Growth/Differentiation Factor 15 Promotes a Pro‐Regenerative Response in Chondrocytes Upon Cartilage Injury
Sara Sofi Marques, Alexandra Liebaug, Svenja Maurer, Dietrich Rothenbacher, Rolf E. Brenner, Jana Riegger

TL;DR
This study shows that GDF-15, a stress-related protein, increases after cartilage injury and may help protect and regenerate cartilage in posttraumatic osteoarthritis.
Contribution
The study first identifies GDF-15 as a senescence-associated yet pro-regenerative cytokine in human posttraumatic osteoarthritis.
Findings
GDF-15 levels are significantly higher in synovial fluid of PTOA patients compared to IOA patients.
GDF-15 is produced by chondrocytes after cartilage injury and is mediated by oxidative stress and p53 activation.
GDF-15 promotes chondrocyte proliferation and cell protection after cartilage trauma.
Abstract
Posttraumatic osteoarthritis (PTOA) is a special form of osteoarthritis (OA), developing after joint injuries. Except for some minor clinical differences, no biologic marker has yet been identified to distinguish idiopathic OA (IOA) from PTOA. In this study, we investigated the expression of the stress‐responsive cytokine growth differentiation factor 15 (GDF‐15) in clinical samples from the Ulm OA study cohort and in a human ex vivo cartilage trauma model. GDF‐15 levels were significantly higher in synovial fluid of PTOA patients as compared to IOA patients. We confirmed that fibroblast‐like synoviocytes secreted GDF‐15 after stimulation with medium of ex vivo‐traumatized cartilage. Moreover, GDF‐15 and its receptor, GFRAL, were elevated in highly degenerated OA cartilage. By means of a human cartilage trauma model, we discovered that chondrocytes produced GDF‐15 upon tissue injury,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
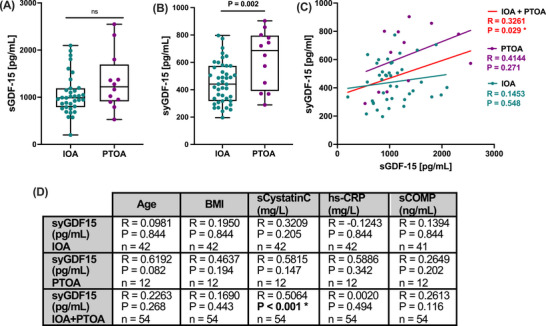 FIGURE 1
FIGURE 1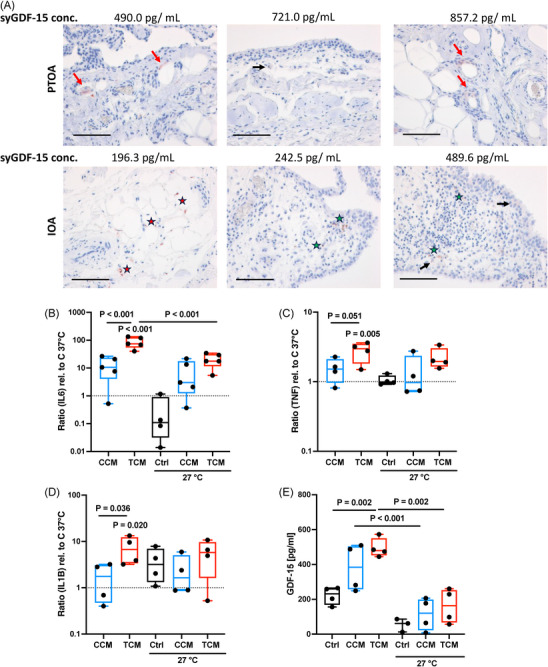 FIGURE 2
FIGURE 2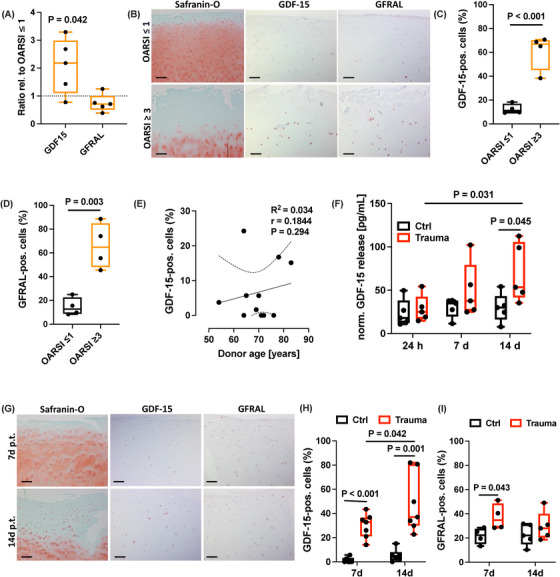 FIGURE 3
FIGURE 3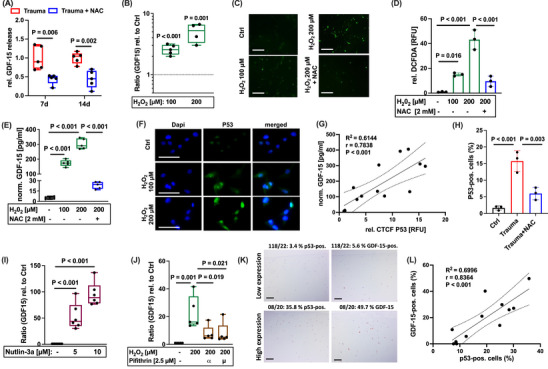 FIGURE 4
FIGURE 4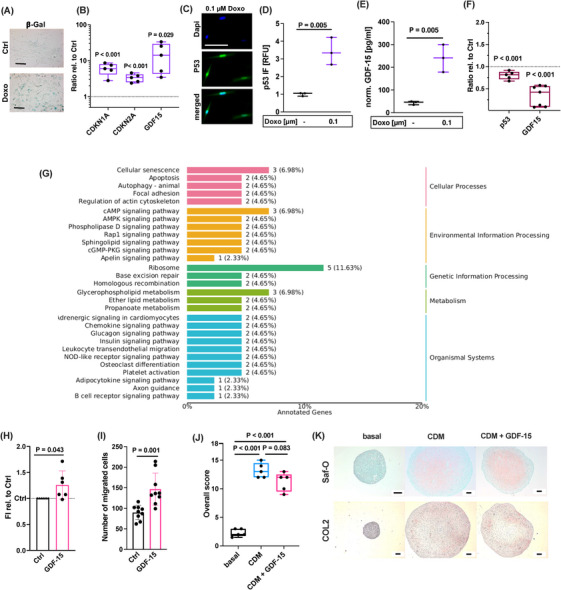 FIGURE 5
FIGURE 5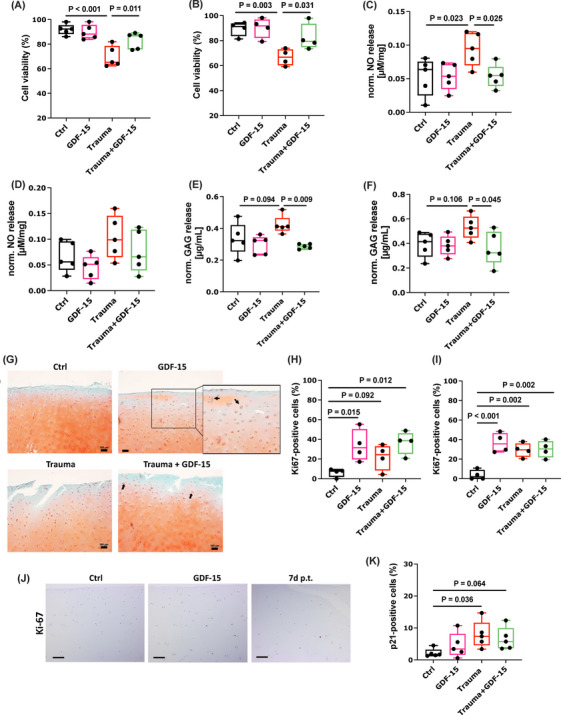 FIGURE 6
FIGURE 6| Parameter | Idiopathic OA ( | Posttraumatic OA ( | Total ( |
|
|---|---|---|---|---|
|
| 66.3 ± 5.22 | 66.5 ± 5.92 | 66.4 ± 5.32 |
0.873 |
|
|
Male: 13 (31.0 %) Female: 29 (69.0 %) |
Male: 2 (16.7 %) Female: 10 (83.3 %) |
Male: 15 (27.8 %) Female: 39 (72.2 %) | 0.260 |
|
| 29.8 ± 4.43 | 28.9 ± 5.46 | 29.6 ± 4.62 | 0.603 |
|
| 6 (14.3 %) | 2 (16.7 %) | 8 (14.8 %) | 0.695 |
- —European Social Fund10.13039/501100004895
- —Ministry of Science
- —Research and Arts Baden‐Württemberg
- —University of Ulm10.13039/501100008977
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGDF15 and Related Biomarkers · Osteoarthritis Treatment and Mechanisms · Connective Tissue Growth Factor Research
Introduction
1
Osteoarthritis (OA) is commonly considered as the most abundant joint disease and major cause of disability in elderly people. Most cases of OA develop without a certain cause, but seem to be associated with gender, age, and genetic predisposition. This form of OA is also referred to as primary or idiopathic OA (IOA). In contrast to IOA, posttraumatic OA (PTOA) results from a certain event, namely a preceding joint injury, such as meniscal or ligament rupture, chondral defects, and articular fracture [1]. Although there are some epidemiological differences between IOA and PTOA, for example the earlier onset of PTOA, it remains largely unclear whether it is possible to distinguish the pathogenesis of the different forms on molecular or biochemical level [2]. PTOA accounts for about 12 % of all OA cases and represents a particularly severe burden due to the significantly younger age of the affected patients, which is accompanied by early invalidity and thus inability to work, as well as enhanced lifetime risk for revision arthroplasty [3]. Not all patients, receiving a joint injury, are deemed to develop a PTOA in later life, because further factors are thought to be involved in disease progression [4]. Identification of novel biomarkers serving as prognostic tools to estimate the actual risk of PTOA development after injury or as therapeutic target structures is of high priority. Such biologic indicators are a fundamental requirement to provide a tailored therapy (e.g., additional physiotherapy or pharmacologic treatment) for high‐risk patients, which may rapidly develop PTOA after joint injury.
