Rickettsia spp. in Finnish Ixodid ticks
Hanna Vauhkonen, Fathiah Zakham, Liina Voutilainen, Ronn Keinänen, Katja Lind, Thanakorn Niamsap, Petteri T. Puonti, Robert S. Castrén, Teemu Smura, Ruut Joensuu, Simo Nikkari, Elina Tonteri, Olli Vapalahti, Essi M. Korhonen, Anu Jääskeläinen, Anne J. Jääskeläinen, Tarja Sironen

TL;DR
This study examines the presence of Rickettsia bacteria in ticks across Finland, finding regional differences and highlighting the need for better diagnosis and monitoring of tick-borne diseases.
Contribution
The study provides new insights into the distribution and prevalence of Rickettsia species in Finnish ticks and their potential impact on human health.
Findings
Rickettsia DNA was more frequently detected in ticks from regions dominated by Ixodes ricinus compared to Ixodes persulcatus.
Rickettsia helvetica was the primary species identified, with Candidatus Rickettsia tarasevichiae found only in I. persulcatus.
A small percentage of patients suspected of tick-borne encephalitis showed Rickettsial antibody reactivity, but not at confirmed infection levels.
Abstract
Rickettsia spp. are intracellular, arthropod-borne bacteria pathogenic to humans and animals. The clinical manifestations of rickettsial infections range from mild to severe, but diagnosis is sometimes missed owing to mild symptoms or empirical antibiotic treatment for suspected tick-borne diseases. This study aimed to determine the prevalence of Rickettsia spp. in ticks across Finland, explore possible regional variations, identify the infecting Rickettsia species, and assess rickettsial exposure of certain human patients. We analysed 5101 ticks from 20 locations, collected by flagging, crowdsourcing, or removal from pet cats and dogs between 2003 and 2021 in three study sets. Tick species were determined by quantitative PCR (qPCR), Sanger sequencing, or morphology. Rickettsial prevalences were determined by qPCR, with species confirmed by Sanger sequencing. Indirect…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
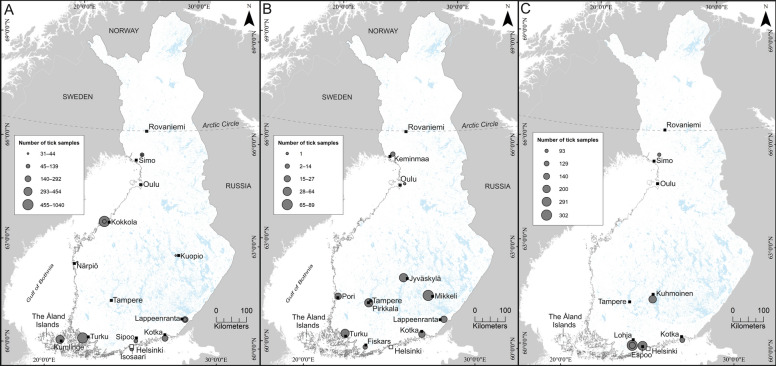 Figure 1
Figure 1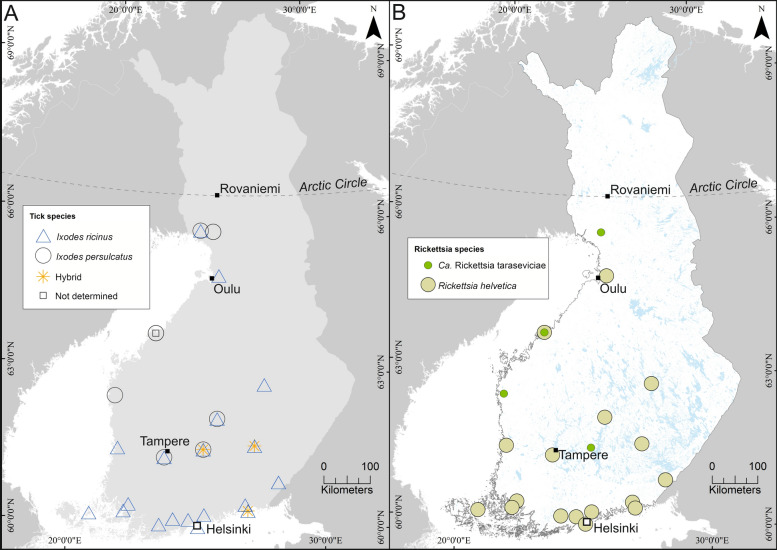 Figure 2
Figure 2 Figure 3
Figure 3- —https://doi.org/10.13039/501100004012Jane ja Aatos Erkon Säätiö
- —Veclimit Academy of Finland
- —Sakari Alhopuro Foundation
- —University of Helsinki (including Helsinki University Central Hospital)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Bartonella species infections research · Yersinia bacterium, plague, ectoparasites research
Rickettsia spp. are intracellular arthropod-borne bacteria, many of which are pathogenic to humans as well as to wild and domestic animals. While certain rickettsial species are non-pathogenic arthropod endosymbionts [1, 2], a growing number of pathogenic species have been described worldwide [3]. The clinical severities of rickettsial infections vary from asymptomatic or mild, self-limiting disease to life-threatening conditions [4]. The general symptoms include fever, malaise, lymphadenopathy and rash, and, sometimes, almost pathognomonic black eschar. If diagnosed early enough, even severe cases can be treated by tetracycline antibiotics. However, misdiagnosis is possible in mild cases where patients do not seek medical care, and, as tetracycline antibiotics are potentially used when suspecting tick-borne infections, undiagnosed rickettsial infections are treated as well [5].
