Approaches to murine T cell isolation and activation
Sangjun Lim, Sohyun Kum, Jin Ouk Choi, Joonbeom Bae, Soo Seok Hwang

TL;DR
This paper provides a guide for isolating and activating mouse T cells in a lab setting.
Contribution
The paper offers a reproducible framework for murine T cell preparation and activation.
Findings
Standard methodologies and troubleshooting insights are integrated for T cell preparation.
The guide supports both basic immunological studies and T cell engineering applications.
Abstract
In this MiniResource, we outlined practical approaches for preparing murine T cells, from isolation to in vitro activation, with emphasis on reproducibility and viability. While specific experimental conditions should be tailored to individual assays, the principles summarized here provide a framework for establishing robust T cell preparation across diverse research settings. By integrating standard methodologies with troubleshooting insights, this resource aims to support both basic immunological studies and future applications in T cell engineering.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
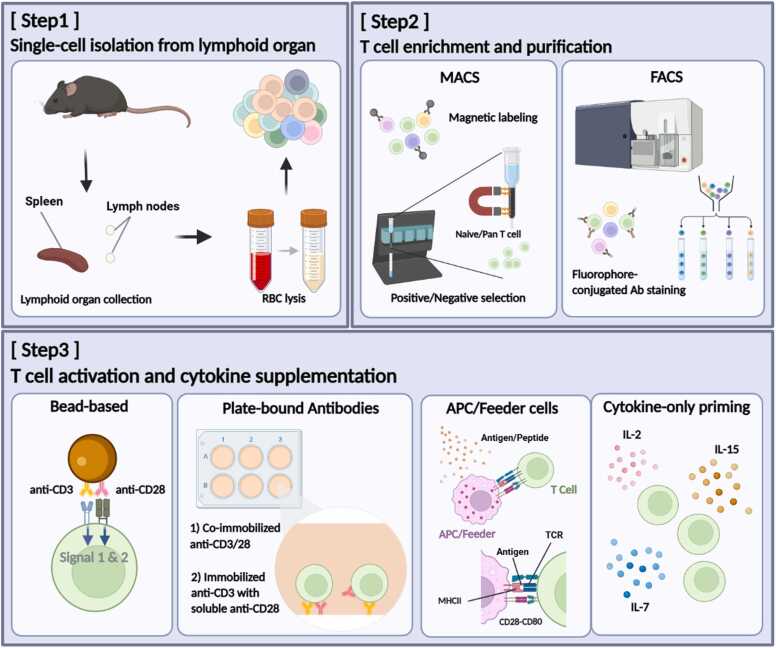 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCAR-T cell therapy research · Immune Cell Function and Interaction · T-cell and B-cell Immunology
INTRODUCTION
T cells are central players in adaptive immunity, governing host defense, immune regulation, and therapeutic responses. Reliable preparation of T cells in mouse models underpins both mechanistic research and translational immunology. Over the years, diverse methodological approaches have been developed to improve yield, purity, and reproducibility in T cell isolation and activation within mouse models. These include magnetic-associated cell separation (MACS) and fluorescence-activated cell sorting (FACS) of lymphoid organ-derived single-cell suspensions, followed by antibody- or antigen-presenting cell (APC)-based activation systems with cytokine supplementation (Miltenyi et al., 1990, Sutermaster and Darling, 2019, Trickett and Kwan, 2003; Fig. 1). In this MiniResource, we provide practical guidelines for isolating and activating murine T cells, particularly for researchers less familiar with primary T cell handling. This article also serves as the entry point of a 2-part series: here we focus on establishing a high-quality T cell population, while the subsequent article addresses viral transduction strategies for genetic engineering, such as transduction for chimeric antigen receptor-T cell (CAR-T) development (Ayala Ceja et al., 2024, Vormittag et al., 2018).Fig. 1. Schematic overview of murine T cell isolation/enrichment and in vitro activation. (Step1) Lymphoid organs are harvested, mechanically dissociated to generate single-cell suspensions, with red blood cells (RBCs) lysed. (Step2) T cells are enriched and/or purified mostly by MACS (magnetic-activated cell sorting; positive or negative selection) or by FACS (fluorescence-activated cell sorting) following staining with fluorophore-conjugated antibodies. (Step3) T cell receptor (TCR) and costimulatory signals can be delivered using antibody-coated beads or plate-bound antibodies; alternatively, primary antigen-presenting cells (APCs) provide peptide-MHC (major histocompatibility complex) together with costimulatory ligands (CD80/CD86) to approximate physiological stimulation.Fig. 1
METHODS FOR T CELL ISOLATION
Secondary lymphoid organs, such as spleens and lymph nodes, are standard sources for T cell preparation (Malhotra et al., 2013). To obtain single-cell suspension, tissues are gently dissociated on a 70-µm strainer and washed with cold phosphate-buffered saline, preferably with the addition of fetal bovine serum to enhance cell viability. Following cell collection by centrifugation, red blood cells are removed with ammonium-chloride-potassium lysing buffer to enrich lymphocyte populations and improve isolation efficiency. The resulting cells are subsequently resuspended and filtered once more through a nylon mesh to remove cellular aggregates.