Recently, we compared the clinical parameters of patients suffering from knee IOA or PTOA among the Ulm Osteoarthritis Study cohort. The study revealed differences between IOA and PTOA patients regarding the WOMAC pain subscale and general physical function at baseline (before knee joint replacement), as well as serum high‐sensitivity cardiac troponin T levels and mortality (manuscript in preparation). In the present study, we focused on growth differentiation factor 15 (GDF‐15) as a potential indicator to differentiate IOA and PTOA. The protein has previously been described as elevated in different forms of rheumatic diseases, including systemic lupus erythematosus, spondyloarthritis, and rheumatoid arthritis, in which it was associated with disease severity and activity [5, 6, 7]. GDF‐15, also referred to as the macrophage inhibitory cytokine 1, is a stress‐inducible cytokine and atypical member of the transforming growth factor‐β (TGF‐β) superfamily, which has emerged as a marker of all‐cause mortality in heart failure and cancer, among others [8, 9]. Accordingly, we previously reported that the cytokine represents a strong and independent risk factor for decreased survival in subjects undergoing unilateral total hip or knee arthroplasty due to advanced OA within the Ulm OA study cohort [10]. Despite its bad reputation as all‐cause mortality marker, increased GDF‐15 expression has also been discussed as a survival response [11]. Accordingly, GDF‐15 is produced by various cell types during acute tissue injury and was described as prominent senescence‐associated secretory phenotype (SASP) factor in senescent cells [12]. Although transient senescence is considered as beneficial in terms of tissue remodeling and repair, chronic accumulation of senescent cells in cartilage or other tissues contributes to progressive degeneration and dysfunction due to the excessive secretion of catabolic and pro‐inflammatory SASP factors [13, 14]. Considering its association with injury response and senescence, mechanisms that are obviously involved in the pathogenesis of PTOA [15], we hypothesized that GDF‐15 might be a potential mediator in trauma‐induced cartilage degeneration.
In the present study, we first confirmed significantly higher concentrations of GDF‐15 in clinical synovial fluid samples of PTOA patients as compared to IOA patients. Moreover, we found clear evidence that GDF‐15 was expressed by chondrocytes in highly degenerated human cartilage as well as after ex vivo trauma. Further investigation revealed that GDF‐15 in human articular chondrocytes (hAC) was promoted by oxidative stress and subsequent activation of p53. Strikingly, we observed that GDF‐15 secretion did not induce pathophysiologic processes but contributed to cell proliferation and protection, implying potentially pro‐regenerative effects after cartilage injury. Taken together, this is the first study on GDF‐15 in PTOA considering mainly clinical samples and providing a novel insight into the role of this stress‐inducible cytokine.
Results
2
Synovial GDF‐15 Concentrations Are Significantly Higher in PTOA as Compared to IOA Samples
2.1
To prove our hypothesis that GDF‐15 is differentially expressed in IOA and PTOA patients, we analyzed clinical serum and synovial fluid samples derived from the Ulm Osteoarthritis study cohort. Although we found increased serum levels of GDF‐15 (sGDF‐15) in the PTOA group, there was no significant difference compared to the IOA group (Figure 1A). To clarify whether GDF‐15 was locally expressed in the affected knee joints of the respective patients, we investigated the synovial GDF‐15 (syGDF‐15) concentrations. In fact, syGDF‐15 levels were significantly higher in synovial fluid samples of PTOA patients as compared to the IOA group (mean ± SD: PTOA 625.9 ng/mL ± 210.2; IOA 452.8 ng/mL ± 150.3; Figure 1B). While syGDF‐15 concentrations significantly correlated with the serum GDF‐15 levels in the overall collective, we could not find any association in the IOA or PTOA group (Figure 1C). Moreover, syGDF‐15 was not significantly associated with age, BMI, serum cystatin C, hs‐CRP, and serum COMP in separate analyses of the IOA and PTOA groups (Figure 1D). However, syGDF‐15 concentrations significantly correlated with the serum cystatin C levels in the overall collective. No significant differences in syGDF‐15 levels were observed between female and male patients (Figure S1).