Traditionally, the genus Rickettsia has been categorized into two main groups on the basis of serological pattern, the Typhus group (TG) and the Spotted fever group (SFG), which have been named after the characteristic human disease symptoms. Scrub typhus, caused by Orientia spp., has been grouped occasionally under rickettsial diseases as well [6]. However, a more detailed grouping, based on whole-genome sequencing and phylogenetics, has been suggested recently, splitting SFG into two distinct groups, SFGI and SFGII, while one SFG Rickettsia, R. helvetica, as a separate clade sharing features with other rickettsial groups [7]. The SFGI rickettsiae and R. helvetica are transmitted by ticks (occasionally by mites), whereas the other pathogenic Rickettsia groups (TG and SFGII) are typically transmitted by arthropods other than ticks, i.e. lice, mites, and fleas [7]. TG rickettsiae are considered to relate to regions with lower hygiene and European cases have mainly been detected in returning travellers [8]. TG rickettsiae, e.g., louse-born R. prowazekii, are also recognized as biothreats with history and potential for deliberate use [9]. The SFGI group has been constantly expanding due to the newly described species [7].
Specific tick species serve as vectors for particular species of SFG rickettsiae, therefore, specific rickettsial diseases can be considered endemic within the distribution range of vector species [3]. However, climate change-driven expansion of tick species to new areas may result in rickettsial disease emergence in unforeseen regions [10]. The main tick species in Finland, Ixodes ricinus (sheep/castor bean tick) and Ixodes persulcatus (taiga tick), have been found to carry R. helvetica, R. monacensis, and Candidatus R. tarasevichiae [11]. The clinical manifestations of R. helvetica and R. monacensis infections are usually mild and patients do not need medical care, however, severe conditions cannot be excluded [12, 13]. The pathogenicity of Ca. R. tarasevichiae is still unclear. It has been considered an emerging pathogen as it has been associated with clinical disease in China [14].
The laboratory diagnosis of Rickettsia spp. infections is based on either serology or molecular (mainly PCR-based) methods [15]. Molecular methods detect the presence of DNA of the infectious agent and are mainly restricted to the acute phase, whereas serology detects the immune response in the later phases of the disease [3, 15]. Additionally, sample material plays a role. Certain rickettsial species produce eschars at the site of the arthropod bite, and the pathogen may be detected in a tissue biopsy or swab by PCR or in biopsy by immunohistochemistry. By contrast, blood samples are seldom useful, as they are rarely PCR-positive even in the acute phase, thus challenging rapid diagnostics [15]. In many Nordic countries, i.e., Sweden, Norway, and the Åland islands, Finland, R. helvetica seropositivity has been detected in individuals with proven tick bites [16, 17] or individuals living in tick-dense areas [18]. In Europe, Rickettsia spp. have been detected occasionally in clinically ill dogs, and serological testing was found to outperform molecular (PCR) methods and reveal undiagnosed infections [19, 20]. Whether or not dogs show signs of infection, they can be used as sentinels of rickettsial and other tick-borne infections [21]. Although sporadic rickettsioses have been detected in returning Finnish travellers [22], to the best of our knowledge, comprehensive data on human or animal clinical cases and seroprevalence rates are still lacking in continental Finland.
