The addition of six novel species and a new record of Amphisphaeria from northern Thailand

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3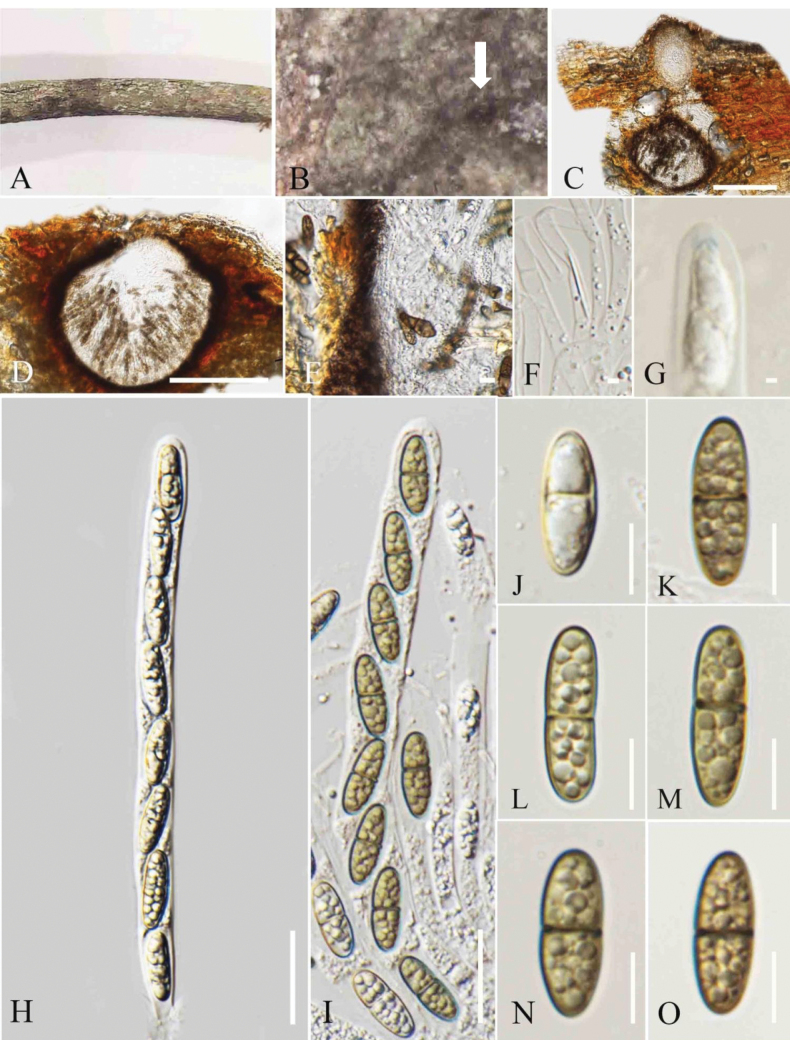 Figure 4
Figure 4 Figure 5
Figure 5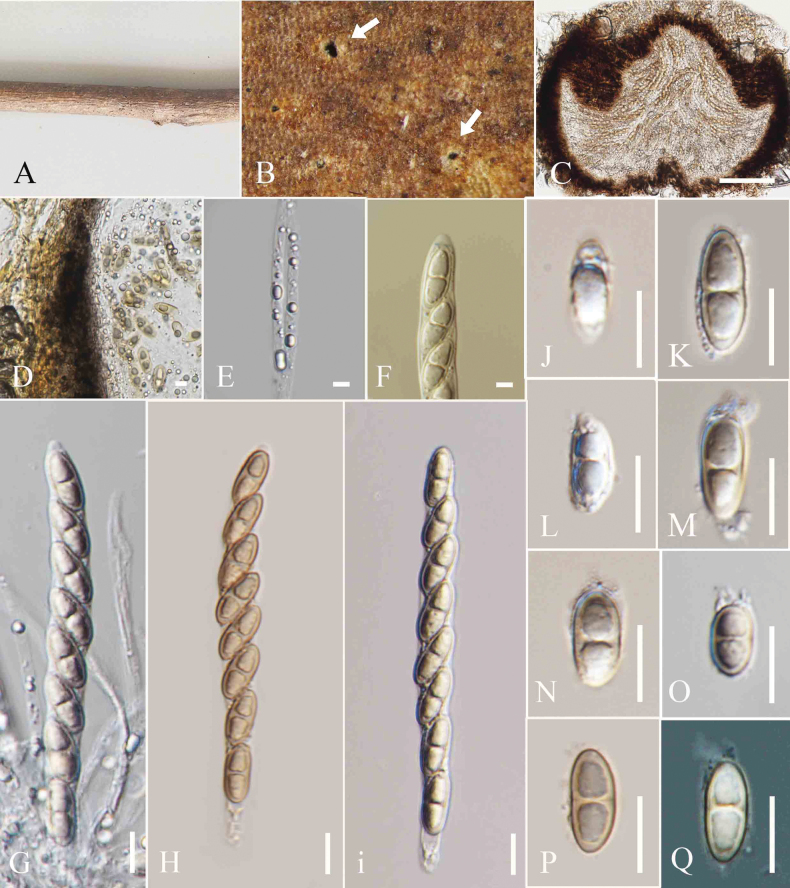 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8| Gene region | Primer pairs | References | |
|---|---|---|---|
|
| LR0R/LR5 | 95 °C/30 s, 55 °C/50 s, 72 °C/60 s | Vilgalys and Hester (1999) |
|
| ITS5/ITS4 | 95 °C/30 s, 55 °C/50 s, 72 °C/60 s |
|
| Taxon | Strain |
|
| Reference |
|---|---|---|---|---|
|
|
|
|
| |
|
|
|
|
| |
|
| KUNCC 23-15521 |
|
|
|
|
| KUNCC 23-15520* |
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
| CGMCC3.23740* |
|
|
|
|
|
|
|
| |
|
| N/A |
|
| |
|
| N/A |
| ||
|
| N/A |
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
| MHZU 24-0515 |
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
| KUNCC 23-15522* |
|
|
|
|
| KUNCC 23-15523 |
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
| PUFD37 |
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
| N/A |
|
| |
|
|
|
| ||
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
| N/A |
|
| ||
|
|
|
|
| |
|
|
|
| ||
|
| N/A |
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
| KUNCC 23-15525 |
|
|
|
|
| KUNCC 23-15524* |
|
|
|
|
|
|
|
| |
|
|
|
|
| |
|
|
|
| ||
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
| |
|
| N/A |
|
| |
|
|
|
|
| Species | Host | Country | References |
|---|---|---|---|
|
| On a branch of | Italy |
|
|
| NA | China |
|
|
| NA | China |
|
|
| In cavities at the end of a rotting log | USA |
|
|
| China |
| |
|
| NA | Thailand |
|
|
| NA | Thailand |
|
|
| USA, | ||
|
| The medullary tissue of the lichen | China |
|
|
| Austria, Germany | De not 1865; | |
|
| NA | Thailand |
|
|
| On attached branches of | Germany, Austria, Belgium | |
|
| Dead leaves of | Australia |
|
|
| NA | Thailand |
|
|
| China |
| |
|
| NA | China |
|
|
| NA | Thailand |
|
|
| NA | China |
|
|
| NA | China |
|
|
| Portugal |
| |
|
| NA | China |
|
|
| Thailand | This study | |
|
| China, Thailand | ||
|
| Thailand | This study | |
|
| New Zealand | Petrak (1923); | |
|
| On branches of | China |
|
|
| The healthy roots of | China |
|
|
| NA | Germany | |
|
| Thailand | This study | |
|
| NA | China |
|
|
| NA | Thailand | This study |
|
| On bark of | Italy |
|
|
| Thailand | This study | |
|
| NA | China |
|
|
| England, France, Germany | ||
|
| Thailand | This study | |
|
| On leaf of | USA | |
|
| NA | China |
|
|
| On branch of | Italy |
|
|
| NA | Thailand |
|
|
| Switzerland |
| |
|
| NA | China | Tusi et al. (2001) |
|
| Branches of | China |
|
|
| On the stem of | Canada, British, Columbia | |
|
| NA | China |
|
|
| NA | China |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Plant Diversity and Evolution
Introduction
Amphisphaeriales was described by Eriksson and Hawksworth in 1986 and is phylogenetically closely related to Xylariales within the Xylariomycetidae group (Senanayake et al. 2015; Samarakoon et al. 2022). The type genus Amphisphaeria was established by Cesati et al. in 1863 for the family Amphisphaeriaceae, and two additional genera, Griphosphaerioma and Lepteutypa, were later added to the family (Wijayawardene et al. 2018). However, Lepteutypa was ultimately synonymized with Amphisphaeria based on holomorphic morphology and multigene phylogeny by Samarakoon et al. (2020), while Griphosphaerioma was regarded as a synonym of Labridella by Rossman et al. (2016). Currently, Amphisphaeriaceae includes Amphisphaeria and Labridella (Hyde et al. 2024a).