MACS and FACS are among the most widely used approaches for T cell isolation (Table 1). MACS uses antibody-conjugated microbeads that target the cognate antigen expressed on the cell surface (Grutzkau and Radbruch, 2010, Khurana et al., 2022). Two principal strategies are employed for T cell isolation: positive and negative selection. In positive selection, T cells are directly labeled using lineage markers such as CD3, CD4, or CD8. Negative selection method targets other cell subsets, thereby depleting non-T cells or unintended T cell populations. Both strategies are easily accessible, showing moderately high yield and purity along with simple workflow. However, it should be noted that the positive selection method may interfere the activation state of T cells via cell-bound antibodies, and the residual beads cannot be avoided when directly performing imaging and flow cytometric analysis (Laghmouchi et al., 2020).Table 1. Comparison of methods for murine T cell isolationTable 1MethodPrinciplesStrengthLimitationsYield/purityRecommended useMACS (positive selection)Cells conjugated with specific antibodies-bead conjugates are strongly capturedHigh purity, simple workflowTargeting CD3 or TCR can induce unintended activation; beads may affect downstream workflowVariable yield;purity > 95%Preclinical CAR studies, needing high purity of chimeric antigen receptor transduced T cellsMACS (negative selection)Antibody-bead cocktails deplete non-T lineages, leaving unlabeled T cells “untouched”Preserve native signaling state; minimal receptor engagementHighly dependent on kit quality; costlyHigh yield;purity: 90%-95%Functional assays (proliferation, differentiation, engineering), unbiased activationFACSDroplet-based high-purity physical sortingPrecise subset-specific sorting; removal of apoptotic and dead cellsTime-consuming;requires highly expensive FACS instrumentation;lower total yieldVariable yield;purity > 98%Subset-specific mechanistic studiesCAR, chimeric antigen receptor; FACS, fluorescence-activated cell sorting; MACS, magnetic-associated cell separation.
FACS can significantly enhance the purity of T cell isolation (Schraivogel et al., 2022). As various fluorochrome-conjugated antibodies can be utilized for this method depending on the purpose, precise and subset-specific T cell isolation is available (Matsumoto et al., 2023). This is particularly useful for mechanistic studies of naïve, memory, or regulatory T cells. Nonetheless, relatively reduced yield and viability due to prolonged processing may affect downstream assays (Cossarizza et al., 2021, Yi et al., 2023). To address this issue, pre-enrichment with MACS before FACS can mitigate these drawbacks (Cossarizza et al., 2021).
Suboptimal experimental conditions can compromise cell viability during T cell isolation, subsequently impairing T cell integrity and functionality. To minimize this issue, all experimental steps should be carried out without delays, and cell suspensions must be carefully handled under controlled temperature (4°C) to avoid apoptosis. Processing steps that impose mechanical or chemical stress, such as tissue dissociation, require optimization to reduce cellular damage.
