Two new species of Chionosphaera and Kurtzmanomyces (Chionosphaeraceae, Agaricostilbomycetes) isolated from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
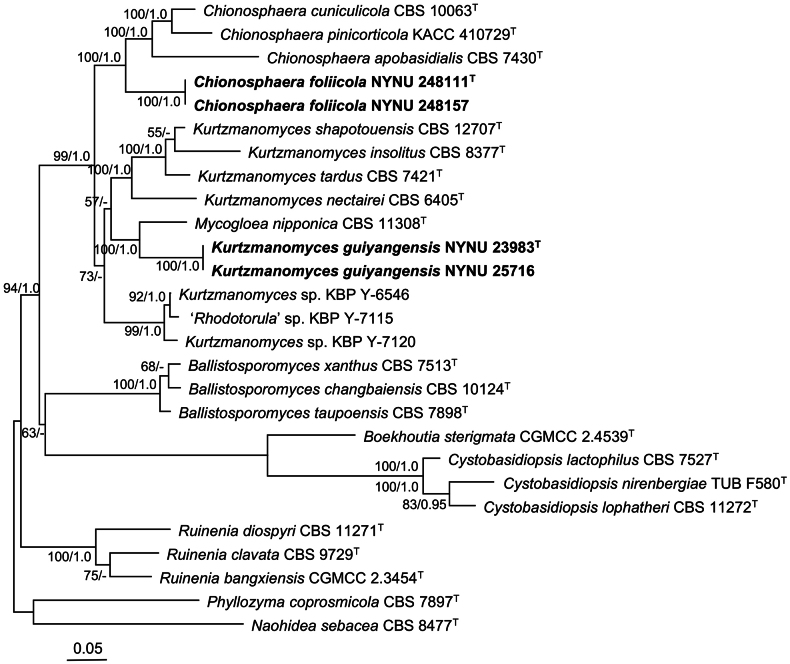 Figure 1
Figure 1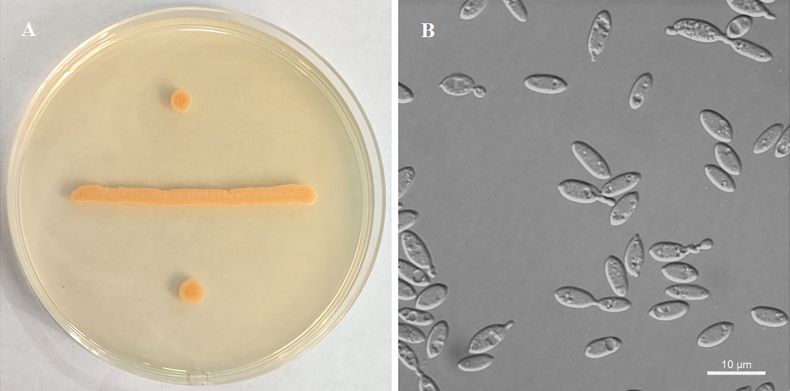 Figure 2
Figure 2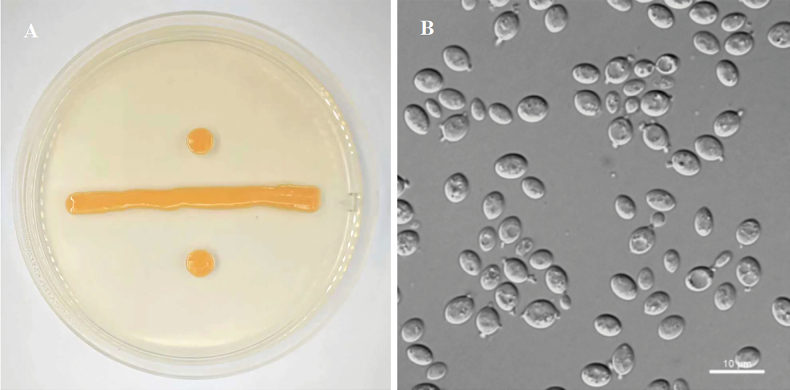 Figure 3
Figure 3| Species | Strain no. | Locality | GenBank accession no. | ||
|---|---|---|---|---|---|
|
|
|
| |||
|
| CBS 10124T | China |
|
| — |
|
| CBS 7898T | Japan |
|
|
|
|
| CBS 7513T | Japan |
|
|
|
|
| CGMCC 2.4539T | China |
|
| — |
|
| CBS 7430T | USA |
|
|
|
|
| CBS 10063T | Germany |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| KACC 410729T | South Korea |
|
| — |
|
| CBS 7527T | Japan |
|
|
|
|
| CBS 11272T | Taiwan, China |
|
|
|
|
| TUB F580T | Germany |
|
| — |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CBS 8377T | Portugal |
|
|
|
|
| CBS 6405T | Japan |
|
|
|
|
| CBS 12707T | China |
|
| — |
|
| CBS 7421T | Portugal |
|
|
|
| KBP Y-7120 | Russia |
|
| — | |
| ‘ | KBP Y-7115 | Russia |
|
| — |
| KBP Y-6546 | Russia |
|
| — | |
|
| CBS 11308 | — |
|
|
|
|
| CGMCC 2.3454T | China |
|
|
|
|
| CBS 9729T | China |
|
|
|
|
| CBS 11271T | Taiwan, China |
|
|
|
|
| CBS 8477T | UK |
|
|
|
|
| CBS 7897T | New Zealand |
|
|
|
| Characteristic | 1 | 2* | 3* | 4* | 5 | 6* | 7* | 8* | 9* | 10* |
|---|---|---|---|---|---|---|---|---|---|---|
| Fermentation of | ||||||||||
| Glucose | – | – | n | n | – | – | – | – | + | – |
| Galactose | – | n | n | n | – | – | – | – | + | – |
| Sucrose | – | n | n | n | – | – | – | – | + | – |
| Raffinose | – | n | n | n | – | – | – | – | + | – |
| Trehalose | – | n | n | n | – | – | – | – | + | – |
| Assimilation of | ||||||||||
| Inulin | d | s | + | – | + | – | – | – | + | – |
| Sucrose | + | – | – | – | + | + | + | – | + | + |
| Raffinose | – | – | – | – | – | – | + | – | + | + |