Amphisphaeria, typified by A. umbrina, generally comprises immersed, clypeate, and ostiolate ascomata that appear as elevated, blackened, round spots on the host surface (Samarakoon et al. 2020). The peridium layers comprise a hyaline inner layer and a brown outer layer of cells. Paraphyses are filamentous, septate, and flexuous (Wang et al. 2004). The cylindrical asci are 8-spored, with J+ or J-, discoid, tubular, or wedge-shaped apical rings, and the ascospores are 1–3 septate, ellipsoidal, and brown (Cesati and De 1863; Wang et al. 2004; Samarakoon et al. 2019, 2020, Samarakoon 2023). Both coelomycetous and hyphomycetous asexual morphs are reported in Amphisphaeria (Samarakoon et al. 2020). Saprobic Amphisphaeria species have been reported from woody branches and various monocotyledon hosts, including grasses from different geographical regions (Samarakoon et al. 2019). Members of Amphisphaeria are predominantly found on dead plant materials in both terrestrial and marine habitats (Senanayake et al. 2015, 2019; Samarakoon et al. 2019; Sun et al. 2025). Given the widespread nature and diverse lifestyles of Amphisphaeria species, further taxonomic studies are essential.
In this study, we examine the taxonomy, lifestyle, ecological roles, and distribution of Amphisphaeria species in northern Thailand. We introduce six species of Amphisphaeria, along with a new host record. These taxa were isolated from dead twigs of Mesua sp., Mimusops elengi, Paraserianthes lophantha, Pterocarpus sp., Schima wallichii, Senna siamea, and an undetermined host, from two provinces in northern Thailand. Morphological illustrations of the taxa are provided. Phylogenetic studies incorporating combined nuclear ribosomal large subunit rDNA (LSU) and the internal transcribed spacer (ITS) region confirm the taxonomic placements of these species as novel within Amphisphaeria and also support the finding of a new host record of an existing Amphisphaeria taxon.
Materials and methods
Sample collection, isolation, and morphology
Dead twigs were collected from the premises of the Mushroom Research Center (MRC) and Mae Fah Luang University during the cold (October, November) and wet (July) seasons in northern Thailand. After the collection details were recorded (Rathnayaka et al. 2025), specimens were brought to the laboratory in plastic bags and stored in a paper envelope. A Motic SMZ 168 Series stereo microscope (Leica Microsystems Company, Germany) was used to observe the fungi colonizing the host, and water-mounted slides were prepared to examine their micro-morphological characters. Melzer’s reagent and Indian ink were utilized for further morphological investigations. Digital images of the micro morphological features were captured using a Canon 750D camera (Canon, Tokyo, Japan) mounted on a Nikon ECLIPSE E600 compound microscope (Nikon, Tokyo, Japan), with objective lenses providing magnifications of 10×, 20×, 40×, 60×, and 100×. Photo plates were produced using Adobe Photoshop CS6 software (Adobe Systems, USA). Measurements of the fungal characteristics were conducted using the Tarosoft® Image Framework software (version 0.9.7).
Single spore isolations were performed as outlined by Senanayake et al. (2020) to obtain pure cultures. Germinated spores were identified after 24 hours of growth on malt extract agar (MEA), then transferred to fresh MEA media and incubated at 25 °C. Pure cultures were maintained on malt extract agar (MEA) at 25 °C, and their cultural characteristics were observed after one month of growth.
Herbarium materials and cultures were deposited in the Mae Fah Luang University Herbarium (MFLU) and Mae Fah Luang University Culture Collection (MFLUCC), respectively. Index Fungorum and faces of fungi numbers were obtained (Jayasiri et al. 2015; Index Fungorum 2025). The species descriptions were added to the Greater Mekong Subregion database (https://gmsmicrofungi.org/) (Chaiwan et al. 2021) and the Fungalpedia webpage (Hyde et al. 2023).
DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from fresh mycelia grown on MEA for 15 days, or DNA was extracted directly from the fruiting bodies using a DNA Extraction Kit (Omega Biotek) following the manufacturer’s protocol. The polymerase chain reaction (PCR) was conducted in a total volume of 25 μL, comprising 12.5 μL of 2× Power TaqPCR Master Mix, 1 μL of each primer (20 M), 2 μL of genomic DNA, and 8.5 μL of distilled water. PCR was performed using an Eppendorf thermal cycler (Mastercycler X50s) to amplify the LSU and ITS loci under the conditions outlined in Table 1. Agarose gel electrophoresis was performed to assess the quality of PCR products prior to sequencing at SolGent Co., South Korea.
The PCR thermal cycling protocol consists of an initial step at 95 °C for 5 minutes, followed by a final elongation step at 72 °C for 10 minutes, and concluding with a hold at 4 °C, as well as annealing at 55 °C.
Phylogenetic analyses
SeqMan (DNAStar, Inc., Madison, WI, USA) was used to generate consensus sequences from the forward and reverse chromatograms obtained. The sequences were subsequently subjected to a BLASTn search in NCBI (https://blast.ncbi.nlm.nih.gov/). The LSU and ITS sequences for Amphisphaeria species were retrieved from the GenBank database (Table 2). Each gene locus was aligned using the default settings in MAFFT v. 7 (https://mafft.cbrc.jp/alignment/server/) (Katoh et al. 2019) and trimmed with trimAl v. 1.2 (Capella-Gutiérrez et al. 2009). Single locus alignments were concatenated using BioEdit v. 7.0.5.2 (Hall 1999). ALTER (http://www.sing-group.org/ALTER/) was used to convert FASTA files into PHYLIP format. Single-locus and multi-locus aligned datasets were analyzed separately using maximum likelihood (ML) and Bayesian inference (BI). Maximum likelihood analysis was performed using IQ webserver (http://iqtree.cibiv.univie.ac.at/) with bootstrap support for 1000 replicates (Nguyen et al. 2015).