Assessment of local GDF‐15 levels in synovial fluid of PTOA and IOA patients. (A) Serum GDF‐15 (sGDF‐15) and (B) synovial GDF‐15 (syGDF‐15) concentrations of IOA and PTOA patients at the time point of knee TEP implantation were quantified by means of ELISAs. (C) Corresponding correlation analysis between syGDF‐15 and sGDF‐15 in IOA and PTOA patients. (D) Overview of correlation analyses between syGDF‐15 concentrations with clinical and laboratory parameters in IOA and PTOA patients separately. sCOMP = serum COMP, sCystatinC = serum cystatin C.
Serum cystatin C concentrations were previously found to correlate with disease activity and chronic inflammation in rheumatic arthritis [16]. Thus, our findings might indicate a potential connection between GDF‐15 and local inflammatory processes in the PTOA‐affected knee.
Synovial Cells Express Enhanced Levels of GDF‐15 in Response to Trauma‐Conditioned Medium
2.2
To investigate the origin of syGDF‐15 and its potential connection to synovitis, we quantified GDF‐15 expressing cells in the synovial tissue of IOA and PTOA patients by immunostaining. As expected for IOA and PTOA, which are considered as low‐grade inflammatory forms of arthritic disease [17], the histologic assessment revealed only mild synovitis and no difference between IOA and PTOA. Overall, GDF‐15 expressing cells could be found in the synovial tissue of both groups and there was no clear association with the posttraumatic history of the patients or the syGDF‐15 concentrations. Furthermore, GDF‐15 was not co‐located to infiltrated immune cells (Figure 2A).
Expression of GDF‐15 in synovial cells. (A) Representative images of IHC staining against GDF‐15 in the synovial membrane of IOA and PTOA patients, including the respective syGDF‐15 concentrations. GDF‐15 expressing cells are highlighted as follows: synovial lining cells = black arrows; infiltrated immune/small follicle‐like lymphocytic cells = green star; endothelial cells = red arrow; adipocytes = red stars. The black bars represent 100 µm. (B) Gene expression analysis of IL6, (C) TNF, and (D) IL1B, as well as (E) release of GDF‐15 by human fibroblast‐like synoviocytes (hFLS) stimulated with cartilage‐conditioned medium (CCM) or trauma‐conditioned medium (TCM) and cultivated under normothermic (37°C) or hypothermic (27°C) conditions for 4 days. mRNA levels were normalized to control tissue at 37°C.
Previously, we reported a pro‐inflammatory response of isolated hFLS after stimulation with trauma‐conditioned medium (TCM) derived from ex vivo traumatized human cartilage explants. This activation is most likely triggered by the binding of trauma‐derived damage‐associated molecular patterns (DAMPs) to cellular pattern recognition receptors, in particularly toll‐like receptors, and was found to be partly attenuated at hypothermic conditions (27°C) [18]. We used this in vitro model to investigate whether activated hFLS express GDF‐15. While the gene expression of IL6, TNF, and IL1B was significantly enhanced in hFLS upon TCM stimulation, only that of IL6 was significantly reduced at hypothermic conditions (Figure 2B–D). In line to the other cytokines, release of GDF‐15 was significantly increased by hFLS in response to TCM. This effect was completely suppressed at 27°C (Figure 2E).
Altogether, the results imply that hFLS might secrete GDF‐15 in response to trauma‐associated DAMPS.
GDF‐15 Expression Is Significantly Enhanced in Chondrocytes of Highly Degenerated or Traumatized Cartilage
2.3
To clarify whether hAC contribute to syGDF‐15 levels and might represent possible recipient cells of the cytokine, we investigated the expression of GDF‐15 and its receptor GFRAL in macroscopically intact (OARSI ≤ 1) and highly degenerated (OARSI ≥ 3) cartilage of the same OA patients. While the gene expression of GDF15 was significantly increased in highly degenerated cartilage tissue, no difference was observed for GFRAL as compared to macroscopically intact tissue (Figure 3A). However, on protein level both GDF‐15 and GFRAL were found to be significantly elevated in highly degenerated cartilage, as determined by IHC (Figure 3B–D). This finding indicates that GDF‐15 expression is associated with cartilage degeneration and that osteoarthritic chondrocytes might be more susceptible to the cytokine due to the upregulation of GFRAL.
Expression of GDF‐15 in highly degenerated and traumatized cartilage tissue. (A) Gene expression analysis of GDF15 and its receptor GFRAL in highly degenerated cartilage (OARSI ≥ 3) relative to macroscopically intact (OARSI ≤ 1) tissue. (B) Representative images of Safranin‐O staining as well as IHC staining against GDF‐15 and GFRAL in the respective tissue. (C) Quantification of GDF‐15 and (D) GFRAL expressing cells in macroscopically intact and highly degenerated cartilage by IHC. (E) Correlation analysis between percentage of GDF‐15‐positive cells in macroscopically intact tissue and the respective donor age. (F) Release of GDF‐15 at 24 h, 7 days, and 14 days after ex vivo cartilage trauma. (G) Representative images of Safranin‐O staining as well as IHC staining against GDF‐15 and GFRAL at 7 days and 14 days after ex vivo cartilage trauma. Respective quantification of IHC staining against (H) GDF‐15 and (I) GFRAL. The black bars in (B) and (G) represent 200 µm. p.t. = post trauma.
Although syGDF‐15 concentration correlated with donor age of PTOA patients, as described above, we did not find an association between the percentage of GDF15‐expressing chondrocytes in macroscopically intact cartilage tissue and age of the respective donors (Figure 3E).
Next, we wanted to clarify the potential link between cartilage injury and GDF‐15 expression. For this purpose, human macroscopically intact cartilage was traumatized by means of an ex vivo drop tower device [19]. On gene expression level, GDF15 was induced 14 days after trauma, but not at the earlier time points (Figure S2A), which was largely consistent with the time‐dependent increase of GDF‐15 biosynthesis as confirmed by ELISA (Figure 3F) and IHC (Figure 3G, H). GFRAL expression, in turn, was elevated 7 days post trauma, but was largely restored after 14 days (Figures 3G, I and S2B).
Qualitative differences between cartilage tissue considered as OARSI ≤ 1 and OARSI ≥ 3 or trauma‐related changes in cellularity and glycosaminoglycan content are demonstrated by means of exemplary Safranin‐O staining (Figure 3B, G).
Overall, these results imply that GDF‐15 expression is associated with cartilage injury and might occur as a delayed—not immediate—response.