Previous studies on Finnish ticks have estimated an overall Rickettsia spp. prevalence of 13.9% in I. ricinus and 6.5% in I. persulcatus [11]. Rickettsia helvetica and sporadically R. monacensis have been found in both tick species and Ca. R. tarasevichiae in I. persulcatus only [11]. The Rickettsia spp. prevalences were reported in a country-wide manner, addressing three large collection areas (I. ricinus-dominated southern Finland, 12.6%; sympatric central Finland with both species present, 10.7%; and I. persulcatus-dominated northern Finland, 6.0%). However, studies in Estonia, which resides near Finland and shares the same tick and rickettsial species, have revealed location-specific differences in prevalences [23, 24]. For example, ticks collected from rodents in Pärnumaa were more often (19.3%) infected than ticks collected from rodents in other sites (2.6–8.1%, Saaremaa to Lääne-Virumaa, respectively) [25]. Therefore, we aimed to reveal whether similar focal differences are present in Finland.
To assess the occurrence of Rickettsia spp. and reveal possible focal differences in Finland, we analyzed a total of 5101 Ixodid ticks, collected from 20 locations between 2003 and 2021, in three separate study sets (Fig. 1, Table 1). Briefly, ticks in Set 1 (n = 3472) were collected during 2003–2015 by flagging in 12 separate locations and were mainly analysed in pools. In Set 2, ticks (n = 342, removed from pet cats and dogs) were obtained during 2020–2021 by ten veterinary clinics throughout Finland [26]. Ticks in Set 3 (n = 1155) were collected during 2017–2021 from four separate locations by flagging (n = 864) or crowdsourcing (n = 291), and in a subset of the collections, the nymphs were analyzed in pools. All ticks were nymphs or adults, except four larvae in Set 1. The pooling strategies and collection sites of questing ticks in Sets 1 and 3 were originally targeted to monitor TBEV presence. The pool sizes varied between 1 and 30 per pool depending on the prevalence of the target pathogens, the specific aims of the given sampling, and the resources available for each study. The corresponding laboratory procedures were carried out during 2015–2017, 2020–2021, and 2021–2022. Figure 1 illustrates the collection areas and approximated tick amounts included in each sample set.Fig. 1. Tick collection sites in the three sample sets with approximated sample sizes. Panel A indicates the collection sites of Set 1. Panel B indicates the locations of veterinary clinics collecting ticks from pets included in Set 2. Panel C indicates the collection sites of Set 3. Maps were generated in ArcMap (version 10.8, Esri, Redlands, CA, USA)Table 1. Sample collections and results for set 1, set 2, and set 3Set 1 area, yearN tested (N pools)Tick species^a^N positive (species)Occurrence (%) (95% CI)/Minimum occurrence (%)^b^N gltA sequences^c^ (species)Seq success (%)Rickettsial speciesIsosaari Island, 200596 (11)96r/-/-5 (r)5.25 (r)100R. helveticaKokkola archipelago, 2003139 (15)Not determined21.42100R. helveticaKokkola archipelago, 2004980 (102)-/980p/-3 (p)0.22 (p)66.7Ca. R. tarasevichiaeKotka archipelago, 2011194 (70)194r/-/-14 (r)7.212 (r)85.7R. helveticaKumlinge Island, 2003454 (46)454r/-/-2 (r)0.52 (r)100R. helveticaKuopio, 201044 (5)44r/-/-2 (r)4.52 (r)100R. helveticaLappeenranta, 2005292 (29)292r/-/-23 (r)7.916 (r)69.6R. helveticaLappeenranta, 2010101 (11)101r/-/-7 (r)6.97 (r)100R. helveticaNärpiö, 200836-/36p/-1 (p)2.7 (0.49–14.2)1 (p)100Ca. R. tarasevichiaeSimo, 200998 (51)-/98p/-00–––Simo, 201531-/31p/-00 (0.00–11.0)–––Sipoo + Karhusaari