First report of VGSC mutations for resistance to synthetic pyrethroids in brown dog ticks (Rhipicephalus sanguineus sensu stricto) from Brazil
Hayden Kleinschmidt, Nathan E. Stone, Rebecca Ballard, Natalie B. Thornton, Grant L. Pemberton, Simone Becker, Guilherme M. Klafke, David M. Wagner, Joseph D. Busch

TL;DR
This study reports the first occurrence of VGSC gene mutations linked to pyrethroid resistance in brown dog ticks from Brazil.
Contribution
First report of VGSC mutations in Rh. sanguineus s.s. ticks from the Americas, confirmed via genetic analysis.
Findings
Mitochondrial gene analysis confirmed the ticks were Rh. sanguineus s.s.
Two known resistance mutations (C190A and G215T) were identified in the VGSC gene.
These mutations are the first reported in Rh. sanguineus s.s. ticks in the Americas.
Abstract
Brown dog ticks, Rhipicephalus sanguineus sensu lato (s.l.), have spread globally and are an important vector of multiple pathogens affecting both dogs and humans. The control of these ticks on dogs and human dwellings often relies on synthetic pyrethroids, a chemical class of acaricides that targets the voltage-gated sodium channel (VGSC) protein of arthropod nerve cells, causing influxes of sodium and ultimately, paralysis. Invasive Rhipicephalus sanguineus s.l. ticks in the Americas can be broadly grouped into two main lineages: temperate and tropical (Rh. sanguineus sensu stricto (s.s.) and Rh. linnaei, respectively). Phenotypic resistance to synthetic pyrethroids in a verified Rh. sanguineus s.s. population has only been reported in the state of Rio Grande do Sul, Brazil. The objectives of our study were to 1) screen a small number (n = 10) of Rh. sanguineus s.s. from Brazil to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
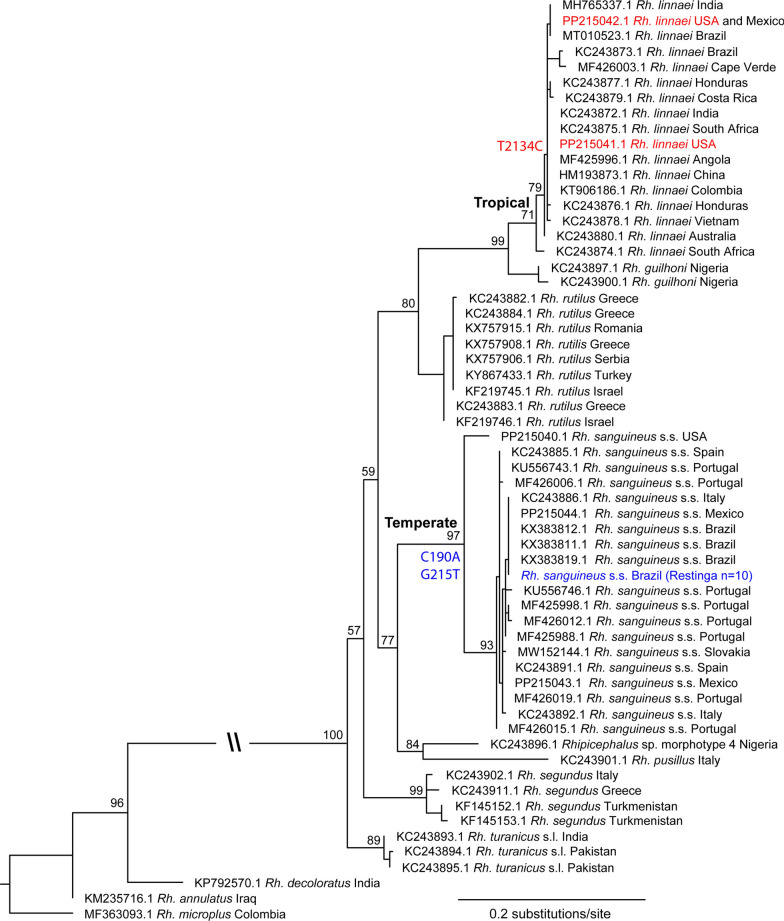 Figure 1