Expression of GDF‐15 Is Regulated by Stress‐Induced p53 Activity
2.4
Oxidative stress is a crucial driver of various pathomechanisms during OA progression, including catabolic and pro‐inflammatory processes, regulated cell death, and senescence [14, 15, 19, 20]. To clarify whether ROS represent a potential trigger of GDF‐15 expression, we investigated its expression after induction of oxidative stress in chondrocytes.
We previously reported about the harmful consequences of enhanced oxidative stress after cartilage trauma in a human ex vivo and rabbit in vivo model, which could be attenuated by antioxidative therapy using N‐acetyl cysteine (NAC) [19, 21, 22]. In accordance with our assumption, addition of NAC significantly reduced the secretion of GDF‐15 after ex vivo cartilage trauma as compared to the untreated impacted tissue (Figure 4A). Moreover, the gene expression and secretion of GDF‐15 was significantly increased upon exposure of isolated hAC to H_2_O_2_ and consequent intracellular ROS accumulation (Figure 4B–E). Intracellular ROS levels and stress‐induced secretion of GDF‐15 was almost completely abolished by addition of the antioxidant NAC (Figure 4C–E). Taken together, these data imply that oxidative stress and subsequent cellular damage induce GDF‐15 expression in hAC.
GDF‐15 expression is modulated by ROS‐induced p53 activation. (A) Relative release of GDF‐15 at 7 days and 14 days after ex vivo cartilage trauma of untreated or NAC‐treated explants. (B) Gene expression of GDF15 and (C, D) detection of intracellular ROS levels by DCFDA assay in isolated hAC upon H2O2 exposure for 48 h. (E) Release of GDF‐15 by H2O2‐treated hAC with or without treatment of NAC. (F) Representative images of IF staining of p53 in H2O2‐treated hAC. (G) Correlation analysis between CTCF values of p53 staining and GDF‐15 concentrations of unstimulated or H2O2‐treated hAC. (H) Quantification of posttraumatic p53 activation (detected by means of IHC) and effects of antioxidative therapy by NAC at 7 days after ex vivo cartilage trauma. Gene expression analysis of GDF‐15 in hAC after stimulation with (I) the MDM2 antagonist (p53 inducing) Nutlin‐3a and (J) the inhibitor of p53 transcription activity, Pifithrin‐α or Pifithrin‐μ. (K) Representative images of IHC staining against p53 and GDF‐15 in human cartilage and (L) corresponding correlation analysis. The white bars in (C) and (F) represent 100 µm. The black bars in (K) represent 200 µm.
Oxidative stress has various effects regarding chondrocyte fate, comprising regulated cell death and senescence [19, 20]. Moreover, the tumor suppressor protein p53 is a key regulator of ROS‐mediated cell fate decision and GDF‐15 is considered as a transcriptional target of this stress‐responsive transcription factor [14, 23]. To determine the role of p53 in GDF‐15 expression in hAC, p53 activity was assessed by immunofluorescence (IF). As expected, p53 activity was significantly enhanced after exposure to H_2_O_2_ as indicated by enhanced fluorescence and nuclear translocation (Figure 4F). Subsequent correlation analysis of the p53 fluorescence signal (CTCF) and the release of GDF‐15 in the corresponding samples revealed a positive association between the proteins (Figure 4G). Accordingly, NAC treatment significantly reduced the posttraumatic activation of p53 after ex vivo cartilage trauma (Figure 4H). Induction of p53 activity by the MDM2 antagonist Nutlin‐3a resulted in a concentration‐dependent increase of the gene expression of GDF‐15, while addition of the p53 inhibitors Pifithrin‐α or Pifithrin‐μ to H_2_O_2_‐treated hAC significantly reduced the expression of the cytokine (Figure 4I, J). The positive correlation between p53 activity and GDF‐15 expression was confirmed in human cartilage samples, comprising different OARSI grades as well as in impacted and unimpacted tissue sections (Figure 4K, L).
Taken together, GDF‐15 expression in hAC is driven by oxidative stress and subsequent activation of the stress‐inducible transcription factor p53.
GDF‐15 Is Secreted by Senescent Chondrocytes but Does Not Induce Paracrine Senescence
2.5
Long‐term exposure to non‐cytotoxic oxidative stress and subsequent cell damage is considered as major driver of cellular senescence [14]. Recently, we described a novel doxorubicin (Doxo)‐based in vitro model to reliably induce stable chondrosenescence in hAC [24]. We used this in vitro model to examine the expression of GDF‐15 as a potential SASP factor of senescent chondrocytes. Chondrocyte senescence was confirmed by ß‐galactosidase staining, the expression of CDKN1A, CDKN2A, and p53 activity (Figure 5A–D). Moreover, senescent chondrocytes expressed high levels of GDF‐15 (Figure 5E). To further confirm the association between chondrosenescence and GDF‐15 expression, high passage OA chondrocytes were treated by the senolytics dasatinib and quercetin as previously described [25]. Selective elimination of senescent chondrocytes resulted in declined expression of GDF15 and its transcription factor p53 (Figure 5F).
GDF‐15 expression by senescent hAC and response of hAC to exogenous GDF‐15. Characterization of Doxo‐stimulated hAC by (A) exemplary ß‐galactosidase (ß‐Gal) staining, (B) gene expression of CDKN1A, CDKN2A, and GDF15, and (C, D) IF of p53. (E) Release of GDF‐15 by Doxo‐stimulated hAC. (F) Gene expression of GDF15 and p53 at day 7 after selective elimination of senescent hAC by dasatinib (250 nM) and quercetin (5 µM) for 3 days. (G) GO enrichment analysis depicting classification of significantly regulated genes after rhGDF‐15 stimulation as compared to untreated control hAC. (H) Relative fluorescence intensity (FI) of the alamarBlue assay 48 h after stimulation of hAC with rhGDF‐15. (I) Number of migrated hAC 48 h after stimulation with rhGDF‐15 in a scratch assay. (J) Scoring of in vitro redifferentiation of hAC after 4 weeks in chondrogenic differentiation medium (CDM) with or without addition of rhGDF‐15. (K) Representative Safranin‐O (Saf‐O) and IHC staining against COL2 of respective redifferentiation. The black bars in (A, K) and white bar in (C) represent 100 µm.
As our findings indicated that hAC not only produce GDF‐15 but might also act as recipient cells, in particularly under pathophysiologic conditions, we investigated the response of isolated hAC to exogenous rhGDF‐15 by means of a bulk RNA sequencing. Comparison of rhGDF‐15‐stimulated hAC with untreated controls revealed that 93 genes were differentially expressed, with 40 genes being upregulated and 53 genes being downregulated (Figures S3 and S4A). By means of a Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis, the regulated genes could be classified to cellular senescence (cellular processes) and glycerophospholipid metabolism among others (Figure 5G). Further analysis by Gene Ontology (GO) enrichment revealed that processes associated with biological regulation, metabolism, signaling, response to stimulus, development, and immune system were among the top regulated biological processes in hAC upon rhGDF‐15 stimulation (Figure S4B–D). However, we could not identify any significant effect of rhGDF‐15 stimulation on the expression of genes related to SASP or cartilage biosynthesis/degradation by RNA sequencing.