| Galactose | d | w | w | – | – | – | – | – | + | + |
| Lactose | d, w | – | – | – | – | – | + | – | – | – |
| Trehalose | + | v | + | + | + | – | + | + | + | + |
| Maltose | + | – | – | s | – | – | – | – | – | + |
| Melezitose | + | – | + | – | + | – | + | – | + | + |
| Cellobiose | + | – | w | – | – | – | + | – | – | + |
| Salicin | w | v | – | – | + | – | S | – | – | s |
| L-Sorbose | d, w | + | w | v | – | – | – | – | + | – |
| D-Xylose | d, w | – | – | v | – | – | + | – | + | s |
| L-Arabinose | d, w | s | w | v | + | – | + | – | + | – |
| D-Arabinose | w | + | w | – | + | – | + | – | + | – |
| D-Ribose | – | – | – | – | + | – | S | – | + | s |
| Glycerol | d, w | + | + | – | – | – | S | – | – | s |
| Erythritol | – | – | – | – | – | – | S | – | – | + |
| Ribitol | w | v | + | s | + | – | S | + | + | + |
| D-Glucitol | d, w | s | + | v | + | w, s | – | + | n | – |
| Growth tests | ||||||||||
| 0.1 % Cycloheximide | – | n | n | n | – | – | + | – | – | – |
| Vitamin-free medium | w | s, w | n | n | – | – | – | – | + | – |
| Growth at 30 °C | + | + | n | w, – | – | + | + | – | + | + |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Infectious Diseases and Mycology
Introduction
Chionosphaeraceae (Agaricostilbomycetes, Agaricostilbales) is an ecologically significant family of basidiomycetous fungi with a broad environmental distribution. The family was originally established to accommodate teleomorphic taxa characterized by gasteroid basidia bearing basidiospores simultaneously on each basidium (Oberwinkler and Bandoni 1982; Bauer et al. 2006). Wang et al. (2015) revised the family to include five genera—Ballistosporomyces, Chionosphaera, Cystobasidiopsis, Kurtzmanomyces, and Stilbum—based on a phylogeny inferred from six gene loci. More recently, the genus Boekhoutia was added to the family (Li et al. 2020).