Figure 1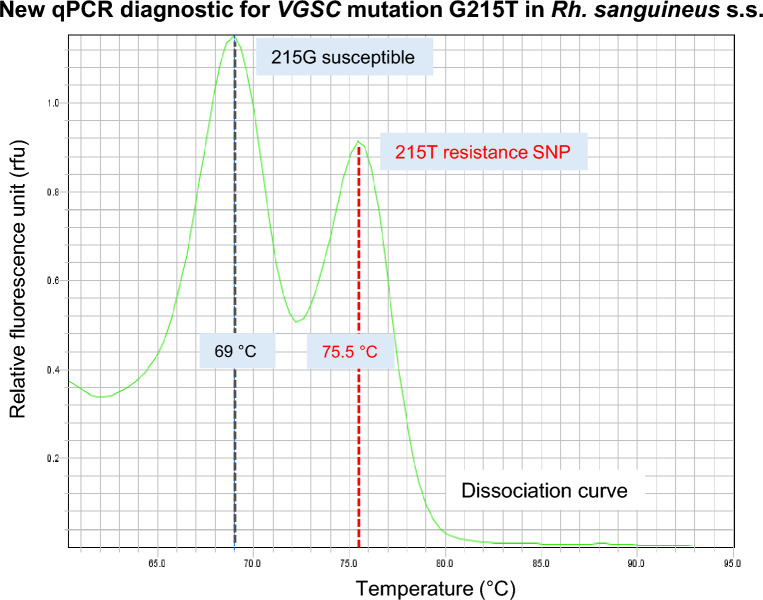 Figure 2
Figure 2- —https://doi.org/10.13039/100008883Northern Arizona University
- —https://doi.org/10.13039/501100003593Conselho Nacional de Desenvolvimento Científico e Tecnológico
- —https://doi.org/10.13039/100020365Centers for Disease Control and Prevention Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Bartonella species infections research · Vector-Borne Animal Diseases
Background
Brown dog ticks, Rhipicephalus sanguineus sensu lato (s.l.), comprise a group of related tick species [1, 2] that have spread globally with domestic dogs via human transport. These ticks are important vectors of the canine pathogens Babesia vogeli and Ehrlichia canis [3] and transmit a number of rickettsial species in the spotted fever group to humans, including Rickettsia rickettsii (causative agent of Rocky Mountain spotted fever) [4, 5], R. massiliae in the southwestern USA and Mexico [6], and R. conorii in Europe [7]. Current methods for disease prevention rely heavily on chemical control of ticks on dogs. Brown dog ticks can live in and around human dwellings and prefer to feed on domestic dogs throughout their three life stages [8–10]. Two main lineages have spread throughout the Americas: Rh. sanguineus sensu stricto (s.s.), commonly referred to as “temperate” lineage ticks [11], and the “tropical” lineage, recently named Rh. linnaei in some parts of the world [12]. Other Rh. sanguineus s.l. taxa are found in Asia and Europe [1, 13]. In Brazil, Rh. sanguineus s.l. has been shown to carry R. rickettsii [14] and is a suspected vector for transmission to humans [15]. Stray dogs in urban environments such as Porto Alegre facilitate the persistence and spread of Rh. sanguineus s.s., which is the most common tick species found on dogs in the state of Rio Grande do Sul [16].