APPROACHES TO T CELL ACTIVATION AND CYTOKINE SUPPLEMENTATION
T cell activation begins when TCR signaling exceeds the activation threshold upon recognition of peptide-MHC on APCs (Irvine et al., 2002, Zhu et al., 2010). This event drives naïve T cells from quiescence into clonal expansion and differentiation (Au-Yeung et al., 2014, Hwang et al., 2020). Various in vitro activation strategies provide a relatively homogeneous starting population for controlled functional studies and downstream applications such as CAR-T engineering (Zhang et al., 2017; Table 2).Table 2. Approaches to T cell activation and cytokine supplementationTable 2MethodPrinciplesCytokineStrengthLimitationsNotesBead-based (anti-CD3/CD28)Beads immobilized with anti-CD3/CD28 enforce uniform polyclonal activationIL-2±IL-7 or IL-15Highly reproducible;scalable;simple hands-on workflowBead carry-over can interfere with imaging/flow; induce overactivation due to crosslinkingAdjust bead/cell ratio to intended activation strengthPlate-bound AbsAnti-CD3 is adsorbed to tissue-culture plate; costimulation is added as soluble or immobilized anti-CD28IL-2Precise control over stimulus strength; inexpensive;no beads removal step neededRelatively inconsistent activation strength due to well-to-well variability from coating heterogeneityCoating density critical for cell viability and consistencyAntigen-presenting cellsDelivering physiologic multisignal activation and enabling antigen-specific expansionIL-2, IL-15Physiological multisignal stimulation; antigen-specific expansionVariable;time-consuming due to isolation of APC and/or maturation/differentiationMore suitable for translational experiments; peptide pulsing process requiredCytokine-only primingMaintains cells with resting stateIL-7, IL-15Preserves stem-like stateSlow activation;lower yield due to non-expansionUseful for Tscm or memory-like T cell preservationAPC, antigen-presenting cell; Tscm, T memory stem cells.
A simple and practical way to monitor activation is by examining cell morphology under a light microscope. Activated T cells undergo blastogenesis, becoming visibly larger than resting cells. This size increase is typically evident by ∼20 hours poststimulation, providing a convenient checkpoint before proceeding to downstream assays such as viral transduction or functional testing.
Antibody-based systems mimic the 2-signal requirement: TCR engagement via anti-CD3 (Signal 1) and costimulation via anti-CD28 (Signal 2) (Jenkins and Johnson, 1993). Plate-bound methods require immobilized anti-CD3 with soluble or immobilized anti-CD28, whereas bead-based systems provide more uniform and sustained activation, though at higher cost. Because activation strength strongly affects outcomes ranging from poor proliferation to functional exhaustion, empirical titration of stimulation dose is essential (Wang et al., 2008).
Alternatively, activation can be achieved using APCs, particularly dendritic cells (Jobin et al., 2025, Storni and Bachmann, 2003). This approach mimics physiological interactions and is suitable for translational studies. However, variability and the labor-intensive preparation of APCs—requiring isolation, differentiation, and antigen loading—limit routine use. Moreover, APC-derived cytokines and costimulatory ligands may confound experiments focused on T cell–intrinsic mechanisms (Gutcher and Becher, 2007).
Cytokine supplementation (Signal 3) is indispensable for sustained activation and survival (Mescher et al., 2006). IL-2 provides essential survival and proliferation signals through STAT5-dependent pathways (Ross and Cantrell, 2018, Villarino et al., 2022). For prolonged culture, IL-7 and IL-15 improve viability and promote memory-like characteristics (Choi et al., 2022, Pilipow et al., 2015, Tan et al., 2001). When maintenance of a quiescent state is required, IL-7 and IL-15 in the absence of TCR stimulation can support cell viability while preserving a resting phenotype (Choi et al., 2025, Sprent and Surh, 2011).