Chionosphaera, the type genus of Chionosphaeraceae, was first described by Cox (1976) and comprises species primarily associated with arboreal environments (Kyung and Kwon-Chung 2011). The genus was initially placed in the family Filobasidiaceae, but was later transferred to the newly established family Chionosphaeraceae based on molecular phylogenetic evidence (Oberwinkler and Bandoni 1982; Hibbett 2006). Wang et al. (2015) recognized six species within the genus: C. apobasidialis, C. coppinsii, C. cuniculicola, C. erythrinae, C. lichenicola, and C. phylaciicola. Subsequent molecular analyses led to the reclassification of C. coppinsii and C. lichenicola into the newly established genus Crittendenia within Pucciniomycotina, thereby excluding them from Chionosphaeraceae (Millanes et al. 2021). Currently, five species are accepted in the genus, including C. pinicorticola, a recently described species from South Korea (Lee et al. 2025). Among these, only C. apobasidialis, C. cuniculicola, and C. pinicorticola are known from living cultures, all exhibiting asexual yeast morphs (Kyung and Kwon-Chung 2011; Lee et al. 2025). The sexual morph occurs exclusively on natural substrates and forms minute, white, stipitate-capitate basidiocarps, typically bearing four to eight one-celled, hyaline, oblong to reniform or ellipsoid to cylindrical basidiospores (Kyung and Kwon-Chung 2011). In contrast, the asexual morph produces slightly mucoid, cream-colored colonies that reproduce by budding; some species may form pseudohyphae and/or true hyphae. Physiologically, Chionosphaera species do not ferment sugars or assimilate nitrate, usually react positively with diazonium blue B (DBB), and show weak or no urease activity (Lee et al. 2025).
Kurtzmanomyces was established by Yamada et al. (1988) to accommodate the asexual, stalk-producing yeast species K. nectairii, based on phenotypic characteristics. This taxonomic placement was later supported by molecular phylogenetic analyses (Fell et al. 2000). The genus was subsequently expanded to include three additional species: K. tardus and K. insolitus from Portugal (Giménez-Jurado et al. 1990; Sampaio et al. 1999), and K. shapotouensis from China (Zhang et al. 2013). Phylogenetic studies have shown that the sexual species Mycogloea nipponica is closely related to Kurtzmanomyces and produces a Kurtzmanomyces-like anamorph (Kirschner et al. 2003; Sampaio 2011). Species of Kurtzmanomyces are characterized by pale orange colonies and the production of blastoconidia at the tips of stalk-like conidiophores. Some species may also form ballistoconidia and septate hyphae (Yamada et al. 1988; Sampaio 2011). Physiologically, members of the genus generally lack fermentative ability, possess ubiquinone Q-10 as the major respiratory quinone, and can assimilate a variety of carbon sources, but not myo-inositol or D-glucuronate (Sampaio 2011).
During a survey of Chionosphaeraceae in China, four yeast isolates were obtained from plant leaf samples collected in Guizhou and Hainan Provinces. Molecular analyses based on the internal transcribed spacer (ITS) region, the D1/D2 domain of the large subunit (LSU) rRNA gene, and the translation elongation factor 1-α gene (tef1α), combined with physiological and biochemical data revealed that these isolates represent two novel species in the genera Chionosphaera and Kurtzmanomyces. The new taxa are described and illustrated below.