Synthetic pyrethroids are a chemical class of acaricides commonly used throughout the world to control tick infestations [17], including brown dog ticks [18]. Resistance to synthetic pyrethroids such as deltamethrin, permethrin, and cypermethrin is usually due to nonsynonymous single-nucleotide polymorphisms (SNPs) in the voltage-gated sodium channel (VGSC) gene that is essential for proper nerve functioning in arthropod species [19], although metabolic detoxification can also play a role [20–22]. Phenotypic resistance to synthetic pyrethroids has been described in various Rh. sanguineus s.l. tick populations throughout the Americas, including Brazil [16], Panama [22], the Caribbean [23], and the USA [18, 24]. Furthermore, a resistance SNP (T2134C) in domain III segment 6 of the VGSC gene has been described in Rh. linnaei samples from multiple states of the USA [25, 26], and this same mutation has been described in a synthetic pyrethroid-resistant population of Rh. sanguineus s.l. from Florida [27]. Additional SNPs conferring resistance to bioassays for deltamethrin and flumethrin have been reported in brown dog ticks from India, including C190A and G215T in domain II segments 4–5 of the VGSC [28]. The C190A and T2134A SNPs are common in a related tick, Rh. (Boophilus) microplus, due to frequent use of these chemicals to treat ticks on cattle [26, 29, 30]. However, little is known about mutations conferring acaricide resistance in Rh. sanguineus s.s. populations in Brazil despite the past use of synthetic pyrethroids there. In this study, we screened a small collection of brown dog ticks from Brazil [16] for any of the known resistance mutations in the VGSC gene.
Methods
DNA was extracted from 10 brown dog ticks (three adult females and seven nymphs) leftover from a previous study [16] conducted by the Instituto de Pesquisas Veterinárias Desidério Finamor (IPVDF), Eldorado do Sul, Brazil (Additional file 1). These ticks (hereafter referred to as Restinga ticks) were collected from a single stray dog in the Restinga sub-district of Porto Alegre, Brazil [31]. Although the 10 ticks were not tested for phenotypic resistance to deltamethrin, six engorged females sampled from the same dog produced an F1 larval generation that demonstrated a low level of resistance to deltamethrin (resistance ratio 5.67) in larval packet tests. The history of acaricide exposure in prior generations leading to this field tick population is unknown.
All molecular procedures followed the methods from two previous studies [25, 26]; primers specific to VGSC targets were designed in Stone et al. [26] and updated in Thomas et al. [32]. Amplicon sequencing (AmpSeq) of three PCR targets was performed on each sample to obtain partial sequences of VGSC [25]. Because resistance to fipronil was detected in other brown dog tick (BDT) samples from the metropolitan area of Porto Alegre [16], we included an additional PCR target for the GABA-gated chloride channel (GABA-Cl), which contains mutations associated with resistance to fipronil and dieldrin [33–36]. The Restinga ticks had previously been identified as Rh. sanguineus s.s. on the basis of morphology according to the Barros-Battesti guide [31] and their location in far southern Brazil [37]. To confirm the species identity and provide a phylogeographic context, we generated one amplicon each for three mitochondrial genes (12S rRNA, 16S rRNA, and COI). The seven targets were amplified in two multiplex PCRs as outlined in Stone et al. [25] and pooled within each individual, then indexed and sequenced on an Illumina NextSeq 1000 instrument using a P1 600 cycle (2 × 300) Reagent Kit with PhiX control (Illumina, San Diego, CA, USA, part# 20100981), with the goal of obtaining hundreds to thousands of reads per amplicon for each individual. Bioinformatics methods for removing errors, SNP calling, and identifying heterozygotes versus homozygotes are described in our recent study of Rh. sanguineus [25]. Mitochondrial sequences for COI were aligned against publicly available sequences of Rh. sanguineus s.l. using BioEdit (v7.1.3) [38] and a maximum likelihood phylogeny was inferred with IQ-TREE (v2.2.0.3) [39] with the “-bb” bootstrapping option (n = 1000) and the integrated ModelFinder method [40]. Presumptive resistance SNPs were mapped onto major phylogenetic clades using Adobe Illustrator. To confirm the presence of VGSC mutations in domain II segments 4–5, real-time genotyping PCR assays were performed [26, 32], followed by Sanger sequencing of domain II segments 4–5, as previously described [26]. Sequences for COI, 12S, and 16S were also aligned against a small set of reference sequences [25] and assigned a haplotype (Additional file 1). Finally, a new real-time genotyping assay for G215T was developed for this study (Additional file 1) following previously described methods [26, 41].