Island, 201399 (48)99r/-/-15 (r)15.213 (r)86.7R. helveticaTurku archipelago, 20071040 (315)1040r/-/-9 (r)0.99 (p)100R. helveticaTotal Set 136042320r/1145p83 (77r/4p)2.371 (68r/1p)86.6Set 2 Vet practice location, year(s)N testedTick speciesN positive (species)Occurrence (%) (95% CI)N gltA sequences (species)Seq success (%)Rickettsial speciesFiskars, 2020–20211414r/-/-2 (r)14.3 (4.0–39.9)00–Jyväskylä, 2020–20214438r/5p/-^d^29 (24r/5p)65.9 (51.1–78.1)16 (15r/1p)66.7R. helveticaKeminmaa, 2021111r/10p/-00 (0.0–25.9)–––Kotka, 20202525r/-/-20 (r)80.0 (60.9–91.1)8 (r)40.0R. helveticaMikkeli, 2020–20218988r/-/1 h43 (r)48.3 (38.2–58.6)27 (r)62.8R. helveticaLappeenranta, 20202323r/-/-11 (r)47.8 (29.2–67.04)6 (r)54.5R. helveticaOulu, 202011r/-/-1 (r)^e^1 (r)100.0R. helveticaPirkkala, 2020–20216429r/35p/-14 (11r/3p)21.9 (13.5–33.4)7 (5r/2p)50.0R. helveticaPori, 2020–20212727r/-/-17 (r)63.0 (44.23–78.5)15 (r)88.2R. helveticaTurku, 2020–20214444/-/-3 (r)6.8 (2.4–18.2)00Total Set 2342289r/51p/1 h140 (132r/8p)40.9 (35.9–46.2)80 (77r/3p)58.8Set 3 area, yearN tested (N pools)Tick speciesN positive (species)Occurrence (%) (95% CI)/minimum occurrence (%)N gltA sequences (species)Seq success (%)Rickettsial speciesEspoo, 2017^f^291291r/-/-16 (r)5.5 (3.4–8.7)8 (r)50.0R. helveticaKotka Kuutsalo Island, 2018129 (102)128r/-/1 h17 (r)13.29 (r)75.0R. helveticaKuhmoinen, 202020018r/173p/9 h3 (1r/2p)1.5 (0.5–4.3)2 (p)66.7Ca. R. tarasevichiaeLohja, 2019140 (64)140r/-/-17 (r)12.112 (r)70.6R. helveticaLohja, 2021302 (64)302r/-/-22 (r)7.322 (r)100R. helveticaSimo, 202093-/93p/-1 (p)1.1 (0.2–5.8)1 (p)100Ca. R. tarasevichiaeTotal Set 31155879r/266p/10 h76 (73r/3p)6.654 (51r/3p)71.1Total Sets 1–35101299 (282r/15p)5.8205 (196r/7p)68.8Tick collection sites, number of samples (and pool numbers in Sets 1 and 3) tested, Ixodes species, number and tick species of Rickettsia spp. –positive samples, occurrence with 95% confidence intervals, number of obtained gltA. sequences, sequencing success, and rickettsial species obtained by sequencing^a^Tick species: r = I. ricinus, p = I. persulcatus, h = I. ricinus-persulcatus hybrid, species determination methods listed in Additional File Table S1^b^Minimum prevalence: assuming only one sample in a pool is positive^c^N gltA sequences; number of gltA (citrate synthase gene) sequences obtained by sequencing^d^One Jyväskylä tick identified as Rhiphicephalus sanguineus, Rickettsia spp. negative^e^Tick number too low for reliable calculation^f^Sample set obtained by crowdsourcing
Slightly different molecular methods were used for tick species identification, rickettsial occurrence, and species determination in each set. Briefly, DNA was extracted from tick homogenates by commercial kits: either TriPure Reagent DNA method, AllPrep DNA/RNA kit (Qiagen, Hilden, Germany), GenJET Genomic DNA purification kit (Thermo Fisher Scientific, Waltham, MA, USA), or DNeasy Blood and Tissue kit (Qiagen). Tick species was determined by either morphology or analysis of internal transcribed spacer 2 (ITS2) gene or mitochondrial 16S RNA gene. The presence of rickettsial DNA and the corresponding species were determined by quantitative genus-specific PCR and Sanger sequencing of citrate synthase (gltA) gene, respectively. Detailed sampling information, life stages, pool sizes, DNA extraction methods and the molecular methods in tick species and Rickettsia analyses are described in Additional File 1: Text S1, Tables S1, and S2. Subsets of the samples have been previously screened for other pathogens [26–32], described in Additional File 1: Table S1.