As the RNA‐sequencing still indicated cellular senescence as potentially modulated process after rhGDF‐15 stimulation, we performed additional qPCR analysis, focusing on senescence‐associated target genes. Stimulation with rhGDF‐15 for 48 h reduced the gene expression of CDKN1A and CDKN2A, while having no effect on common SASP factors described in the context of OA [20, 24], such as CXCL1, IL‐6, and MMP13 (Figure S4E). In line with the reduced mRNA levels of the cell cycle inhibitors, rhGDF‐15 stimulation induced the proliferation of hAC, as determined by an alamarBlue assay (Figure 5H). Furthermore, addition of rhGDF‐15 significantly increased the number of migrated cells in a scratch assay, which indicates elevated migratory and mitotic activity (Figure 5I). Addition of lower (50 ng/mL) or higher (1000 ng/mL) concentrations of rhGDF‐15 had minor effects on proliferation and migration of nAC (Figure S5A, B). It should be noted that rhGDF‐15 had no chemoattractive effect on mesenchymal stem cells, as investigated by means of a Boyden chamber assay (Figure S6A). As proliferation and migration are commonly associated with tissue regeneration, we conducted an in vitro chondrogenesis assay on dedifferentiated hAC in presence of the cytokine. Addition of rhGDF‐15 during chondrogenic re‐differentiation of hAC decreased the overall score of neocartilage formation to some extent, but did not inhibit chondrogenesis per se (Figure 5J, K).
Overall, these data imply that GDF‐15 might serve as a SASP factor, which promotes migratory and mitotic properties in hAC, but does not accelerate chondrosenescence.
Exogenous GDF‐15 Mitigates Trauma‐Induced Pathomechanisms in Human Cartilage and Might Promote a Pro‐Regenerative Phenotype
2.6
To investigate the potential modulative effect of GDF‐15 on hAC after cartilage injury, the cytokine was added for 7 days and 14 days after ex vivo trauma. While the mechanical impact reduced the cell viability as expected, addition of rhGDF‐15 significantly prevented chondrocyte death at both time points (Figure 6A, B). Moreover, rhGDF‐15 significantly reduced trauma‐induced NO production after 7 days, whereas this effect could not be observed at the later time point (Figure 6C, D). Although trauma‐induced release of GAG was only increased by trend, treatment with rhGDF‐15 significantly attenuated the amount of the matrix components in culture media of impacted cartilage explants (Figure 6E, F). Exemplary Safranin‐O staining of human cartilage explants confirmed higher proteoglycan content in rhGDF‐15‐treated explants after ex vivo trauma and indicated enhanced cell cluster formation, regardless of a preceding impact (Figure 6G).
Effects of exogenous rhGDF‐15 after ex vivo cartilage trauma. Cell viability of hAC at (A) 7 days and (B) 14 days after ex vivo cartilage trauma and/or rhGDF‐15 stimulation, determined by a live/dead assay. Normalized release of NO at (C) 7 days and (D) 14 days after ex vivo cartilage trauma and/or rhGDF‐15 stimulation, determined by a Griess assay. Normalized GAG release at (E) 7 days and (F) 14 days after ex vivo cartilage trauma and/or rhGDF‐15 stimulation, determined by a DMMB assay. (G) Representative Saf‐O staining of human cartilage explants 7 days after ex vivo trauma and/or rhGDF‐15 stimulation. Percentage of Ki67‐positive cells at (H) 7 days and (I) 14 days after ex vivo cartilage trauma and/or rhGDF‐15 stimulation. (J) Representative images of IHC staining against Ki‐67 7 days after rhGDF‐15 stimulation or ex vivo cartilage trauma. (K) Percentage of p21‐positive cells at day 14 after ex vivo cartilage trauma and/or rhGDF‐15 stimulation. The black bars in (G) represent 100 µm and in (J) represent 200 µm. p.t. = post trauma.
To clarify potential pro‐mitotic activity of chondrocytes upon rhGDF‐15 stimulation, a Ki67 staining was performed. In line with the cluster formation described above, the percentage of Ki67‐positive chondrocytes was significantly increased upon addition of rhGDF‐15 in impacted and non‐impacted cartilage explants. Interestingly, the pro‐mitotic stimulus of exogenous rhGDF‐15 was stronger than that of the cartilage trauma at day 7, whereas no difference could be found between traumatized explants with or without rhGDF‐15 treatment after 14 days (Figure 6H–J). While we observed a significant increase of p21‐positive chondrocytes at day 14 post trauma, long‐term exposure to rhGDF‐15 did not induce the expression of the cell cycle inhibitor in hAC (Figure 6K).
Moreover, we examined the immunomodulatory properties of rhGDF‐15 in activated human macrophages (THP‐1 cells). In accordance with the literature, rhGDF‐15 mitigated the gene expression of TNF and IL1B to some extent but had no effect on IL6 (Figure S7A–C). In hAC exposed to 10 ng/mL IL‐1β, addition of rhGDF‐15 restored the transcription of COL2A1 but had no effect on IL‐1 β‐induced COX2 or MMP13 gene expression (Figure S7D–F).
Taken together, our results imply that exogenous rhGDF‐15 has pro‐regenerative and protective effects after cartilage injury without inducing chondrosenescence.
Discussion
3
Traumatic injuries of joint‐related tissues are considered to carry a high risk for development of a PTOA. However, the underlying pathomechanisms, which are driving the pathogenesis, are complex and not well understood yet.
In the present study we identified the stress‐responsive cytokine GDF‐15 as a potential indicator or even biomarker of PTOA in synovial fluid samples of end‐stage OA patients. Further investigation revealed that GDF‐15 expression was generally upregulated in OA cartilage tissue and highly induced after ex vivo cartilage trauma. Posttraumatic GDF‐15 expression in chondrocytes was found to be triggered by oxidative stress and subsequent p53 activation. Although GDF‐15 was associated with cellular stress and might represent a new SASP factor of senescent chondrocytes, the cytokine induced a pro‐regenerative response, including cell proliferation, migration, and survival, in cartilage tissue.
Regarding our present findings, the strong correlation between syGDF‐15 and cystatin C in PTOA and IOA patients might result from the fact that both GDF‐15 and cystatin C are considered as common age‐ and disease‐related biomarkers. Accordingly, we previously reported that sGDF‐15 was significantly correlated with cystatin C and hs‐CRP within the cohort of the Ulmer OA study, comprising 636 patients suffering from end‐stage hip or knee OA [10]. Moreover, cystatin C and GDF‐15 have lately been described among the three most significant plasma SASP markers in age and were associated with adverse health effects as demonstrated by various physiological parameters, including inflammation (IL‐6 levels), gait speed, and grip strength [26]. It should be noted that the lack of a significant correlation between serum cystatin C and syGDF‐15 in the separate analyses of the PTOA and IOA groups may be attributed to the small sample size.