Materials and methods
Sample collection and yeast isolation
Plant leaf samples were collected from East Mountain Park (26°45'26"N, 106°21'31"E) in Guiyang City, Guizhou Province, and from Wuzhi Mountain (18°17'21"N, 109°40'55"E) in Wuzhishan City, Hainan Province, China. Out of the 19 collected plant leaf samples, strains NYNU 248111 and NYNU 248112 were isolated from two different leaf samples of Symplocos adenophylla collected in Wuzhi Mountain, while NYNU 23983 and NYNU 25716 were recovered from Debregeasia orientalis and Glochidion sp. in East Mountain Park and Wuzhi Mountain, respectively. These yeasts were isolated from the leaf surfaces using the washing and dilution method described by Jiang et al. (2024). Fresh leaves were cut into small pieces using sterile scissors, placed into sterile 10 mL centrifuge tubes, and suspended in sterile water containing 0.05 % (v/v) tween 80. The samples were shaken for 10 min, and the washing solution was serially diluted to 10^−2^. An aliquot of 200 μL from each dilution was spread onto yeast extract–malt extract (YM) agar medium (0.3 % yeast extract, 0.3 % malt extract, 0.5 % peptone, 1 % glucose, and 2 % agar) supplemented with 200 μg/mL chloramphenicol. The plates were then incubated at 20 °C until visible colonies developed. Colonies with distinct yeast morphologies were selected and purified by streaking onto fresh YM agar plates. Purified strains were suspended in 20 % (v/v) glycerol and stored at −80 °C for long-term preservation.
Phenotypic studies
Morphological observations and physiological and biochemical tests were conducted following the methods described by Kurtzman et al. (2011). Carbon and nitrogen assimilation tests were performed twice in liquid media and monitored for up to 4 weeks. Nitrogen assimilation was tested using starved inocula. To induce sexual reproduction, single or double strains were mixed on corn meal agar (CMA: 2.5 % corn starch and 2 % agar), potato dextrose agar (PDA: 20 % potato infusion, 2 % glucose, and 2 % agar), and V8 agar (10 % V8 juice and 2 % agar) at 17 °C for 6 weeks, with observations made every two weeks (Li et al. 2020). Ballistoconidium formation was assessed by the inverted-plate method (do Carmo-Sousa and Phaff 1962) using CMA at 17 °C for two weeks. Temperature tolerance was determined by culturing on YM agar at 15, 20, 25, 30, 35, and 37 °C.
DNA extraction, PCR amplification, sequencing
Genomic DNA was extracted from yeast cultures using the Ezup Column Yeast Genomic DNA Purification Kit (Sangon Biotech, Shanghai, China) following the manufacturer’s instructions. The ITS region, the D1/D2 domain of the LSU rRNA gene, and the partial tef1α gene were amplified with primers ITS1/ITS4 (White et al. 1990), NL1/NL4 (Kurtzman and Robnett 1998), and EF1-526F/EF1-1567R (Rehner and Buckley 2005), respectively. Each 25 μL PCR reaction consisted of 9.5 μL nuclease-free water, 12.5 μL of 2 × Taq PCR Master Mix with blue dye (Sangon Biotech, Shanghai, China), 1 μL genomic DNA template, and 1 μL of each primer. PCR amplification was conducted as described by Toome et al. (2013) for ITS and LSU. For tef1α, we used a touchdown PCR protocol as described (Wang et al. 2014). PCR products were analyzed by electrophoresis on 1 % agarose gels. Amplicons showing clear single bands were purified and sequenced by Sangon Biotech Co., Ltd. (Shanghai, China). Forward and reverse sequences were edited and assembled into consensus sequences using BioEdit v.7.1.3.0 (Hall 1999). All newly obtained sequences were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank/).
Sequence alignment and phylogenetic analyses
The obtained sequences were compared against the GenBank database using the BLASTn algorithm to identify closely related taxa (Altschul et al. 1997). For phylogenetic inference, all newly generated sequences were combined with reference sequences retrieved from GenBank (Table 1), following the taxon sampling frameworks proposed by Zhang et al. (2013) and Lee et al. (2025). The species of the genus Stilbum are not included in phylogenetic analyses because sequence data for type species of this genus are presently not available from public databases. Naohidea sebacea CBS 8477 and Phyllozyma coprosmicola CBS 7897 were designated as the outgroups. The ITS, LSU, and tef1α sequences were aligned separately using MAFFT v.7.110 (Katoh and Standley 2013) with the G-INS-i algorithm. Sequence alignments were visually inspected and manually refined in BioEdit v.7.1.3.0 (Hall 1999) to improve positional homology. The final ITS, LSU, and tef1α alignments were concatenated into a single dataset using PhyloSuite v.1.2.3 (Zhang et al. 2020) for subsequent phylogenetic analyses.