Results
Analysis of partial COI sequences (346 bp) grouped the Restinga ticks together in a clade with multiple Rh. sanguineus s.s. (temperate lineage) reference samples (Fig. 1), confirming the previous identification on the basis of morphology and geographical location. The sensu stricto classification was also confirmed by the 12S (339 bp) and 16S (215 bp) sequences (Additional file 1). All 10 Restinga ticks carried identical sequences at each of the three mitochondrial gene targets (Additional file 1). A total of 2 of the 10 ticks carried nonsynonymous SNPs in domain II segments 4–5 of the VGSC; both were heterozygotes (Additional file 1). The first (Rs_Restinga_04) carried the well-described synthetic pyrethroid resistance SNP C190A that leads to amino acid replacement L925I, and the second (Rs_Restinga_07) carried a putative resistance SNP G215T that leads to amino acid replacement G933V (aka G72V). The ticks included in this study were screened against an existing qPCR assay for the C190A SNP as well as the new G215T qPCR assay developed in this study, which confirmed the presence of these resistance genotypes. Both ticks carried only one of the resistance SNPs and were homozygous for susceptible states at other VGSC positions associated with resistance to synthetic pyrethroids in ticks (i.e., nucleotide positions 170 and 2134). Sanger sequencing of all 10 ticks confirmed the qPCR results. Sequences from the GABA-Cl gene suggested the presence of three gene copies: alpha, beta, and gamma (Additional file 1), as reported previously for Rh. sanguineus s.l. [25]. Six Restinga ticks carried allele 2 of the GABA-Cl-beta copy (see Additional file 1; RsGABAchannel_beta_A02), which contains amino acid substitution A301S that is associated with dieldrin/fipronil resistance in other arthropods [42]. However, phenotypic resistance to fipronil was not tested in any Restinga samples, and it remains unknown if this allele is associated with resistance in Rh. sanguineus s.s.Fig. 1. Maximum likelihood phylogenetic tree of Rhipicephalus sanguineus sensu lato based on mitochondrial COI partial sequence (346 bp), with resistance mutations mapped onto terminal branches. Two Rh. sanguineus s.l. clades that are invasive in South America (tropical and temperate) are labeled at interior nodes; bootstrap confidence values from 1000 permutations are labeled above major branches. Font colors denote ticks with resistance SNPs in VGSC domain II segments 4–5 (C190A or G215T; blue font) or domain III segment 6 (T2134C; red font)
Discussion
The partial COI sequences verified that these Restinga tick samples were Rh. sanguineus s.s., which was expected because this species has been described previously in Rio Grande do Sul, Brazil [37], where the temperate climate is predicted to be favorable for this lineage [43]. Here, we report the first confirmed instance of VGSC resistance SNPs C190A and G215T from Rh. sanguineus s.s. ticks in the Americas. Both SNPs have been associated with synthetic pyrethroid resistance in other arthropods [19]; however, it remains unknown whether C190A and G215T convey resistance in Rh. sanguineus s.s. These mutations have been identified previously in two brown dog ticks from India [28], but mitochondrial lineage analysis was not reported in that study, so it is unknown if those ticks also were Rh. sanguineus s.s. Although the Restinga sample size is small, our findings underscore the urgent need to investigate VGSC SNPs in Brazil and elsewhere in South America by conducting in-depth studies to confirm phenotype–genotype correlations.
Synthetic pyrethroids have been widely used for tick (and flea) prevention on domestic dogs for > 25 years [18], and resistance SNPs in Rh. linnaei ticks sampled from companion animals are widespread in the USA [25]. The Restinga ticks were sampled from a stray dog with an unknown history of ectoparasite treatment; the presence of ticks carrying two independent resistance SNPs may indicate that prior tick generations had experienced synthetic pyrethroid exposure at some point, most likely when parasitizing pet dogs. It is also possible that metabolic detoxification [22] is an important mechanism of resistance in brown dog tick populations in Brazil. Regardless, the discovery of multiple resistance SNPs in Rh. sanguineus s.s. and Rh. sanguineus s.l. from different parts of the world emphasizes the need to develop alternative approaches for managing tick populations, such as anti-tick vaccines [44].