The stress‐responsive protein p53 is considered as a pivotal regulator in tissue regeneration as described in epithelial and myocardial repair as well as limb regeneration, among others [27, 28, 29]. In this context, p53 orchestrates cell fate decision and plasticity in a concentration‐dependent manner. For one thing, low levels allow cell cycle re‐entry of postmitotic cells and accumulation of progenitor cells, then again, high levels of p53 induce differentiation and transient senescence. In our study, we observed enhanced GDF‐15 expression upon ROS‐induced p53 activation and subsequent senescence of chondrocytes. Usually, accumulation of senescent chondrocytes is considered as one of the major drivers of OA progression, due to the detrimental effects of the secreted SASP factors [14]. However, our findings demonstrated that addition of exogenous rhGDF‐15 does not promote spreading of the senescent phenotype, but, on the contrary, induced proliferation, migration, and cell survival of hAC. Similar results were currently reported in case of IL‐6, which is a common SASP factor and controversially discussed in cartilage degeneration and regeneration [30, 31, 32]. Overall, it is widely accepted that transient senescence contributes to tissue regeneration and that SASP factors initially accelerate related processes, such as immune and stem cell recruitment, fibroblast activation, and ECM remodeling [33]. In cartilage, transient senescence might become chronic due to the tissue‐specific properties, in particularly hypocellularity and avascularity, resulting in a poor tissue regeneration and insufficient clearance of senescent chondrocytes. Consequent accumulation of senescent cells and continuing secretion of SASP mediators has emerged to be highly detrimental [13, 14]. Therefore, it cannot be completely ruled out that the pro‐mitotic effect of GDF‐15 in particularly in combination of other pro‐inflammatory SASP factors and oxidative stress may switch from a pro‐regenerative to an adverse effect, as previously demonstrated for pro‐mitotic TGF‐ß1 and FGF2 in combination with irradiation‐induced DNA damage [34].
Besides the fact that cellular senescence is not per se a pathophysiologic process, it should be noted that GDF‐15 has been described as an anti‐inflammatory and immunomodulatory cytokine. In line with this, a previous case‐control study, including 910 RA patients and healthy controls, identified two GDF‐15 gene polymorphisms, which were associated with an enhanced risk for RA [35]. They hypothesized that the immunomodulatory nature of GDF‐15 might possess anti‐arthritic effects. In fact, the cytokine was assumed to attenuate CXCL10/CXCR3‐dependent infiltration of T lymphocytes during glomerulonephritis [36]. Accordingly, GDF‐15 was found to impair dendritic cell maturation and thus their ability to serve as antigen‐presenting cells during T‐cell priming [37]. Its role as immunomodulatory cytokine might also explain that serum levels of GDF‐15 were reported to correlate with disease activity in RA patients [38]. In our study, we could confirm an increased secretion of GDF‐15 by TCM‐stimulated hFLS, and anti‐inflammatory effects of rhGDF‐15 on activated THP1 macrophages to some extent.
Strikingly, we observed that hAC respond to exogenous rhGDF‐15 and thus provided first evidence of GFRAL, the only member of the GNDF family receptor‐α (GFRα) capable of binding GDF‐15 [39], in cartilage tissue. This implies that rhGDF‐15‐mediated effects result from its interaction with GFRAL. As we found enhanced levels of GFRAL in highly degenerated and traumatized cartilage tissue, we assume an increased susceptibility of chondrocytes to the cytokine in the course of disease progression and trauma response. Although it has been initially hypothesized that GFRAL expression was restricted to the central nervous system [40], more recent studies report the presence of GFRAL in peripheral tissues, such as adipose, pancreatic ductal adenocarcinoma, and gastric cancer tissue [41, 42, 43]. Moreover, we excluded that the response of hAC toward exogenous rhGDF‐15 was mediated via interaction with the TGF receptor or ErbB2, which both have been discussed as alternative receptors of GDF‐15 [44, 45], by addition of the corresponding inhibitors in migration assays (Figure S6B,C).
With regard to the current knowledge about GDF‐15, which is also described as a moonlight protein due to its various, independent functions beyond originally identified ones [46], we are convinced that our results display only a small range of the comprehensive role of GDF‐15 in cartilage biology and pathophysiology. Although our findings are based on human material, thus providing valuable information about the clinical situation, further in vivo studies might be needed to understand the role of GDF‐15 in age‐related and posttraumatic OA development. Evidence of potentially detrimental effects of GDF‐15 could not be found in this study, but cannot be fully excluded as mentioned above. Moreover, the limited sample size in the PTOA group and the fact that the joint trauma occurred a considerable time ago, should be taken into account. Future research should aim to include larger PTOA cohorts and place greater emphasis on the early stages of osteoarthritis pathogenesis in order to evaluate the prognostic potential of GDF‐15.
Altogether, the current study provides first evidence for the p53‐mediated expression of GDF‐15 in chondrocytes upon cellular stress and its potential cell and chondroprotective effects after ex vivo cartilage trauma. Although GDF‐15 expression was induced by the transcription factor p53 and was linked to cellular senescence, we could not observe any adverse impact of the cytokine on cartilage homeostasis. Regarding our findings, we conclude that GDF‐15 might represent a novel marker of PTOA and might initially promote pro‐regenerative processes. Whether GDF‐15 may serve as a future target in OA therapy has to be clarified in subsequent studies.
Material and Methods
4
Clinical Samples From the Ulm Osteoarthritis Study
4.1
Serum, synovial fluid, and synovial tissue samples were collected in the course of the longitudinal Ulm Osteoarthritis Study from patients suffering from advanced OA of the knee as determined by radiographic changes of grade ≥2 according to Kellgren and Lawrence [47], and undergoing unilateral total knee joint replacement (TKR) between January 1995 and December 1996 and stored at –80°C until use. The inclusion criteria (i.e., Caucasian, age <76 years, absence of malignancies, inflammatory diseases, rheumatoid arthritis, or corticosteroid medication; no previous joint replacement) were fulfilled by 54 patients. The patients were assigned in two groups: IOA patients (n = 42) and PTOA (n = 12), which included traumatic meniscus lesion/ligament rupture/knee joint luxation, intraarticular fractures (knee joint, patella, tibia head), general severe knee injury (with joint effusion, hemarthros, etc.). Relevant clinical characteristics of patients included in this study are described in Table 1. Serum concentrations of cystatin C, COMP, and hs‐CRP were detected as previously described [10, 48]. The study was approved by the local Ethics Committee of the University Ulm (No. 40/94 and 164/14) and written informed consent of the patients was given.
Chondrocyte Culture and Ex Vivo Cartilage Trauma Model
4.2
Human cartilage was obtained from OA patients undergoing TRK with informed consent according to the regulations of the ethics committee of the University of Ulm (245/12, 353/18). For the ex vivo cartilage trauma model and chondrocyte isolation, only macroscopically intact cartilage (OARSI ≤ 1) was used.