Maximum Likelihood (ML) and Bayesian Inference (BI) analyses were conducted using RAxML v.8.2.3 (Stamatakis 2014) and MrBayes v.3.2.7a (Ronquist et al. 2012), respectively. The ML analysis was performed under the GTR+G+I model, and branch support was evaluated with 1000 rapid bootstrap (BS) replicates. For BI, six Markov Chain Monte Carlo (MCMC) chains were run for 50 million generations under the best-fit substitution models selected using jModelTest (Posada 2008), with trees sampled every 1000 generations. The first 25 % of trees were discarded as burn-in, and the remaining trees were used to calculate Bayesian posterior probabilities (BPPs). Phylogenetic trees were visualized in FigTree v.1.4.3 (Andrew 2016), with support values indicated for nodes with BS ≥ 50% and BPPs ≥ 0.95.
Results
Molecular phylogeny
In this study, 12 newly generated ITS, LSU, and tef1α sequences, together with 58 reference sequences retrieved from GenBank (Table 1), were included in the phylogenetic analyses. After removal of 656 ambiguously aligned regions (131 regions from ITS, 59 regions from LSU, and 466 regions from tef1α), the final concatenated alignment comprised 2332 base pairs, including 691 bp from ITS, 629 bp from LSU, and 1012 from tef1α. The ML and BI analyses yielded congruent topologies; therefore, only the ML tree is presented (Fig. 1). All Chionosphaeraceae taxa formed a strongly supported monophyletic clade (BS = 94 %, BPPs = 1.0). The four newly analyzed strains were grouped into two distinct lineages, which are here recognized as two putative novel species in the genera Chionosphaera and Kurtzmanomyces, respectively.
Phylogenetic tree of Chionosphaeraceae based on the ITS, LSU, and tef1α dataset. Bootstrap support (BS) ≥ 50% and Bayesian posterior probabilities (BPPs) ≥ 0.95 are shown. Naohidea sebacea CBS 8477 and Phyllozyma coprosmicola CBS 7897 were selected as the outgroups. New species are marked with bold.
Strains NYNU 248111 and NYNU 248112, both isolated from Symplocos adenophylla collected in Wuzhi Mountain, exhibited identical ITS and LSU sequences, indicating that they are conspecific. Phylogenetic analyses placed these strains within the genus Chionosphaera, closely related to C. cuniculicola, C. pinicorticola, and C. apobasidialis (Fig. 1). However, the novel strains differed from these species by 37–68 nucleotide (nt) substitutions (~6.2–11.1 %) in the LSU region and by more than 58–80 nt differences (~9.6–13.3 %) in the ITS region. These levels of sequence divergence support their recognition as a distinct taxon. Although it cannot be ruled out that these strains represent the asexual morph of an already-known parasite with no sequence information available, it is meaningful to erect a new species in our opinion. Therefore, the name Chionosphaera foliicola sp. nov. is proposed for these two strains.
Strains NYNU 23983 and NYNU 25716 were isolated from Debregeasia orientalis and Glochidion sp. in East Mountain Park and Wuzhi Mountain, respectively. They were placed in the Kurtzmanomyces clade with an affinity to M. nipponica CBS 11308, a reference material of M. nipponica, based on phylogenetic analyses of the ITS, LSU, and tef1α sequences (Fig. 1). They differed from M. nipponica CBS 11308 by 36 nt substitutions (~5.3 %) in the LSU region and by more than 59 nt differences (~10.1 %) in the ITS region. These molecular differences, together with physiological and biochemical characteristics (see below), support its recognition as a distinct species. Due to the unavailability of M. nipponica type strain in reference culture collections (Sampaio 2011), we recommend keeping it classified as M. nipponica until additional molecular data are available. Therefore, we propose the name Kurtzmanomyces guiyangensis sp. nov. for this novel taxon.