Mutations in the VGSC gene can have a strong phenotype–genotype correlation [30] that allows the prediction of synthetic pyrethroid resistance in field tick populations using only genotypes. We were not able to directly test for a phenotype–genotype correlation because no DNA was available from the F1 larvae used in larval packet tests (LPTs) [16]. Despite our small sample size, we note that a low deltamethrin resistance ratio (5.67) in the Restinga dog ticks is consistent with finding only two heterozygous ticks that carried putative VGSC resistance SNPs. When acaricides are used frequently and without a systematic management plan, a single heterozygous mutation conferring resistance against a widely used acaricide such as deltamethrin could lead to large-scale spread of resistant ticks in urban areas.
Conclusion
This study provides the first molecular evidence of putative VGSC resistance mutations in Rh. sanguineus s.s. from South America, highlighting the importance of using applied genetic tools that can 1) identify additional nonsynonymous SNPs in the VGSC (e.g., AmpSeq) and 2) rapidly screen for known mutations using real-time PCR [26, 32]. To accommodate this need for the G215T SNP, we developed a new assay in this study (Additional file 1). Both Rh. sanguineus s.s. and Rh. linnaei have colonized Brazil [37] and other countries in South America [45], and there is a need to monitor for resistance to synthetic pyrethroid acaricides used in the control of brown dog ticks. Future work in Brazil by IPVDF will expand the geographic scope of brown dog tick collections across both urban and rural areas, and a key objective is to integrate molecular analyses with standardized phenotypic assays (e.g., larval packet tests) to validate the predictive value of resistance-associated SNPs in the VGSC. These efforts will be complemented by epidemiological studies mapping the distribution of acaricide resistance and tick-borne pathogens. Together, these initiatives aim to establish an evidence-based framework for controlling Rh. sanguineus infestations in southern Brazil, particularly in densely populated urban areas. The use of molecular tools to monitor resistance levels (via genotype) will contribute to more effective management of tick populations, and by extension, reduce the occurrence of diseases transmitted by brown dog ticks.
Supplementary Information
** Additional file 1: Tables S1, S2, and S3.**Genotypes generated for ten Rhipicephalus sanguineus sensu stricto ticks from Restinga, Brazil.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dantas-Torres F, de Sousa-Paula LC, Otranto D. The Rhipicephalus sanguineus group: updated list of species, geographical distribution, and vector competence. Parasit Vectors. 2024;17:540. 10.1186/s 13071-024-06572-3.10.1186/s 13071-024-06572-3PMC 1168166239731169 · doi ↗ · pubmed ↗
- 2Obaid MK, Islam N, Alouffi A, Khan AZ, da Silva Vaz I, Jr., Tanaka T, et al. Acaricides resistance in ticks: Selection, diagnosis, mechanisms, and mitigation. Front Cell Infect Microbiol. 2022;12:1-20. 10.3389/fcimb.2022.941831.10.3389/fcimb.2022.941831 PMC 929943935873149 · doi ↗ · pubmed ↗
- 3Duscher GG, Galindo RC, Tichy A, Hummel K, Kocan KM, de la Fuente J. Glutathione S-transferase affects permethrin detoxification in the brown dog tick, Rhipicephalussanguineus. Ticks Tick Borne Dis. 2014;5:225–33. 10.1016/j.ttbdis.2013.11.006.10.1016/j.ttbdis.2013.11.00624548526 · doi ↗ · pubmed ↗
- 4Nepveu-Traversy ME, Fausther-Bovendo H, Babuadze GG. Human tick-borne diseases and advancesin anti-tick vaccine approaches: A comprehensive review. Vaccines. 2024;12 2:1-38. 10.3390/vaccines 12020141.10.3390/vaccines 12020141 PMC 1089156738400125 · doi ↗ · pubmed ↗