Human chondrocytes were isolated by enzymatic digestion. Cartilage was minced and pre‐digested with 0.2% pronase for 45 min, followed by an overnight incubation with 0.025% collagenase at 37°C in a rotator. Chondrocytes were cultured in chondrocyte medium (CM) containing 10% FCS and used at passages ≤ 2. Experiments were performed under serum‐reduced conditions (1:1 CM and serum‐free medium (SFM)).
Ex vivo traumatization of cartilage explants was performed as previously described. In brief, 6 mm diameter full‐thickness cartilage explants were prepared. After a resting time of 24 h, explants were impacted by means of our drop tower device with an energy of 0.59 J [18, 19, 21]. Tissue experiments were performed under serum‐free conditions in SFM. Cartilage tissue and chondrocytes were cultured at 37°C, 5% CO_2_, and 95% humidity.
Human FLS Experiments
4.3
Human fibroblast‐like synoviocytes (FLS) were isolated as previously described [18]. In short, synovial tissue of six patients was minced and incubated in 25 mg collagenase type VIII for 90 min at 37°C, filtered (70 µm), and cultured in FLS serum‐containing medium. Cells were used at passages 2–4. After adherence, cells were stimulated with conditioned medium from unimpacted or impacted cartilage (see above), diluted 1:1 in SFM, and cultured at 27°C or 37°C for 4 days.
Medium Composition
4.4
Chondrocyte medium (CM): 1:1 DMEM (1 g/L glucose)/Ham's F12, 10% heat‐inactivated fetal calf serum (FCS), 0.5% penicillin/streptomycin, 2 mM L‐glutamine, 10 µg/mL 2‐phospho‐L‐ascorbic acid trisodium salt.
Serum‐free medium (SFM): DMEM (1 g/L glucose), 1% sodium pyruvate, 0.5% L‐glutamine, 1% non‐essential amino acids, 0.5% penicillin/streptomycin, 10 µg/mL 2‐phospho‐L‐ascorbic acid trisodium salt, 0.1% insulin‐transferrin‐selenium (ITS).
FLS serum‐containing medium: High glucose (4.5 g/L) DMEM, 10% fetal bovine serum, 0.5% L‐glutamine, 0.5% penicillin/streptomycin (Sigma‐Aldrich).
Gene Expression Analysis
4.5
In case of cartilage samples, tissue was snap frozen in liquid nitrogen and pulverized with a microdismembrator S (B. Braun Biotech, Melsungen, Germany) before total RNA isolation by means of the Lipid Tissue Mini Kit (Qiagen, Hilden, Germany). Isolated cells were lysed in RLT buffer, followed by total mRNA isolation using the Qiagen Mini Kit. RNA was reverse transcribed with the Omniscript RT Kit (Qiagen), according to the manufacturer`s protocol.
Quantitative real‐time PCR analysis was performed by using a StepOnePlus Real‐Time PCR System (Applied Biosystems, Darmstadt, Germany). Target transcripts were analyzed by TaqMan Gene Expression Assays or self‐designed primers and the respective Master Mix, as indicated in Table S1.
In brief, relative expression levels of target genes were determined using the 2^−ΔΔCt^ method, by normalization to the endogenous controls (18S rRNA, GAPDH and HPRT1) as previously described [19].
AlamarBlue Assay
4.6
The alamarBlue assay (BioRad, Munich, Germany) was used to measure the metabolic activity of living cells in order to determine proliferation/cytotoxicity. In brief, cells were incubated for 3 h in a 5% alamarBlue solution in serum‐reduced medium at 37°C. The emitted fluorescence was detected at a 550 nm excitation and 590 nm emission by using the multimode microplate reader Infinite M200 Pro (Tecan Deutschland, Crailsheim, Germany).
Scratch Assay
4.7
A scratch assay, also known as wound healing or migration assay, was used to assess migratory activity of chondrocytes in presence of GDF‐15. A straight scratch was applied in a confluent chondrocyte monolayer by using the tip of a 200 µL sterile pipette scraped across the surface. Images were captured after 0, 24, and 48 h.
GDF‐15 ELISA
4.8
To quantify GDF‐15 in synovial fluid samples and cell culture supernatants, the human GDF‐15 ELISA Kit by R&D Systems was used, according to the manufacturer`s protocol. In brief, 100 µL of undiluted samples were applied. In case of synovial fluid, samples were subjected to a hyaluronidase digest (4 mg/mL; 1:1 mixed with sample) for 60 min at 37°C. Influence of the hyaluronidase digest on GDF‐15 detection is provided in Figure S8.
Immunohistochemistry
4.9
For IHC staining, cartilage and synovial tissue was fixed (4% paraformaldehyde) and embedded in paraffin. In brief, tissue sections were dewaxed and rehydrated, followed by H_2_O_2_ incubation for 30 min. For antigen retrieval, sections were incubated in citrate buffer (pH 6.0) at 60°C overnight. Primary antibodies were used as follows: GDF‐15 (Novus Biologicals, Wiesbaden, Germany; NBP1‐81050; 1:100), GFRAL (Invitrogen, PA5‐24545; 1:100), p53 (LS‐Bio, Lynnwood, WA, US; LS‐B7723; 1:100), p21 (Thermo Fisher Scientific, MA5‐14949; 1:50), incubated overnight at 4°C. The staining was performed with the Dako LSAB2 System‐HRP kit (Dako, Glostrup, Denmark). Subsequently, cell nuclei were counter stained by Gill's hematoxylin No. 3 (Sigma‐Aldrich). Documentation was performed with an Axioskop 2 mot plus (Zeiss, Oberkochen, Germany).
Immunofluorescence Staining of p53
4.10
hAC were fixed with formalin, permeabilized with 0.1% Triton, and incubated for 1 h at 37°C with blocking buffer (Agilent Technologies, Waldbronn, Germany). Afterward, cells were stained with anti‐p53 (LSBio, LS‐B7723; 1:250) for 3 h at RT, followed by an incubation with a secondary antibody (abcam, ab150077, Alexa 488, 1:500) for 30 min at RT. Nuclei were counterstained with 0.25 µg/mL Dapi for 15 min. Area, fluorescence, and integrated density of the cells were measured using Fiji (Version 2.1.0/1.53c; open‐source software). To distinguish between specific and nonspecific signals, the average fluorescence of untreated hAC was calculated and then subtracted from the measured fluorescence of each cell. Subsequently, the corrected total cell fluorescence (CTCF) was determined for the p53‐positive cells.
DCFDA Assay
4.11
Analysis of cytoplasmatic ROS levels was performed by means of the DCFDA/H2DCFDA‐Cellular ROS Assay Kit (Abcam). In short, cultured hAC were incubated with a 1 µM DCFDA working solution for 45 min at 37°C. Afterward, samples were analyzed with a fluorescence microscope. The CTCF was assessed as described above.