MrModeltest v. 2.2 was used to estimate the evolution model using the Akaike information criterion (AIC), implemented in PAUP v. 4.0b10 (Nylander 2004). MrBayes v. 3.1.2 was used to conduct Bayesian inference (BI) analyses for estimating posterior probabilities (PP) through Markov chain Monte Carlo sampling (MCMC) under the GTR+I+G model (Huelsenbeck et al. 2001; Ronquist and Huelsenbeck 2003). Markov chains were executed for 1,000,000 generations, with trees sampled every 100^th^ generation. The initial 25% of trees were discarded during the burn-in phase, while the remaining trees were used to compute the posterior probability (PP) in the majority rule consensus tree. FigTree v.1.4 was employed to visualize the phylograms (Rambaut and Drummond 2012), which were subsequently edited using Microsoft PowerPoint.
Abbreviations: CBS: Westerdijk Fungal Biodiversity Institute, Utrecht, the Netherlands; CFCC: China Forestry Culture Collection Center, Research Institute of Forest Ecology, Environment and Protection, Beijing, China; CGMCC: China General Microbiological Culture Collection Center, Beijing, China; CMUB: Chiang Mai University, Chiang Mai, Thailand; GZAAS and GZCC: Guizhou Academy of Agricultural Sciences, Guizhou, China; HKAS: Herbarium of Cryptogams Kunming Institute of Botany Academia Sinica, China; HKUCC: University of Hong Kong Culture Collection, Department of Ecology and Biodiversity, Hong Kong, China; KUMCC: Kunming Institute of Botany Culture Collection, China; KoLRI: Kholodny Institute of Botany, Tereshchenkivska, Kiev, Ukraine; MFLU, MFLUCC: Mae Fah Luang University, Chiang Rai, Thailand; PRA: The Herbarium of the Institute of Botany, Czech Academy of Sciences, Průhonice, Czech Republic; UESTCC: University of Electronic Science and Technology culture Collection, Xiyuan, Chengdu, China; WU: The Herbarium of the University of Vienna, Austria. Type species are denoted in ‘*’; “N/A” indicates the sequences are not available.
Results
Sequence alignment and phylogenetic analyses
The combined LSU and ITS sequence matrix comprised 74 Amphisphaeriaceae taxa, including our ten new strains. The tree is rooted with Beltrania rhombica (CBS 123.58), Beltraniella endiandrae (CBS 137976), and Beltraniopsis longiconidiophora (MFLUCC 17-2139). The combined alignment comprised 1976 characters, including gaps (LSU: 1–1,332 and ITS: 1,333–1,970). Both ML and BI analyses yielded trees with similar topologies.
The ML phylogram was used as the backbone tree (Fig. 1). The best-scoring RAxML tree had an optimization likelihood value of -14759.462. The matrix contained 1,000 distinct patterns, with 29.25% of the characters being undetermined or gaps. Estimated base frequencies were: A = 0.250, C = 0.250, G = 0.250, T = 0.250; substitution rates AC = 0.72949, AG = 2.67549, AT = 1.00000, CG = 0.72949, CT = 3.60004, GT = 1.0; the gamma distribution shape parameter was 0.467, Tree Length: 2.512. For the Bayesian analysis, the best-fit models generated from MrModeltest under the Akaike information criterion (AIC) are as follows: LSU: TI M3+I+G and ITS: TVM+I+G.
Phylogram generated from ML analysis based on combined LSU, ITS sequence data. Bootstrap support values for ML ≥75% and PP ≥0.90 are indicated near the corresponding nodes. The tree is rooted with Beltrania rhombica (CBS123.58), Beltraniella endiandrae (CBS:137976), and Beltraniopsis longiconidiophora (MFLUCC17-2139). Type and reference strains are in bold, while the novel strains are in bold red.
Amphisphaeria mesuae (MFLUCC 25-0197) clustered with Amphisphaeria hibiscicola (HKAS 136910) and Amphisphaeria umbrina (PRA JV24328). Amphisphaeria micheliae (MFLU 25-0077) clustered in a clade shared by other A. micheliae strains (HKAS 107012, MFLU 20-0172, MFLUCC 21-0207, MFLUCC 24-0324, UESTCC 23-0123, UESTCC 23-0124, UESTCC 23-0125). Amphisphaeria mimusopis (MFLU 25-0076, MFLU 25-0159) was sister to A. flava (MFLU 18-0102). Amphisphaeria paraserianthis (MFLU 25-0075) formed a separate lineage, basal to A. neoaquatica (MFLUCC 14-0045) and A. hongheensis (GMB1135). Amphisphaeria pseudomicheliae was a sister to the A. micheliae group. Amphisphaeria pterocarpi (MFLU 25-0073, MFLUCC 25-0195) formed a separate lineage, sister to A. curvaticonidia (MFLUCC 18-0620, HKAS 102288). Amphisphaeria schimae (MFLU 25-0071, MFLUCC 25-0196) is sister to A. ailaoshanensis (KUNCC 23-15520, KUNCC 23-15521).
Taxonomy
Amphisphaeria
mesuae
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
Z.L. Tun & K.D. Hyde sp. nov.
45E59076-317D-5F4D-AB3B-FFD94A4465BE
Index Fungorum: IF903423
Facesoffungi Number: FoF17659
Etymology.
The epithet refers to the host genus, Mesua, from which the fungus was isolated.
Amphisphaeria mesuae (MFLU 25-0069, holotype). A. Decaying woody twig; B. Appearance of an ascoma on the host; C, D. Vertical section through an ascoma; E. Vertical section of peridium; F. Paraphyses; G. J+ Apical ring; H, I. Asci; J–M. Ascospores; N. Upper view of culture; O. Reverse view of culture. Scale bars: 200 μm (B, C); 100 μm (D); 20 μm (E); 5 μm (F, G); 40 μm (H, I); 10 μm (J–M).
Holotype.
MFLU 25-0069.
Description.
Saprobic on decaying twigs of Mesua sp. Sexual morph: Ascomata 520–580 μm high, 260–290 µm wide (xˉ = 549 × 272 µm, n = 5), immersed, visible as black spots with tiny pores, flat, scattered or aggregated, globose to subglobose, ostiolate. Peridium 14–18 µm (xˉ = 15 µm, n = 5), two-layered; outer layer wide, comprising thick-walled, dark brown cells of textura angularis, inner layer thin, composed of hyaline textura angularis cells. Paraphyses 3–5 µm wide, hyaline, septate, longer than asci, narrow towards the apex. Asci 86–134 × 7–13 µm (xˉ = 115 × 9 µm, n = 20), 8-spored, unitunicate, cylindrical, with short pedicel, apically rounded, with a J+, apical ring. Ascospores 14–18 × 5–6.8 µm (xˉ = 15 × 6 µm, n = 20), uniseriate, ellipsoidal, hyaline when immature, turning sub-hyaline to pale brown at maturity, 1-septate, slightly constricted at the septum, guttulate, smooth-walled, lacking a sheath in Indian ink. Asexual morph: Not observed.
Culture characteristics.