We found I. ricinus to prevail in southern Finland and I. persulcatus in northern Finland, and sympatric occurrence of both species in the central part of southern Finland: in Pirkkala and Jyväskylä (Set 2, previously reported by Zakham et al. [26]), and in Kuhmoinen (Set 3). Table 1 provides a detailed description and Fig. 2 a compilation of the results of this study. Our species distribution data are in line with previous reports by Laaksonen et al. [11] and are supported also by habitat suitability predictions by Uusitalo et al. [33]. Intriguingly, 4.5% of the field-collected ticks from Kuhmoinen (Set 3) were identified as I. ricinus–persulcatus first-generation hybrids using duplex real-time quantitative PCR [34], modified by Zakham et al. [32]. Respective hybrids have previously been described in regions co-habited by the two species, e.g., Estonia [35], Lempäälä, and Tampere [26, 36]. One hybrid was found in Kotka (Set 2), and one Rhipicephalus sanguineus in Jyväskylä (Set 2). The R. sanguineus was collected from a traveling dog returning from Spain [26]. The tick was not infected with Rickettsia.Fig. 2. Presence of tick and Rickettsia spp. in the collection sites. Panel A shows the presence of Ixodes ricinus (red), Ixodes persulcatus (blue) and I. ricinus-persulcatus hybrids (pink) in the collection sites. Collection sites where tick species detection was based on methods other than molecular methods are indicated by a small grey box symbol. Panel B shows Rickettsia species (R. helvetica light green, Ca. R. tarasevichiae, dark green) in the respective collection sites. Maps were generated in ArcMap (version 10.8, Esri, Redlands, CA, USA)
Overall, in Sets 2 and 3, Rickettsia spp. infections were more common in I. ricinus (132/289, 45.7% and 73/879, 8.3%, respectively) than in I. persulcatus (8/51, 15.7% and 3/266, 1.1%) (p < 0.001, 2-sample z-test). The 6.6% overall occurrence obtained from Set 3 is in line with previous observations [11], whereas the considerably higher (40.9%) occurrence in ticks collected from pets (Set 2) was unexpected and could not be explained by differences in laboratory procedures or sample origin. Indeed, the crowdsourced samples used by Laaksonen et al. [11] were mainly obtained from companion animals during 2015 and probably are geographically biased toward residential areas [37]. However, while the specific locations of ticks collected from pets cannot be verified because of scant traveling details provided by owners, crowdsourcing is still considered a valid method of monitoring pathogen occurrences in larger areas [11, 37]. In total, 27 of the veterinary clinic pets included in our study had more than one tick, and only 3 pets had two engorged rickettsia-positive ticks, suggesting a minor role for infection by co-feeding. Since most of the samples in Set 1 were analyzed in pools of varying sizes between and within collection sites, occurrence could be approximated only to the lowest percentage i.e., assuming only one positive sample in the pool, resulting in a total minimum occurrence of 2.3% (83/3604).
