, P. & Thoning. K.W. Trends in globally-averaged CO 2 determined from NOAA Global Monitoring Laboratory measurements. Version 2024-0610.15138/9N 0H-ZH 07 (2024).
- 2IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. https://www.ipcc.ch/report/ar 6/wg 1/ (2021).
- 3Findlay, H. S., Feely, R. A., Jiang, L., Pelletier, G. & Bednaršek, N. Ocean acidification: another planetary boundary crossed. Glob Change Biol 31, e 70238 (2025).10.1111/gcb.70238 PMC 1214706440485607 · doi ↗ · pubmed ↗
- 4Smith, J. N. et al. Ocean acidification reduces demersal zooplankton that reside in tropical coral reefs. Nat. Clim. Change 6, 1124–1129 (2016).
- 5Randall, C. J. et al. Larval precompetency and settlement behaviour in 25 Indo-Pacific coral species. Commun. Biol.7, 142 (2024).10.1038/s 42003-024-05824-3PMC 1083050938297134 · doi ↗ · pubmed ↗
- 6Guo, W. et al. Ocean acidification has impacted coral growth on the great barrier reef. Geophys. Res. Lett.47, e 2019 GL 086761 (2020).
- 7Dickson, A. G. Guide to Best Practices for Ocean CO 2Measurements. https://www.ncei.noaa.gov/access/ocean-carbon-acidification-data-system/oceans/Handbook_2007/Guide_all_in_one.pdf (2007).
- 8Lewis, E. & Wallace, D. Program Developed for CO 2System Calculations. https://www.ncei.noaa.gov/access/ocean-carbon-acidification-data-system/oceans/CO 2SYS/co 2rprt.html (1998).