Colonies on MEA, reaching 4 cm diam. after 15 days at 27 °C, from above white to pale yellow radiating outwards, dense, circular to slightly irregular, flattened with smooth surface, with smooth margin; reverse pale brown in the middle, yellowish brown at the margin.
Material examined.
Thailand • Chiang Rai Province, Mae Fah Luang University (20°02′42″N, 99°53′41″E), on decaying dead twigs of Mesua sp. (Calophyllaceae), 02 October 2023, Zaw Lin Tun P11 (holotype MFLU 25-0069); ex-type culture MFLUCC 25-0197.
Notes.
Phylogenetic analyses revealed that Amphisphaeria mesuae (MFLUCC 25-0197) clustered with Amphisphaeria hibiscicola (HKAS 136910) and Amphisphaeria umbrina (PRA JV24328) in (Fig. 1). The interspecies genetic distances between A. mesuae (MFLU25- 0069) and A. hibiscicola (HKAS 136910) showed the following base pair differences (without gaps): 5.7% for LSU (49/935 bp) and 11.18% for ITS (66/559 bp). Amphisphaeria mesuae (MFLUCC 25-0197) and Amphisphaeria umbrina (PRA JV24328) showed the following base pair differences (without gaps): 3.3% for LSU (28/844 bp) and 9.45% for ITS (54/571 bp). Morphologically, A. mesuae differs from A. hibiscicola by having taller ascomata (520–580 μm) compared to those of A. hibiscicola (212–450 μm), whereas A. hibiscicola has broader ascomata (456–570 μm wide) than A. mesuae (260–290 μm) (Sun et al. 2025). The asci of A. mesuae (86–134 × 7–13 µm) are smaller than those of A. hibiscicola (122–152 × 6–8 µm) (Sun et al. 2025). The ascospores of A. mesuae (14–18 × 5–6.8 µm) are ellipsoidal, while the ascospores of A. hibiscicola (8–14 × 3.5–5 μm) are fusiform (Sun et al. 2025). Amphisphaeria hibiscicola has a gelatinous sheath, whereas A. mesuae lacks a sheath (Sun et al. 2025
Morphologically, A. mesuae differs from A. umbrina by having higher ascomata (520–580 μm vs. 400–480 μm) but narrower width (260–290 μm vs. 560–640 μm) (Wang et al. 2004).The asci of A. mesuae (122–152 × 6–8 µm) are also smaller than those of A. umbrina (150–170 × 11–13 µm) (Wang et al. 2004). Similarly, the ascospores of A. mesuae (14–18 × 11–12 µm) are smaller than those of A. umbrina (18–22 × 6–8 μm) (Wang et al. 2004).
Based on the distinct morphology and phylogenetic evidence, along with the recommendations for species delineation proposed by Chethana et al. (2021) and Maharachchikumbura et al. (2021), we introduce A. mesuae as a new species.
Amphisphaeria
micheliae
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
Samarak., Jian K. Liu & K.D. Hyde, 2020
5A9ED485-F938-5CFD-9E3D-2586DC1E0AF2
MycoBank No: 836112
Facesoffungi Number: FoF08752
Description.
Saprobic on dead twigs of Senna siamea. Sexual morph: Ascomata 308–320 µm high, 348–360 µm wide, (xˉ = 312 × 353 µm, n = 5), immersed, visible as black spots in light-coloured areas on the host, solitary, scattered, subglobose to oblate, papillate. Ostiole 100–106 µm high, 65–70 µm diam (xˉ = 102 × 67 µm, n = 5), centric. Peridium 45–58 µm (xˉ = 48 µm, n = 5) two-layered; outer layer thick, dense, made up of red-dish-brown cells of textura angularis; inner layer thin, comprising hyaline cells. Paraphyses 3–5 µm wide (xˉ = 4.4 µm, n = 5), hyaline, longer than asci, septate, guttulate, embedded in a gelatinous matrix. Asci 85–115 × 6.5–7 µm (xˉ = 101 × 6.8 µm, n = 20), 8-spored, unitunicate, cylindrical, thin-walled, short-pedicellate, apically rounded, with a J+, discoid apical ring. Ascospores 15–16 × 5–6 µm (xˉ = 15.5 × 5.9 µm, n = 20), uniseriate, oblong or narrowly fusiform, guttulate, hyaline when immature, turning sub-hyaline to olivaceous grey, 1-septate, slightly constricted at the septum, straight to slightly curved, smooth-walled, lacking a sheath in Indian ink. Asexual morph: Not observed.
Amphisphaeria micheliae (MFLU 25-0077). A. Senna siamea tree; B. Appearance of ascomata on host (arrows indicate ascomata visible as black spots); C. Vertical section through an ascoma; D. Vertical section of ascomatal wall; E. Paraphyses; F–H. Asci; I. J+ Apical ring; J–L. Ascospores. Scale bars: 200 μm (C); 100 μm (D); 20 μm (E); 5 μm (F, G); 50 μm (H, I); 10 μm (J–L).
Material examined.
Thailand • Chiang Rai Province, Mae Fah Luang University premises (20°02′42″N 99°53′41″E), on dead decaying twigs of Senna siamea (Fabaceae), 02 October 2024, Zaw Lin Tun T7, (MFLU 25-0077).
Notes.
Based on the phylogenetic analyses, our strain (MFLU 25-0077) clustered with Amphisphaeria micheliae group (Fig. 1). Morphologically, our strain (MFLU25-0077) resembles the type of A. micheliae (MFLU 20-0172) (Samarakoon et al. 2020). The asci length of our collection (MFLU25-0077) measures 85–115 × 6.5–7 µm, while those of the type of A. micheliae (HKAS 107012) range from 92–135 × 7–10.5 µm (Samarakoon et al. 2020). The ascospore length of A. micheliae (MFLU 25-0077 and HKAS 107012) is also similar (15–16 × 5–6 µm vs. 15.5–21 × 6–7.5 µm) (Samarakoon et al. 2020). The comparison of inter-species genetic distances between our strain (MFLU25-0077) and HKAS 107012 reveals largely similar base pair differences in the LSU (99%) and ITS (99%) regions. Based on this morpho-phylogenetic evidence, we identify our isolate as A. micheliae. Previously, A. micheliae was isolated from dead twigs of Acer truncatum, Alstonia scholaris, Micromelum integerrimum, and Michelia alba in China and Thailand (Samarakoon et al. 2020; De Silva et al. 2022; Li et al. 2024; Pathirana et al. 2025). This is the first report of A. micheliae from Senna siamea in Thailand.
Amphisphaeria
mimusopis
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
Z.L. Tun & K.D. Hyde sp. nov.
CED7A44E-2ADA-5329-A2E4-BE2F6DCA0AD8
Index Fungorum: IF903593
Facesoffungi Number: FoF17660
Etymology.
The epithet refers to the host genus, Mimusops, from which the fungus was isolated.
Amphisphaeria mimusopis (MFLU 25-0076, holotype). A. Decaying dead branch; B. Appearance of ascomata on host (arrows indicate the ostiolar openings); C, D. Vertical section through an ascoma; E. Vertical section of ascomatal wall; F. Paraphyses; G. J+ Apical ring; H, I. Asci; J–O. Ascospores. Scale bars: 500 μm (C); 200 μm (D); 10 μm (E); 5 μm (F, G); 50 μm (H, I); 20 μm (J–O).
Holotype.
MFLU 25-0076.