At locations where the only tick species found was I. ricinus, the rickettsial occurrences varied within and between sample sets (0–80.0%), with Set 2 having the highest occurrences. Southwestern Finland and its archipelago had the lowest occurrences (0.5–6.8%). Accordingly, Southwestern Finland and the Åland islands have previously been described to present lower rickettsial occurrences (0–5.1%) than the national average (10.8%) [16, 34]. At locations where the only tick species found was I. persulcatus, the occurrences were lower (0–2.7%), as also reported previously [11]. The only tick sample from Oulu, Northern Finland (Set 2), was Rickettsia spp*.* –positive I. ricinus detached from a dog and was not considered to represent the whole area, as it could have originated from a migratory bird or a traveling dog.
Among the three sympatric locations, Kuhmoinen was dominated by I. persulcatus (173/200, 86.5%), whereas in Pirkkala I. persulcatus was nearly equally abundant (35/64, 54.7%) as I. ricinus. In Jyväskylä, I. persulcatus was present only as a minority (5/44, 11.4%). The overall rickettsial occurrence was 1.5% in Kuhmoinen (Set 3, two I. persulcatus and one I. ricinus). In Jyväskylä and Pirkkala (Set 2), overall Rickettsia spp. occurrences of 65.9% (29/44) and 21.9% (14/64) were observed, respectively. All (5/5) I. persulcatus ticks and 63.2% (24/38) of I. ricinus in Jyväskylä and 8.6% (3/35) of I. persulcatus and 37.9% (11/29) I. ricinus in Pirkkala were Rickettsia spp. positive. The nine I. ricinus—persulcatus hybrids of Kuhmoinen and the one of Kotka were *Rickettsia-*negative.
The main rickettsial species observed by sequencing was R. helvetica, and it was most often found in I. ricinus-dominated regions. Six Ca. R. tarasevichiae-positive samples or pools were found in I. persulcatus ticks (Sets 1 and 3) in Kuhmoinen, Simo, Närpiö, and Kokkola 2004. Only R. helvetica was found in Set 2, including the five sequenceable I. persulcatus samples in Jyväskylä and Pirkkala. No R. monacensis or any other rickettsial species were found in the entire study. The sequencing success rate ranged from 40 to 100% between the sample sets, Set 2 with the lowest and Set 3 with the highest success rate (Table 1). Variation in DNA extraction methods and gltA sequencing strategies (un-nested versus nested) between the study sets may explain the differences in success rates. Although the nested PCR strategy in Sanger sequencing has been suggested as more sensitive, the better sequencing success observed in Set 1 with an un-nested strategy could be explained by pooled samples. In addition, samples with qPCR Ct values above 35 failed more often to produce sequence, reflecting either lower rickettsial proportion or lower DNA quality of the sample. Previously, Laaksonen et al. [11] reported a sequencing success rate of 68.1%.
We found no obvious reason for the substantially higher occurrences in our collection of ticks detached from pets (Set 2). Six engorged rickettsia-positive ticks were collected from three pets, two ticks from each, suggesting the possibility of infection by co-feeding or acquiring rickettsial infection from the host. However, different collection years or seasonal variation affecting the habitat might explain the differences [37–39]. As different nutritional or climatic factors have been suggested to affect the tick microbiome [40, 41], rickettsial abundances within the tick host may also depend on environmental conditions, including weather. In addition, competition between pathogenic and nonpathogenic Rickettsia species has been observed within the vector cell, e.g., a nonpathogenic endosymbiont may hinder the infection of pathogenic species [42], therefore explaining patchy occurrences. For more accurate comparison of the occurrence of pathogens in ticks collected from pets with those collected from the environment, environmental sampling close to the veterinary clinics collecting ticks from pets and around the same time is needed. This will be considered in future studies. To our knowledge, no seroprevalence data on Rickettsia spp. in companion animals living in Finland exist, and thus, their actual exposure is unknown. Antibodies showing high exposure to SFG rickettsiae have been reported from Sweden and Germany [43, 44].