CONCLUDING REMARKS
We outlined core methodologies for murine T cell isolation and activation, emphasizing reproducibility and viability across experimental contexts. While individual conditions require optimization, the principles highlighted here offer a practical foundation for downstream applications such as functional assays and T cell engineering. For issues related to yield and viability, troubleshooting strategies are available for consultation (Table 3). Read in tandem with the following MiniResource on viral transduction, these protocols together provide an integrated pipeline from T cell preparation to genetic modification.Table 3. Troubleshooting guide for T cell isolation and activationTable 3ProblemPossible causesSolutionLow T cell viability after single-cell isolationMechanical and chemical stress during single-cell suspension
-
•Tissue must be grinded more gently
-
•Add ice-cold PBS with serum as needed during tissue grinding to reduce friction-induced heat
-
•Adjust ACK lysis duration/buffer volume Low postsort yield during FACSExcessively high event rate of sample acquisition during sorting procedure
-
•Adjust the flow rate to minimize cell loss
-
•Pre-enrich the sample with MACS using less antibodies/beads Reduced T cell viability after in vitro TCR stimulationStrong TCR signaling may cause activation-induced cell death
-
•Optimize appropriate dose for anti-CD3/28 Cell death after prolonged cultureDepletion of nutrients in the culture mediumExhaustion of T cells due to long exposure of TCR stimulation
-
•Adjust cell density before T cell stimulation
-
•Change or add media
-
•Adjust proper days of culture and once T cells are activated, maintain them in anti-CD3/CD28-free medium to prevent exhaustion (supply suitable cytokines) ACK, ammonium-chloride-potassium; FACS, fluorescence-activated cell sorting; MACS, magnetic-associated cell separation; PBS, phosphate-buffered saline.
Author Contributions
Sangjun Lim: Writing – review & editing, Writing – original draft. Jin Ouk Choi: Writing – review & editing, Funding acquisition. Sohyun Kum: Writing – original draft, Visualization. Soo Seok Hwang: Writing – review & editing, Writing – original draft, Supervision, Project administration, Funding acquisition, Conceptualization. Joonbeom Bae: Writing – review & editing, Funding acquisition.
Declaration of Competing Interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Au-Yeung B.B.Zikherman J.Mueller J.L.Ashouri J.F.Matloubian M.Cheng D.A.Chen Y.Shokat K.M.Weiss A.A sharp T-cell antigen receptor signaling threshold for T-cell proliferation Proc. Natl. Acad. Sci. U. S. A.111352014 E 367936882513612710.1073/pnas.1413726111 PMC 4156735 · doi ↗ · pubmed ↗
- 2Ayala Ceja M.Khericha M.Harris C.M.Puig-Saus C.Chen Y.Y.CAR-T cell manufacturing: major process parameters and next-generation strategies J. Exp. Med.22122024 e 2023090310.1084/jem.20230903 PMC 1079154538226974 · doi ↗ · pubmed ↗
- 3Choi J.O.Ham J.H.Hwang S.S.RNA metabolism in T lymphocytes Immune Netw.2252022 e 3910.4110/in.2022.22.e 39PMC 963414236381959 · doi ↗ · pubmed ↗
- 4Choi J.O.Seo Y.Hwang S.S.Guardians of silence: transcriptional networks in T cell quiescence Exp. Mol. Med.5782025166316724075974010.1038/s 12276-025-01516-y PMC 12411625 · doi ↗ · pubmed ↗
- 5Cossarizza A.Chang H.D.Radbruch A.Abrignani S.Addo R.Akdis M.Andra I.Andreata F.Annunziato F.Arranz E.Guidelines for the use of flow cytometry and cell sorting in immunological studies (third edition)Eur. J. Immunol.51122021270831453491030110.1002/eji.202170126 PMC 11115438 · doi ↗ · pubmed ↗
- 6Grutzkau A.Radbruch A.Small but mighty: how the MACS-technology based on nanosized superparamagnetic particles has helped to analyze the immune system within the last 20 years Cytometry A 77720106436472058327910.1002/cyto.a.20918 · doi ↗ · pubmed ↗
- 7Gutcher I.Becher B.APC-derived cytokines and T cell polarization in autoimmune inflammation J. Clin. Invest.11752007111911271747634110.1172/JCI 31720 PMC 1857272 · doi ↗ · pubmed ↗
- 8Hwang S.S.Lim J.Yu Z.Kong P.Sefik E.Xu H.Harman C.C.D.Kim L.K.Lee G.R.Li H.B.m RNA destabilization by BTG 1 and BTG 2 maintains T cell quiescence Science 36764832020125512603216558710.1126/science.aax 0194 · doi ↗ · pubmed ↗