Description.
Saprobic on the decaying branch of Mimusops elengi. Sexual morph: Ascomata 286–370 μm high, 364–455 µm wide, (xˉ = 321 × 397 µm, n = 5), immersed, visible as black spots with tiny pores, staining the host surface pale reddish brown around the ostioles, solitary, scattered to aggregated, globose to sub-globose. Ostiole central, prominent, 150–166 μm high, 82–84 µm wide (xˉ = 154 × 82 µm, n = 5). Peridium 12–25 µm (xˉ = 20 µm, n = 5), two-layered; outer layer thick, dense, reddish-brown cells of textura angularis, inner layer thin, cells hyaline, of textura angularis. Paraphyses 3–6 µm wide (xˉ = 4.2 µm, n = 5), hyaline, septate, longer than asci and embedded in a gelatinous matrix. Asci 126–164 × 8–13 µm (xˉ = 145.7 × 10. 2 µm, n = 20), 8-spored, unitunicate, cylindrical, with short pedicel, apically rounded, with a J+, discoid, apical ring. Ascospores 18–22 × 5–7 µm (xˉ = 20 × 6 µm, n = 20), uniseriate, cylindrical to oblong, hyaline when immature, turning subhyaline to brown at maturity, 1-septate, slightly constricted at the septum, guttulate, smooth-walled, lacking a sheath in Indian ink. Asexual morph: Not observed.
Material examined.
Thailand • Chiang Rai Province, Mae Fah Luang University (20°02′42″N, 99°53′41″E), on the dead, decaying twigs of Mimusops elengi (Sapotaceae), 2 October 2023, ZL Tun P3 (holotype MFLU 25-0076).
Additional specimens examined.
Thailand • Chiang Rai Province, Mae Fah Luang University (20°02′42″N, 99°53′41″E), on the dead decaying branch of Mimusops elengi (Sapotaceae), 2 October 2023, ZL Tun P3A (MFLU 25-0159).
Notes.
Amphisphaeria mimusopis is sister to A. flava (MFLU 18-0102) with 100% ML and 1.00 PP bootstrap support (Fig. 1). The interspecies genetic distances between A. mimusopis (MFLU 25-0076) and A. flava (MFLU 18-0102) showed the following base pair differences (without gaps): 1.4% across LSU (13/892 bp) and 6.7% across ITS (37/530 bp). Amphisphaeria mimusopis can be distinguished from A. flava by having larger ascospores (18–22 × 5–7 µm vs 13–16 × 5–7μm) (Samarakoon et al. 2019). Amphisphaeria mimusopis has narrower paraphyses (3–6 µm) than A. flava (7–16.3 µm) (Samarakoon et al. 2019). The ascomata of A. mimusopis produce a pale reddish-brown pigment surrounding the ostioles, whereas A. flava results in a pale-yellow pigmentation on the surface (Samarakoon et al. 2019). Based on morphological and phylogenetic evidence, as well as the recommendations for species delineation by Chethana et al. (2021), we establish Amphisphaeria mimusopis as a new species.
Amphisphaeria
paraserianthis
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
Z.L. Tun & K.D. Hyde sp. nov.
80565701-C43E-5DF1-AFAF-02E95634C9B0
Index Fungorum: IF903724
Facesoffungi Number: FoF17661
Etymology.
The epithet refers to the host genus, Paraserianthes, from which the fungus was isolated.
Amphisphaeria paraserianthis (MFLU 25-0075, holotype). A. Dead twigs of Paraserianthes lophantha; B. Appearance of ascomata on the host surface (arrow indicates the appearance of ascomata on the host surface); C. Vertical section of an ascoma; D. Ostiole; E. Peridium; F. Paraphyses; G–I. Asci; J. J+, apical ring turning blue in Melzer’s reagent; K–O. Immature and mature ascospores. Scale bars: 200 μm (C); 50 μm (D); 20 μm (E); 5 μm (F, J); 50 μm (G–I); 10 μm (J–O).
Holotype.
MFLU 25-0075.
Description.
Saprobic on decaying twigs of Paraserianthes lophantha. Sexual morph: Ascomata 430–520 µm wide, 290–400 µm high (xˉ = 497 × 355 µm, n = 5), immersed, solitary or grouped, scattered, globose to subglobose, dark brown to black, ostiolate. Ostiole central, comprising a short papilla, with an ostiolar canal lined with hyaline periphyses. Peridium 16–20 µm (xˉ = 16 µm, n = 5) two-layered; outer layer wide, dark brown, thick-walled cells of textura angularis, inner layer comprising one layer of pale yellow or sub-hyaline to hyaline cells of textura angularis, thin-walled. Paraphyses 3–5 µm wide (xˉ = 4.5 µm, n = 5), hyaline, filiform, septate, guttulate, longer than asci. Asci 85–152 × 7.5–16 µm (xˉ = 125.5 × 10.5 µm, n = 20), 8-spored, unitunicate, cylindrical, thin-walled, short-pedicellate, apically rounded, with a J+, conspicuous, discoid, apical ring. Ascospores 16–20 × 5–9 µm (xˉ = 19 × 6.9 µm, n = 20), uniseriate, oblong to ellipsoid, hyaline when young, turning yellow brown at maturity, 3-septate, rounded to obtuse ends, smooth-walled, guttulate, without a sheath. Asexual morph: Not observed.
Material examined.
Thailand • Chiang Mai Province, in the forests around the Mushroom Research Center (19°07.200'N, 98°44.044'E), on fallen decaying twigs of Paraserianthes lophantha (Fabaceae), 14 November 2022, Zaw Lin Tun M14, (holotype MFLU 25-0075).
Notes.
Amphisphaeria paraserianthis (MFLU 25-0075) formed a distinct lineage that is basal to A. neoaquatica (MFLUCC 14-0045) and A. hongheensis (GMB1135) (Fig. 1). The interspecies genetic distances between A. paraserianthis and A. neoaquatica (MFLUCC 14-0045) showed the following base pair differences (without gaps): 7% for LSU (58/818 bp) and 5.11% for ITS (25/489 bp). Amphisphaeria paraserianthis and A. hongheensis (GMB1135) showed the following base pair differences (without gaps): 2.8% for LSU (26/923 bp) and 7.7% for ITS (35/455 bp). The ascomata of A. paraserianthis (430–520 × 290–400 µm) are larger than those of A. neoaquatica (250–320 × 300–330 μm) and smaller than those of A. hongheensis (430–750 × 360–640 μm). The asci of A. paraserianthis (85–152 × 7.5–16 µm) are larger than those of A. neoaquatica (126–138 µm × 8–10 μm), and smaller than A. hongheensis (175−265 × 10–15 µm) (Luo et al. 2019, Liu et al. 2024a). The ascospores of A. paraserianthis (16–20 × 5–9 µm) are larger than A. neoaquatica (15–17 × 5–7 μm) and smaller than A. hongheensis (20−40 × 5−11μm) µm (Luo et al. 2019, Liu et al. 2024a). In addition, A. paraserianthis possesses 3-septate ascospores while A. neoaquatica and A. hongheensis possess 1-septate ascospores (Luo et al. 2019, Liu et al. 2024a). Based on the species delineation suggestions made by Chethana et al. (2021) and the findings from morpho-phylogenetic analyses, we establish A. paraserianthis (MFLU 25-0075) as a new species.