Taxonomy
Chionosphaera
foliicola
Taxon classificationFungiAgaricostilbomycetesChionosphaeraceae
C.Y. Chai & F.L. Hui sp. nov.
3DE5FD4B-C68A-5380-9EA0-9CB64D12D347
MB 859682
Etymology.
The specific epithet foliicola refers to the ex-type strain isolated from a leaf.
Morphology of Chionosphaera foliicola (NYNU 248111). A. Colony on YM agar after 7 days at 20 °C; B. Budding cells in YM broth after 3 days at 20 °C.
Typus.
China • Hainan Prov.: Wuzhishan City, Wuzhi Mountain, in the phylloplane of Symplocos adenophylla, August 2024, S.L. Lv, NYNU 248111 (holotype GDMCC 2.531^T^ preserved as a metabolically inactive state, culture ex-type PYCC 10141).
Description.
On YM agar after 7 days at 25 °C, the streak culture is pale orange tough, smooth and glossy with an entire margin. In YM broth after 3 days at 25 °C, cells are ellipsoidal to cylindrical (2.7–6.0 × 4.1–10.5 µm) and occur singly or in pairs. Budding is polar through minute stalks, normally one to two per cell. After a month in YM broth at 20 °C, a sediment is formed, but no ring and pellicle are observed. After 2 weeks in Dalmau plate culture on CMA at 20 °C, hyphae were observed. Sexual structures are not observed on PDA, CMA or V8 agar at 17 °C. Ballistoconidia are not produced. Sugar fermentation is absent. The following carbon sources are assimilated: glucose, inulin (delayed), sucrose, D-galactose (delayed), lactose (weak and delayed), trehalose, maltose, melezitose, methyl α-D-glucoside (delayed), cellobiose, salicin (weak), L-sorbose (weak and delayed), L-rhamnose (weak and delayed), D-xylose (weak and delayed), L-arabinose (weak and delayed), D-arabinose (weak), 5-keto-D-gluconate, ethanol (weak), glycerol (weak and delayed), ribitol (weak), galactitol, D-mannitol, D-glucitol (weak and delayed), DL-lactate (weak and delayed), succinate, citrate (weak and delayed), and d-glucono-1,5-lactone. Raffinose, melibiose, d-ribose, methanol, erythritol, myo-inositol, D-glucosamine, N-acetyl-D-glucosamine, 2-keto-d-gluconate, and D-glucuronate are not assimilated. L-Lysine is assimilated as sole nitrogen sources. Nitrate, nitrite, ethylamine, and cadaverine are not assimilated. Growth is observed at 30 °C, but not at 35 °C. Growth in the presence of 0.01 % cycloheximide, 10 % NaCl plus 5 % glucose as well as 1% acetic acid is negative. Growth in the vitamin-free medium is positive. Growth on 50 % (w/w) glucose-yeast extract agar is negative. There is no hydrolysis of urea and no starch formation. The DBB reaction is positive.
Additional strain examined.
China • Hainan Prov.: Wuzhishan City, Wuzhi Mountain, in the phylloplane of Symplocos adenophylla, August 2024, S.L. Lv, NYNU 248157.
GenBank accession numbers.
Holotype GDMCC 2.531^T^ (ITS: PV770167, LSU: PQ571723; tef1α: PX046189); additional strain NYNU 248157 (ITS: PV774782, LSU: PV774781; tef1α: PX046190).
Note.
Physiologically, C. foliicola sp. nov. differs from the closely related species, C. apobasidialis, C. cuniculicola, and C. pinicorticola, in its ability to assimilate sucrose and lactose (Table 2).
Kurtzmanomyces
guiyangensis
Taxon classificationFungiAgaricostilbomycetesChionosphaeraceae
C.Y. Chai & F.L. Hui sp. nov.
583A6DD5-B967-5C6D-BABF-99BDF35E3D08
MB 859684
Etymology.
The specific epithet guiyangensis refers to the geographic origin of the ex-type strain, Guiyang City, Guizhou Province.
Morphology of Kurtzmanomyces guiyangensis (NYNU 23983). A. Colony on YM agar after 7 days at 20 °C; B. Budding cells in YM broth after 3 days at 20 °C.