To estimate the prevalence of human rickettsial infections in Finland, sera from 226 individual patients suspected of tick-borne encephalitis (TBE) were obtained from late May to early July 2018 throughout the country. The samples were screened for rickettsial antibodies using commercial immunofluorescence assay (IFA) to detect possible undiagnosed rickettsioses. Sample and IFA analysis details are provided in Additional File 2: Text S2. Overall, 13 sera (5.8%) were considered borderline or low positive, and these might be due to an unspecific reaction. There were no high positive findings using commercial IFA, and no infection caused by Rickettsia spp. was detected (Additional File 1: Table S3). As no data on potential eschars or traveling and no paired sample sera were available for borderline positive patients, no conclusions could be made regarding endemic, travel-related, or past or present infections. For comparison, in Denmark, 2147 individuals were tested for rickettsial antibodies during 2008–2015, yielding 561 positives (26%; cut-off 1:512), 86% of which were SFG-positive [45].
In conclusion, we describe Rickettsia spp. occurrence in 20 geographical locations in Finland Ticks collected from pets appeared to be more frequently Rickettsia-positive than those collected by other methods, and I. ricinus ticks seem to be more commonly infected with rickettsiae than I. persulcatus ticks. The reason for regional differences in rickettsial occurrence—whether attributable to the predominance of specific tick species, climatic or nutritional factors, interactions with competing endosymbiotic organisms, or other yet unidentified determinants—remains to be elucidated. The variable sampling strategies presented in this study limit the assessment of these determinants. We found no clear rickettsial seropositivity in patients suspected of TBE; however, for verified serology-based diagnosis, paired serum samples showing that rising titres are needed. In any case, our findings underscore the common occurrence of Rickettsia spp*.* in ticks, and hence, the potential for rickettsial diseases, necessitating enhanced diagnostic frameworks to address disease burden and improve disease surveillance. Since potentially pathogenic rickettsial species clearly exist in Ixodid ticks in Finland, rickettsioses need to be included among differential diagnoses by both physicians and veterinarians.
Supplementary Information
Additional file 1.Additional file 2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hodosi R, Kazimirova M, Soltys K. What do we know about the microbiome of I. ricinus? Front Cell Infect Microbiol. 2022;12:990889. 10.3389/fcimb.2022.990889.
- 2Abdad MY, Abou Abdallah R, Fournier P-E, Stenos J, Vasoo S. A Concise Review of the Epidemiologyand Diagnostics of Rickettsioses: Rickettsia and Orientia spp. J Clin Microbiol. 2018;56:e 01728-17. 10.1128/jcm.01728-17.
- 3Laaksonen M, Klemola T, Feuth E, Sormunen JJ, Puisto A, Mäkelä S, et al. Tick-borne pathogens in Finland: comparison of Ixodes ricinus and I. persulcatus in sympatric and parapatric areas. Parasit Vectors. 2018;11:556. 10.1186/s 13071-018-3131-y.
- 4Lindblom A, Wallménius K, Sjöwall J, Fryland L, Wilhelmsson P, Lindgren P-E, et al. Prevalenceof Rickettsia spp. in ticks and serological and clinical outcomes in tick-bitten individuals in Sweden andon the Åland islands. P Lo S One. 2016;11:e 0166653. 10.1371/journal.pone.0166653.
- 5Solano-Gallego L, Caprì A, Pennisi MG, Caldin M, Furlanello T, Trotta M. Acute febrile illness isassociated with Rickettsia spp. infection in dogs. Parasit Vectors. 2015;8:216. 10.1186/s 13071-015-0824-3.
- 6Vikentjeva M, Geller J, Remm J, Golovljova I. Rickettsia spp. in rodent-attached ticks in Estonia andfi rst evidence of spotted fever group Rickettsia species Candidatus Rickettsia uralica in Europe. Parasit Vectors. 2021;14:65. 10.1186/s 13071-020-04564-7.
- 7Uusitalo R, Siljander M, Lindén A, Sormunen JJ, Aalto J, Hendrickx G, et al. Predicting habitatsuitability for Ixodes ricinus and Ixodes persulcatusticks in Finland. Parasit Vectors. 2022;15:310. 10.1186/s 13071-022-05410-8.
- 8Militzer N, Pinecki Socias S, Nijhof AM. Changes in the Ixodes ricinus microbiome associated withartifi cial tick feeding. Front Microbiol. 2023;13:1050063. 10.3389/fmicb.2022.1050063.