Amphisphaeria
pseudomicheliae
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
Z. L. Tun & K.D. Hyde sp. nov.
D78D68D1-A966-5F25-9819-7D7824AA83D1
Index Fungorum: IF903743
Facesoffungi Number: FoF17662
Etymology.
Refers to the morphological similarity with Amphisphaeria micheliae.
Amphisphaeria pseudomicheliae (MFLU 25-0077, holotype). A. Host; B. Appearance of ascomata on host (arrows indicate ascomata as tiny pores on host surface); C. Vertical section through an ascoma; D. Vertical section of peridium; E. Paraphyses; F. An inconspicuous J+ apical ring; G–I. Asci; J–Q. Ascospores; Q. An ascospore with a thin gelatinous sheath (in Indian ink). Scale bars: 100 μm (C); 10 μm (D–F); 20 μm (G–I); 10 μm (J–Q).
Holotype.
MFLU 25-0074.
Description.
Saprobic on decaying twigs. Sexual morph: Ascomata 202–273 μm high, 349–400 µm diam (xˉ = 246 × 337 µm, n = 5), immersed, visible as tiny pores, flat, solitary to aggregated, scattered, globose to subglobose, brown, ostiolate. Peridium 17–23 µm wide (xˉ = 20.6 µm, n = 5), two-layered; outer layer wide, dark brown, comprising thick-walled cells of textura angularis, inner layer comprising hyaline cells of textura angularis, thin-walled. Paraphyses 4–4.4 µm (xˉ = 4.2 µm, n = 5) wide, hyaline, longer than asci, filiform, guttulate, septate, embedded in a gelatinous matrix. Asci 84–97× 7–9 µm (xˉ = 89.8 × 8.45 µm, n = 20), 8-spored, unitunicate, cylindrical, with short pedicel, apically rounded and narrowed, with a J+ inconspicuous apical ring. Ascospores 11–17 × 4–6 µm (xˉ = 15 × 5 µm, n = 20), uniseriate, fusiform, hyaline when immature, turning yellow to yellowish brown on maturity, 1-septate, guttulate, smooth-walled, slightly constricted at septum, straight to slightly curved, surrounded by a thin mucilaginous sheath. Asexual morph: Not observed.
Material examined.
Thailand • Chiang Rai Province, Mae Fah Luang University premises (20°02′42″N, 99°53′41″E), on decaying dead twigs of an undetermined host, 06 July 2023, Zaw Lin Tun E15 (holotype MFLU 25-0074).
Notes.
Amphisphaeria pseudomicheliae is closely related to A. micheliae, receiving robust support with 97% ML and 1 PP bootstrap support (Fig. 1). However, it can be distinguished from A. micheliae by its larger ascomata (202–273 × 349–400 µm vs. 180–210 × 225–370 µm) and smaller asci (84–97 × 7–9 µm vs. 92–135 × 7–10.5 μm) (Samarakoon et al. 2020). Additionally, the ascospores of A. pseudomicheliae are smaller (11–17 × 4–6 µm) than those of A. micheliae (15.5–21 × 6–7.5 μm) (Samarakoon et al. 2020). Amphisphaeria pseudomicheliae has a thin mucilaginous sheath, whereas A. micheliae lacks (Samarakoon et al. 2020). The genetic analysis reveals interspecies distances of 4.3% base pair differences (without gaps) in the ITS (23/524 bp) and 0.43% in the LSU (4/873 bp) between A. pseudomicheliae (MFLU25-0074) and A. micheliae (HKAS 107012). Amphisphaeria micheliae is uncertain since our new species lack protein coding genes. Thus, numerous collections with the protein coding genes can provide better resolution between the species. Amphisphaeria pseudomicheliae is described as a new species based on its distinct morphology and phylogenetic data.
Amphisphaeria
pterocarpi
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
Z.L. Tun & K.D. Hyde sp. nov.
A235A1BC-E8B3-51B8-BC50-CF2EE7E4E8A9
Index Fungorum: IF903750
Facesoffungi Number: FoF17663
Etymology.
The epithet refers to the host genus Pterocarpus, from which the fungus was isolated.
Amphisphaeria pterocarpi (MFLU 25-0073, holotype). A. Dead branches; B. Appearance of ascomata on host (arrows indicate ascomata as spots on host surface); C, D. Vertical sections through ascomata; E. Vertical section of peridium; F. Paraphyses; G, H. Asci; I. J+ apical ring; J–L. Ascospores; M. Germinated ascospore; N. Upper view of culture; O. Reverse view of culture. Scale bars: 100 μm (C, D); 10 μm (E); 5 μm (F, G); 50 μm (H, I); 20 μm (J–M).
Holotype.
MFLU 25-0073.
Description.
Saprobic on recently dead branches of Pterocarpus sp. Sexual morph. Ascomata 225–237 μm high, 355–373 µm wide, (xˉ = 231 × 360 µm, n = 5), immersed, visible as black spots, host tissue becoming reddish around the apical pores, solitary to aggregated, scattered, globose to sub-globose, brown. Ostiole central, 70–60 wide, comprising a short papilla. Peridium 12–16 µm wide (xˉ = 14 µm, n = 5), two-layered; outer layer wide, dark brown, thick-walled cells of textura angularis, inner layer comprising one layer of hyaline cells of textura angularis, thin-walled. Paraphyses 4–6 µm wide (xˉ = 4.8 µm, n = 5), hyaline, septate, guttulate, embedded in a gelatinous matrix. Asci 95–116 × 9–14 µm (xˉ = 106 × 11 µm, n = 20), 8-spored, unitunicate, cylindrical, deliquescing, with short pedicel, apically rounded, with a J+, wedge-shaped, apical ring. Ascospores 19–21 × 5–7 µm (xˉ = 20 × 6 µm, n = 20), ellipsoidal, hyaline when immature, turning yellow to yellowish-brown when mature, 3-septate, guttulate, smooth-walled, surrounded by a mucilaginous sheath. Asexual morph: Not observed.
Culture characteristics.
Colonies on MEA reaching 4 cm diam. after 15 days at 27 °C, from above white, dense, irregular, flattened with smooth surface, with lobate margin; reverse yellow red in the middle, yellow at the margin.
Material examined.
Thailand • Chiang Rai Province, Mae Fah Luang University premises (20°02′42″N, 99°53′41″E), on recently dead branches of Pterocarpus rotundifolius (Fabaceae), 06 November 2023, Zaw Lin Tun H62 (holotype MFLU 25-0073), ex-type culture MFLUCC 25-0195.
Additional specimens examined.
Thailand • Chiang Rai Province, Mae Fah Luang University premises (20°02′42″N, 99°53′41″E), on dead branches of Pterocarpus rotundifolius (Fabaceae), 06 November 2023, Zaw Lin Tun 2H62 (MFLU 25-0072).
Notes.