Typus.
China • Guizhou Prov.: Guiyang City, East Mountain Park, in the phylloplane of Debregeasia orientalis, August 2023, D. Lu, NYNU 23983 (holotype GDMCC 2.499^T^ preserved as a metabolically inactive state, culture ex-type PYCC 9992).
Description.
On YM agar after 7 days at 20 °C, streak cultures are pale orange, smooth and butyrous, and the margin is entire. In YM broth after 3 days at 20 °C, cells are spherical to ovoid (3.4–4.7 × 4.5–6.8 µm) and occur singly or in pairs. The conidiogenous stalks, normally one to three per cell, are short, 0.4–1.0 µm. After a month in YM broth at 17 °C, a sediment and a ring are formed, but no pellicle is observed. After 7 days in Dalmau plate culture on CMA at 17 °C, pseudohyphae are absent. Sexual structures are not observed on PDA, CMA or V8 agar at 17 °C. Ballistoconidia are not produced. Glucose fermentation is absent. The following carbon sources are assimilated: glucose, inulin, sucrose, trehalose, melezitose, salicin, L-arabinose, D-arabinose, 5-keto-D-gluconate, D-ribose, ethanol, ribitol, galactitol, D-mannitol, D-glucitol, and succinate. Raffinose, melibiose, D-galactose, lactose, maltose, methyl α-D-glucoside, cellobiose, L-sorbose, L-rhamnose, D-xylose, methanol, glycerol, erythritol, myo-inositol, DL-lactate, citrate, D-glucuronate, D-glucosamine, N-acetyl-D-glucosamine, 2-keto-D-gluconate, and D-glucono-1,5-lactone are not assimilated. Nitrate and nitrite are weakly assimilated as sole nitrogen sources. Ethylamine, l-lysine, and cadaverine are not assimilated. Growth is observed at 25 °C, but not at 30 °C. Growth in the presence of 0.01 % cycloheximide, 10 % NaCl plus 5 % glucose as well as 1 % acetic acid is negative. Growth in the vitamin-free medium is absent. Growth on 50 % (w/w) glucose-yeast extract agar is positive. Starch-like compounds are not synthesized. Diazonium blue B reactions is positive.
Additional strain examined.
China • Hainan Prov.: Wuzhishan City, Wuzhi Mountain, in the phylloplane of Glochidion sp., July 2025, S.L. Lv, NYNU 25716.
GenBank accession numbers.
Holotype GDMCC 2.499^T^ (ITS: OR961459, LSU: OR958742, tef1α: PX046187); additional strain NYNU 25716 (ITS: PX046462, LSU: PX046461; tef1α: PX046188).
Note.
Physiologically, K. guiyangensis sp. nov. differs from the closely related species M. nipponica in its ability to assimilate inulin, L-arabinose, D-arabinose, and D-glucitol and its inability to assimilate raffinose, galactose, maltose, and glycerol. Furthermore, M. nipponica is able to grow at 30 °C, whereas K. guiyangensis sp. nov. does not exhibit growth at this temperature. A comparative summary of the distinguishing physiological characteristics between K. guiyangensis sp. nov. and other Kurtzmanomyces species is presented in Table 2.
Discussion
In this study, four strains belonging to the family Chionosphaeraceae were isolated during an investigation of leaf-inhabiting fungi from different regions of China. Based on a polyphasic approach combining physiological, biochemical, and molecular data, two novel species were proposed in the genera Chionosphaera and Kurtzmanomyces. The phylogenetic analyses presented here are consistent with previous studies (Zhang et al. 2013; Lee et al. 2025) and contribute additional insights into the taxonomy and phylogenetic relationships within Chionosphaeraceae.
Chionosphaera species predominantly inhabit tree bark and other natural substrates. For instance, C. apobasidialis was isolated from decayed bark of Quercus macrocarpa, C. cuniculicola from beetles in coniferous forests, C. erythrinae from leaves of Erythrina tomentosa, and C. pinicorticola from pine bark, collectively indicating a preference for arboreal habitats (Cox 1976; Kirschner et al. 2001; Kirschner and Chen 2008; Lee et al. 2025). In line with this ecological tendency, the newly described species C. foliicola sp. nov. was isolated from the phylloplane of Symplocos adenophylla. Although its complete ecological role in situ remains to be fully understood, its occurrence in a similar arboreal environment suggests that it shares ecological preferences with its congeners.