Live/Dead Cell Cytotoxicity Assay
4.12
To determine the percentage of viable cells, a Live/Dead Viability/Cytotoxicity Assay (Molecular Probes, Invitrogen) was performed. Unfixed tissue sections (0.5 mm thickness) were stained with 1 µM calcein AM and 2 µM ethidium homodimer‐1 for 30 min. After washing in PBS, they were microscopically analyzed by means of a z‐stack module (software AxioVision, Carl Zeiss, Jena, Germany).
DMMB and Griess Assay
4.13
Content of proteoglycan release into culture media (µg/mL) was quantified using the photometric 1,9‐dimethylmethylene blue assay as previously described [19]. Analysis is based upon the binding of sulfated glycosaminoglycan (GAG) in the presence of 0.24 M GuHCl. In case of NO release, levels were determined by quantification of nitrite, a stable end product of the NO metabolism, using a Griess assay (Griess Reagent System; Promega) [49].
RNA Sequencing
4.14
Total RNA from cells was extracted as described above. Quality assessment, library preparation, RNA sequencing, and bioinformatic analysis was carried out by Biomarker Technologies (BMK; Münster, Germany). Libraries were sequenced using an Illumina Novaseq 6000 (PE150) system.
Statistical Analysis
4.15
Data were analyzed using GraphPad Prism9 (GraphPad Software, Inc., San Diego, CA, USA). Datasets with n ≥ 5 were tested for outliers by means of the Grubbs’ outlier test. Outliers were not included in the statistical analyses. Each data point represents an independent biological replicate (donor). Data sets of two groups were analyzed by means of a two‐tailed (multiple) t‐test. In case of data sets with three groups and more, a one‐way ANOVA with Sidak post hoc test (parametric distribution) or Kruskal–Wallis with a Dunn's post hoc test (non‐parametric distribution) was used. Association between two parameters was analyzed by means of a Pearson's (parametric) or Spearman's (non‐parametric) correlation analysis with a Bonferroni‐Holm post‐hoc test in case of multiple comparison. In each case, significance level was set to α = 0.05.
Author Contributions
JR: funding acquisition, study concept and design, data acquisition, interpretation of data, figure preparation, statistical analysis, and writing/editing of the manuscript. SSM: data acquisition, conducting experiments, and participation in manuscript preparation. AL, SM: data acquisition, conducting experiments, and revision of manuscript. RB, DR: administration Ulm OA study, data interpretation, and revision of the manuscript. All authors approved the final version of the manuscript and ensure that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Funding
This research study was supported by the European Social Fund and by the Ministry of Science, Research and Arts Baden‐Württemberg as well as the University of Ulm (Hertha‐Nathorff‐Programm).
Conflicts of Interest
The authors have nothing to declare.
Ethics Statement
The study was approved by the local Ethics Committee of the University Ulm (No. 40/94, 245/12, 164/14, and 353/18) and written informed consent of the patients was given.
Supporting information
Figure S1: Difference of syGDF15 concentrations in female and male patients.Figure S2: Results of the gene expression analysis of GDF‐15 and GFRAL after cartilage trauma.Figure S3: Heatmap presenting significantly upregulated (red) and downregulated (blue) genes after GDF‐15 stimulation as compared to untreated control hAC. “Ctrl” = unstimulated hAC; “Treat” = hAC stimulated with 100 ng/mL rhGDF‐15 for 48 h. Labeling at the right side describes gene names of proteins related to the glycerophospholipid metabolism.Figure S4: Additional findings of RNA‐seq analysis and confirmation of senescence‐associated genes by means of qRT‐PCR.Figure S5: Dose‐dependent effects of rhGDF‐15 on proliferation (alamarBlue) and migration (scratch assay) of hACFigure S6: Influence of GDF‐15 on directed (Boyden chamber) and non‐directed (scratch assay) migrationFigure S7: Anti‐inflammatory/immunomodulatory effects of rhGDF‐15.Table S8: List of primer and TaqMan gene expression assays used for qRT‐PCR.Figure S9: Influence of previous hyaluronidase digestion on concentrations of synovial GDF‐15.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1B. Snoeker , A. Turkiewicz , K. Magnusson , et al., “Risk of Knee Osteoarthritis After Different Types of Knee Injuries in Young Adults: A Population‐Based Cohort Study,” British Journal of Sports Medicine 54, no. 12 (2020): 725–730.31826861 10.1136/bjsports-2019-100959 · doi ↗ · pubmed ↗
- 2C. R. Maia , R. F. Annichino , E. de Azevedo , E. G. Machado , E. Marchi , and M. C. Castano‐Betancourt , “Post‐Traumatic Osteoarthritis: The Worst Associated Injuries and Differences in Patients' profile When Compared With Primary Osteoarthritis,” BMC Musculoskeletal Disorders [Electronic Resource] 24, no. 1 (2023): 568.37438788 10.1186/s 12891-023-06663-9PMC 10337069 · doi ↗ · pubmed ↗
- 3L. E. Bayliss , D. Culliford , A. P. Monk , et al., “The Effect of Patient Age at Intervention on Risk of Implant Revision After Total Replacement of the Hip or Knee: A Population‐Based Cohort Study,” Lancet 389, no. 10077 (2017): 1424–1430.28209371 10.1016/S 0140-6736(17)30059-4PMC 5522532 · doi ↗ · pubmed ↗
- 4A. C. Thomas , T. Hubbard‐Turner , E. A. Wikstrom , and R. M. Palmieri‐Smith , “Epidemiology of Posttraumatic Osteoarthritis,” Journal of Athletic Training 52, no. 6 (2017): 491–496.27145096 10.4085/1062-6050-51.5.08PMC 5488839 · doi ↗ · pubmed ↗
- 5Y. Song , Y. Cui , X. Zhang , et al., “Increased Serum Levels of MIC 1/GDF 15 Correlated With Bone Erosion in Spondyloarthritis: A Pilot Study,” Medicine 97, no. 51 (2018): e 13733.30572513 10.1097/MD.0000000000013733 PMC 6320148 · doi ↗ · pubmed ↗
- 6W. D. Xu , Q. Huang , C. Yang , R. Li , and A. F. Huang , “GDF‐15: A Potential Biomarker and Therapeutic Target in Systemic Lupus Erythematosus,” Frontiers in Immunology 13 (2022): 926373.35911685 10.3389/fimmu.2022.926373 PMC 9332889 · doi ↗ · pubmed ↗
- 7D. A. Brown , J. Moore , H. Johnen , et al., “Serum Macrophage Inhibitory Cytokine 1 in Rheumatoid Arthritis: A Potential Marker of Erosive Joint Destruction,” Arthritis and Rheumatism 56, no. 3 (2007): 753–764.17328047 10.1002/art.22410 · doi ↗ · pubmed ↗
- 8J. Wischhusen , I. Melero , and W. H. Fridman , “Growth/Differentiation Factor‐15 (GDF‐15): From Biomarker to Novel Targetable Immune Checkpoint,” Frontiers in Immunology 11 (2020): 951.32508832 10.3389/fimmu.2020.00951 PMC 7248355 · doi ↗ · pubmed ↗