Based on our phylogenetic analyses, A. pterocarpi formed a separate lineage, sister to A. curvaticonidia (MFLUCC 18-0620, HKAS 102288), with 100% ML and 1.00 PP bootstrap support (Fig. 1). Amphisphaeria pterocarpi can be distinguished from A. curvaticonidia by its smaller ascomata (225–237 × 355–373 µm vs. 320–390 × 360–410 µm), asci (95–116 × 9–14 µm vs. 121–162 × 10.5–17.5 µm), and ascospores (19–21 × 5–7 µm vs. 17–23 × 6–9 µm) (Samarakoon et al. 2020). Additionally, the ascospores of A. pterocarpi are ellipsoidal, whereas A. curvaticonidia have oblong or narrowly fusiform ascospores. The asexual morph of A. curvaticonidia has been documented as coelomycetous in culture, while the asexual morph of A. pterocarpi has yet to be observed in culture (Samarakoon et al. 2020). When considering the base pair differences (without gaps) between A. pterocarpi and A. curvaticonidia (MFLUCC 18-0620, HKAS 102288), 2.5% base pair differences (without gaps) were revealed in LSU (27/1040 bp) and 3.30% base pair differences (without gaps) in ITS (18/545 bp). Due to the distinct morphology and phylogenetic evidence, along with the species delineation guidelines provided by Chethana et al. (2021), we introduce A. pterocarpi as a new species.
Amphisphaeria
schimae
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
Z.L. Tun & K.D. Hyde sp. nov.
18EC6EC8-7863-5BA8-ACB4-06C7E18BEE93
Index Fungorum: IF903771
Facesoffungi Number: FoF17664
Etymology.
The epithet refers to the host genus, Schima, from which the fungus was isolated.
Amphisphaeria schimae (MFLU 25-0070, holotype). A. Host; B. Close-up of ascoma on host (Arrow indicates ascoma visible as a black spot, with ostiole breaking through the host); C. Vertical section of an ascoma; D. Ostiole; E. Vertical section of peridium; F. Paraphyses; G–I. Asci (I in Melzer’s reagent); J–N. Ascospores with sheaths; O. Germinated ascospore; P. Upper view of culture; Q. Reverse view of culture. Scale bars: 100 μm (C); 20 μm (D); 10 μm (E, F); 50 μm (G–I); 20 μm (J–O).
Holotype.
MFLU 25-0070.
Description.
Saprobic on decaying twigs of Schima wallichii. Sexual morph: Ascomata 480–620 μm high × 520–683 µm wide, (xˉ = 531 × 583 µm, n = 5), immersed with ostiole breaking through host surface, visible as black spot, solitary to aggregated, scattered, globose to subglobose, brown, ostiolate. Ostiole central, comprising a short papilla, with an ostiolar canal. Peridium 13–16 µm wide (xˉ = 14.2 µm, n = 5), two-layered; outer layer thick, dense, reddish-brown cells of textura angularis, inner layer thin, hyaline cells of textura angularis. Paraphyses 3–5 µm wide, hyaline, filiform, septate, embedded in a gelatinous matrix. Asci 196–275 × 21–30 µm (xˉ = 236.2 × 22.9 µm, n = 20), 8-spored, unitunicate, cylindrical, with short pedicel, apically rounded, with J-, apical ring. Ascospores 36.9–40 × 13.5–15 µm (xˉ = 38.6 × 14 µm, n = 20), uniseriate, broadly fusiform, hyaline, turning olivaceous brown to brown at maturity, one median and constricted euseptum, with two distosepta, guttulate (especially at immaturity), broad to conically rounded at both ends, smooth, surrounded by a thick mucilaginous sheath. Asexual morph: Not observed.
Culture characteristics.
Colonies on MEA slow-growing, reaching 3 cm in diam. after 15 days at 27 °C, from above white to pale yellow radiating outwards, dense, circular, irregular, flattened with smooth surface, with lobate margin; reverse yellow brown in the middle, white at the margin.
Materials examined.
Thailand • Chiang Mai Province, in forests around the Mushroom Research Center (19°07.200'N, 98°44.044'E), on fallen dead twigs of Schima wallichii (Theaceae), 14 November 2022, Zaw Lin Tun C1 (holotype MFLU 25-0070), ex-type culture MFLUCC 25-0196.
Additional specimens examined.
Thailand • Chiang Mai Province, in forests around the Mushroom Research Center (19°07.200'N, 98°44.044'E), on fallen dead twigs of Schima wallichii (Theaceae), 14 November 2022, Zaw Lin Tun C2 (MFLU 25-0071).
Notes.
Based on our phylogenetic analyses, Amphisphaeria schimae (MFLU 25-0071, MFLUCC 25-0196) is sister to A. ailaoshanensis (KUNCC 23-15520, KUNCC 23-15521) (Fig. 1). However, A. schimae can be distinguished from A. ailaoshanensis by having larger ascomata (480–620 μm high × 520–683 μm vs. 100–140 μm high × 250–350 μm), asci (196–275 × 21–30 μm vs. 70–100 × 7–10 μm), and ascospores (36.9–40 × 13.5–15 μm vs. 14–20 × 5–8 μm) (Dissanayake et al. 2024). Additionally, the ascospores of A. schimae are broadly fusiform, hyaline, turning olivaceous brown to brown at maturity, with one median, conspicuously constricted euseptum, and surrounded by a thick mucilaginous sheath, whereas those of A. ailaoshanensis are fusiform, hyaline, guttulate, turning brown, 1–3-septate, and lack a mucilaginous sheath (Dissanayake et al. 2024). When comparing base pair differences (excluding gaps) between A. schimae (MFLUCC 25-0196) and A. ailaoshanensis (KUNCC 23-15520, KUNCC 23-15521), there is a 4.9% divergence in the LSU region (34/690 bp) and 8.2% divergence in the ITS region (43/525 bp). Based on the distinct morphological and phylogenetic evidence, along with the species delineation guidelines proposed by Chethana et al. (2021), we describe A. schimae as a new species.
Discussion
In this study, we introduce six new Amphisphaeria species, along with a new host and geographical record, based on morphological and multigene phylogenetic analyses of combined LSU and ITS alignments. This work significantly advances the taxonomy of Amphisphaeria by providing important insights into species delimitation, phylogenetic relationships, and morphological diversity. Moreover, the discovery of a new host and geographic record broadens our understanding of the ecological distribution and host specificity of these species, offering a more comprehensive framework for future taxonomic and ecological research in this group.
Most strains of Amphisphaeria lack protein gene sequences, with the exception of A. camelliae, A. flava, A. fuckelii, A. hongheensis, A. hydei, A. micheliae, A. parvispora, A. sambuci, A. thailandica, and A. uniseptata (Tusi et al. 2001; Jaklitsch et al. 2016; Voglmayr et al. 2019; Liu et al. 2019, 2024a; Samarakoon et al. 2019; Samarakoon 2023; Li et al. 2024). In this study, we encountered challenges in sequencing protein markers across our Amphisphaeria isolates. Specifically, all isolates lacked the β-tub and the rpb2 gene. These difficulties align with findings from previous studies, which also reported challenges in sequencing these protein markers for Amphisphaeria (Senanayake et al. 2019; Samarakoon et al. 2020; Zhang et al. 2023; Dissanayake et al. 2024; Sun et al. 2025).
To address these limitations, we constructed two phylogenetic trees: one using a combination of LSU, ITS, rpb2, and β-tub (not shown), and another using only LSU and ITS (Fig. 1). Comparison of the topologies of the two-locus (LSU and ITS) and four-locus trees showed mostly similar phylogenetic placements of the taxa. Since most of our species lacked β-tub and rpb2, we predominantly relied on LSU and ITS for our phylogenetic analyses. This approach allowed us to achieve robust phylogenetic resolution despite the absence of key protein markers in most isolates.