To date, four asexual species have been described in the genus Kurtzmanomyces. These species are rarely encountered in nature, with most known only from a single isolate. K. insolitus was isolated from the basidiocarp of an unidentified Exidiopsis species in Portugal (Sampaio et al. 1999), K. nectairei from cheese in France (Rodrigues de Miranda, 1975), K. shapotouensis from soil crusts in China (Zhang et al. 2013), and K. tardus from contaminated demineralized water in Portugal (Giménez-Jurado et al. 1990). In addition, Mycogloea nipponica, which possesses a Kurtzmanomyces-like anamorph, appears to be a mycoparasite on ascomycetes and has been reported from Japan and Taiwan (Bandoni 1998; Kirschner et al. 2003). Furthermore, at least three unpublished or misidentified yeasts —Kurtzmanomyces sp. KBP Y-7120, ‘Rhodotorula’ sp. KBP Y-7115, and Kurtzmanomyces sp. KBP Y-6546 —may represent additional novel species of the genus (Fig. 1). All three were isolated from the lichen Cladonia stellaris in Russia. In this study, K. guiyangensis sp. nov. was recovered from the leaf surfaces of Debregeasia orientalis and Glochidion sp. in different regions of China. This distinct ecological origin suggests that K. guizhouensis occupies a different ecological niche compared to previously known members of the genus Kurtzmanomyces. This expands the known ecological range of Kurtzmanomyces and underscores the importance of exploring diverse habitats for uncovering yeast diversity.
Supplementary Material
XML Treatment for Chionosphaera foliicola
XML Treatment for Kurtzmanomyces guiyangensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Altschul SF Madden TL Schäffer AA Zhang J Zhang Z Miller W Lipman DJ (1997) Gapped BLAST and PSI-BLAST: A new generation of protein database search programs.Nucleic Acids Research 25(17): 3389–3402. 10.1093/nar/25.17.33899254694 PMC 146917 · doi ↗ · pubmed ↗
- 2Andrew R (2016) Fig Tree: Tree figure drawing tool Version 1.4.3. Institute of Evolutionary Biology, United Kingdom, University of Edinburgh Press.
- 3Bandoni RJ (1998) On some species of Mycogloea.Mycoscience 39: 31–36. 10.1007/BF 02461575 · doi ↗
- 4Bauer R Begerow D Sampaio JP WeißM Oberwinkler F (2006) The simple-septate basidiomycetes: A synopsis.Mycological Progress 5: 41–66. 10.1007/s 11557-006-0502-0 · doi ↗
- 5Cox DE (1976) A new homobasidiomycete with anomalous basidia.Mycologia 68: 481–510. 10.1080/00275514.1976.12019937 · doi ↗
- 6do Carmo-Sousa L Phaff HJ (1962) An improved method for the detection of spore discharge in the Sporobolomycetaceae.Journal of Bacteriology 83(2): 434–435. 10.1128/jb.83.2.434-435.196216561932 PMC 277751 · doi ↗ · pubmed ↗
- 7Fell JW Boekhout T Fonseca A Scorzetti G Statzell-Tallman A (2000) Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit r DNA D 1/D 2 domain sequence analysis.International Journal of Systematic and Evolutionary Microbiology 50(3): 1351–1371. 10.1099/00207713-50-3-135110843082 · doi ↗ · pubmed ↗
- 8Giménez-Jurado G Plácido T Cidadão AJ Cabeça-Silva C Fonseca E Roeijmans H Jvan Eijk G Wvan Uden N (1990) Kurtzmanomyces tardus sp. nov., a new anamorphic yeast species of basidiomycetous affinity.Antonie van Leeuwenhoek 58: 129–135. 10.1007/BF 004227292264723 · doi ↗ · pubmed ↗