Currently, 312 names are listed under Amphisphaeria in Index Fungorum (July 2025). Wang et al. (2004) examined 170 type specimens and accepted only 12 species in Amphisphaeria, highlighting taxonomic confusion within this genus. The lack of molecular data for many Amphisphaeria species makes it difficult to confirm their taxonomic positions (Thiyagaraja et al. 2025). Additionally, the absence of protein gene sequences for Amphisphaeria species often leads to inaccurate identifications (Samarakoon 2023; Dissanayake et al. 2024). The high morphological similarity and lack of molecular data among Amphisphaeria species can cause misidentifications (Thiyagaraja et al. 2025). Therefore, incorporating new collections and multi-gene molecular data is essential for accurate species delimitation and a clearer understanding of species boundaries within Amphisphaeria.
The saprobes were collected from forest areas during both the wet and cold seasons of 2022–2024. The distribution of Amphisphaeria species is shown in Table 3. Amphisphaeria species are found worldwide, with reports from 15 countries. They occur on 24 host genera across 17 different plant families, with Sapindaceae being the most diverse (Table 3). Additionally, Amphisphaeria has been reported in other families, including two species each in Theaceae, Leguminosae, and Malvaceae, three in Fabaceae, and one each in Proteaceae, Gramineae, Calophyllaceae, Magnoliaceae, Actinidiaceae, Apocynaceae, Rutaceae, Oleaceae, Asparagaceae, Agavaceae, Rosaceae, and Euphorbiaceae (Table 3). Therefore, Malvaceae and Fabaceae are the second most diverse families for Amphisphaeria species (Table 3).
Amphisphaeria chiangmaiensis, A. curvaticonidia, A. flava, A. hydei, A. parvispora, A. micheliae, and A. thailandica have been previously documented in northern Thailand (Samarakoon et al. 2019–2023; Pathirana et al. 2025), highlighting the region as a hotspot for fungal diversity. In our study, we further contribute to the understanding of Amphisphaeria species in this region by describing six new species, each associated with different host families, as shown in Table 3. This discovery not only broadens the known taxonomic range of Amphisphaeria but also underscores the ecological adaptability of this genus to various plant hosts. Additionally, we report A. micheliae on Senna siamea for the first time in Thailand, marking an important addition to the fungal flora of the region. These findings collectively highlight the impressive diversity of Amphisphaeria species in northern Thailand and their associations with a wide array of host families, indicating a complex and dynamic ecological interaction that warrants further study. This genus also needs to be studied in other tropical regions to determine if it is equally diverse there (Hyde et al. 2024b).
Historically, there have been no records of Amphisphaeria species being identified as pathogens (Wang et al. 2004). However, recent studies have revealed new insights into the ecological roles of these fungi (Sun et al. 2025). For instance, A. hibiscicola was reported on diseased leaves of Hibiscus mutabilis (Sun et al. 2025). Additionally, A. orixae has been identified as an endophyte, highlighting its ability to colonize plant tissues without causing disease (Wang et al. 2023). These findings indicate that Amphisphaeria species may exhibit diverse ecological roles, ranging from pathogenicity to endophytism, and may have evolved from endophytic species (Bhunjun et al. 2024). Furthermore, while most Amphisphaeria species are typically found in terrestrial environments (Dissanayake et al. 2024), the discovery of Amphisphaeria aquatica in freshwater habitats on submerged, decaying wood in China and Thailand (Tsui et al. 2001; Luo et al. 2019) expands our understanding of their ecological adaptability. This contrast between terrestrial and aquatic habitats highlights the ecological versatility of the genus and raises questions about how environmental factors influence their behavior and role in ecosystems. Therefore, future research should concentrate on clarifying the complex ecological interactions between these fungi and their hosts, as well as examining the impact of environmental factors on their distribution and behavior. Furthermore, it is essential to enhance the understanding of the ecological and evolutionary importance of Amphisphaeria species. This acknowledges the ongoing significance of northern Thailand as a vital center for continued research, as emphasized by Hyde et al. (2018).
Supplementary Material
XML Treatment for Amphisphaeria mesuae
XML Treatment for Amphisphaeria micheliae
XML Treatment for Amphisphaeria mimusopis
XML Treatment for Amphisphaeria paraserianthis
XML Treatment for Amphisphaeria pseudomicheliae
XML Treatment for Amphisphaeria pterocarpi
XML Treatment for Amphisphaeria schimae
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barr ME Ramaley AW (1996) Two more species of Amphisphaeria (Xylariales).Mycotaxon 58: 349–351.
- 2Bhunjun CS Phukhamsakda C Hyde KD Mc Kenzie EHC Saxena RK Li Q (2024) Do all fungi have ancestors with endophytic lifestyles? Fungal Diversity 125: 73–98. 10.1007/s 13225-023-00516-5 · doi ↗
- 3Capella-Gutiérrez S Silla-Martínez JM Gabaldón T (2009) Trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses.Bioinformatics (Oxford, England)25(15): 1972–1973. 10.1093/bioinformatics/btp 34819505945 PMC 2712344 · doi ↗ · pubmed ↗
- 4Cesati V De NG (1863) Classification scheme of the Aschigerian Italic spherical sphericae: More or less belonging to the genus Sphaeria in the ancient meaning attributed to it by Persoon.Italian Cryptogamic Herbarium 1: 177–240.
- 5Chaiwan N Gomdola D Wang S Monkai J Tibpromma S Doilom M Wanasinghe D Mortimer P Lumyong S Hyde KD (2021) An online database providing updated information of microfungi in the Greater Mekong Subregion.Mycosphere: Journal of Fungal Biology 12(1): 1513–1526. 10.5943/mycosphere/12/1/19 · doi ↗
- 6Chang R Yan Z Jiang J Wang Y Si H Bose T Miao C (2025) Four novel endolichenic fungi from Usnea spp. (Lecanorales, Parmeliaceae) in Yunnan and Guizhou, China: Taxonomic description and preliminary assessment of bioactive potentials. Myco Keys 118: 55. 10.3897/mycokeys.118.122228 PMC 1214988740496201 · doi ↗ · pubmed ↗
- 7Chethana KT Manawasinghe IS Hurdeal VG Bhunjun CS Appadoo MA Gentekaki E RaspéO Itthayakorn P Hyde KD (2021) What are fungal species and how to delineate them? Fungal Diversity 109: 1–25. 10.1007/s 13225-021-00483-9 · doi ↗
- 8Crous PW Shivas RG Quaedvlieg Wvan der Bank M Zhang Y Summerell BA Guarro J Wingfield MJ Wood AR Alfenas AC Braun U Cano-Lira JF García D Marin-Felix Y Alvarado P Andrade JP Armengol J Assefa Aden Breeÿen A Camele I Cheewangkoon R De Souza JT Duong TA Esteve-Raventós F Fournier J Frisullo S García-Jiménez J Gardiennet A GenéJ Hernández-Restrepo M Hirooka Y Hospenthal DR King A Lechat C Lombard L Mang SM Marbach PAS Marincowitz S Marin-Felix Y Montaño-Mata NJ Moreno G Perez CAPérez Sierra AM Robertson JL Roux J Rubio E Schumacher RK Stchigel AM Sutton DA Tan YP Thompson EH Vanderlinde E Walker A · doi ↗ · pubmed ↗