Three new species of Mycena sect. Calodontes (Mycenaceae, Agaricales) from Northeastern China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
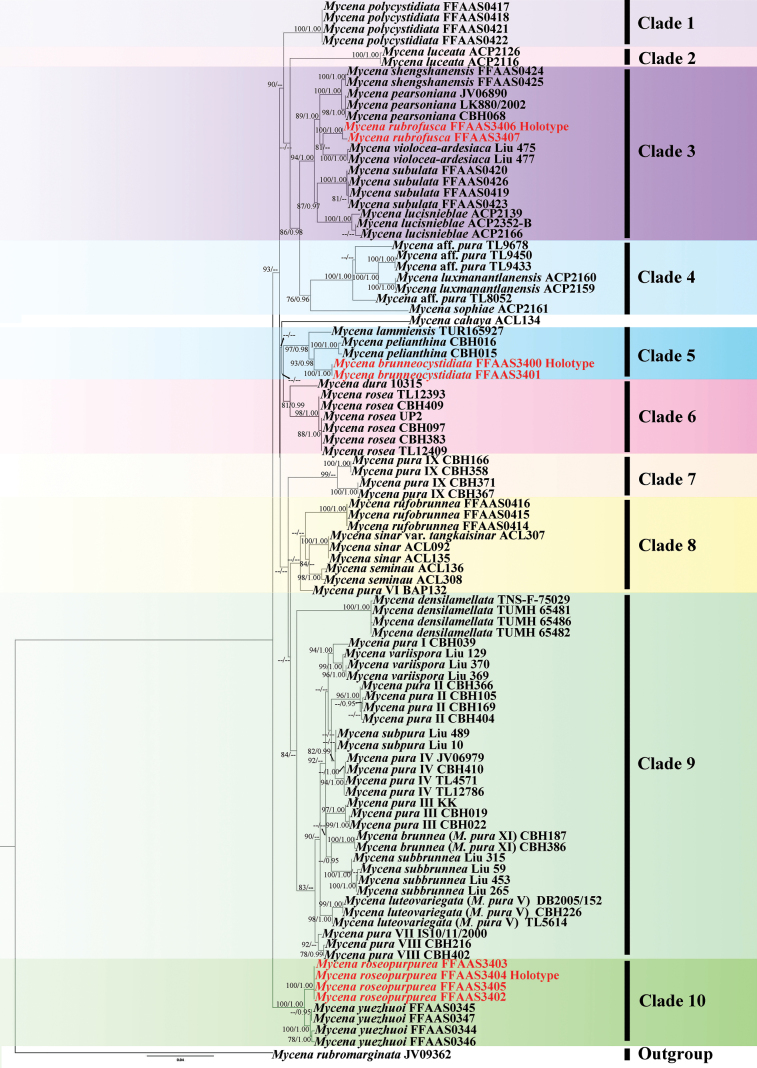 Figure 1
Figure 1 Figure 2
Figure 2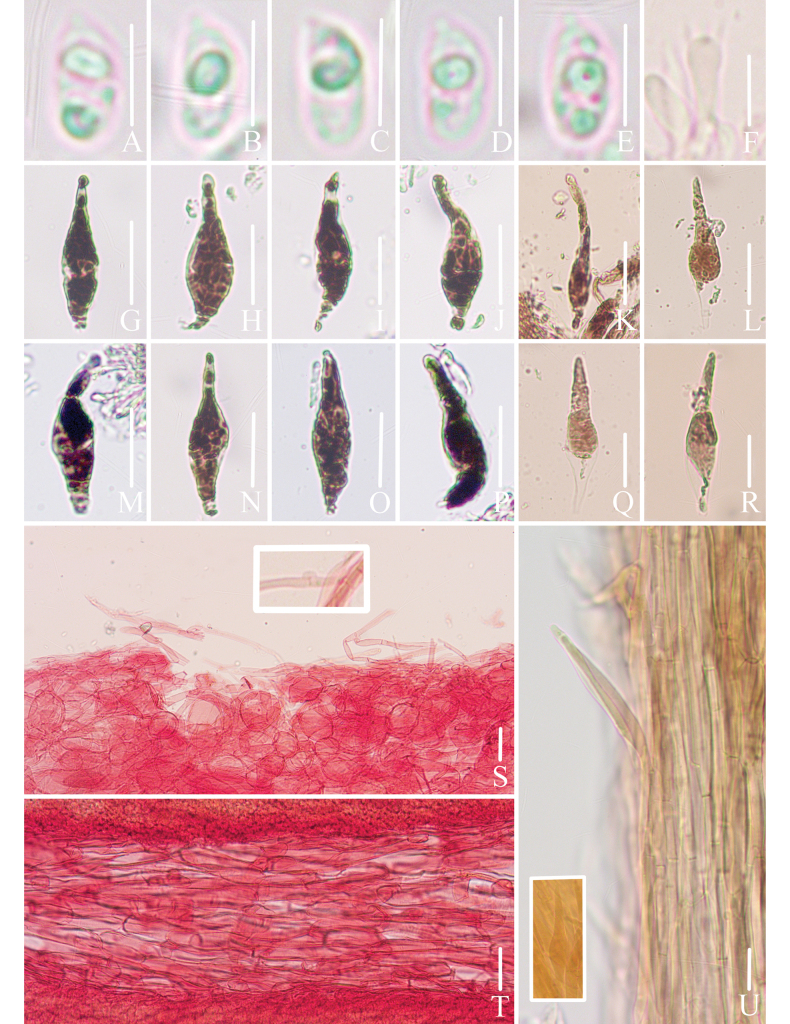 Figure 3
Figure 3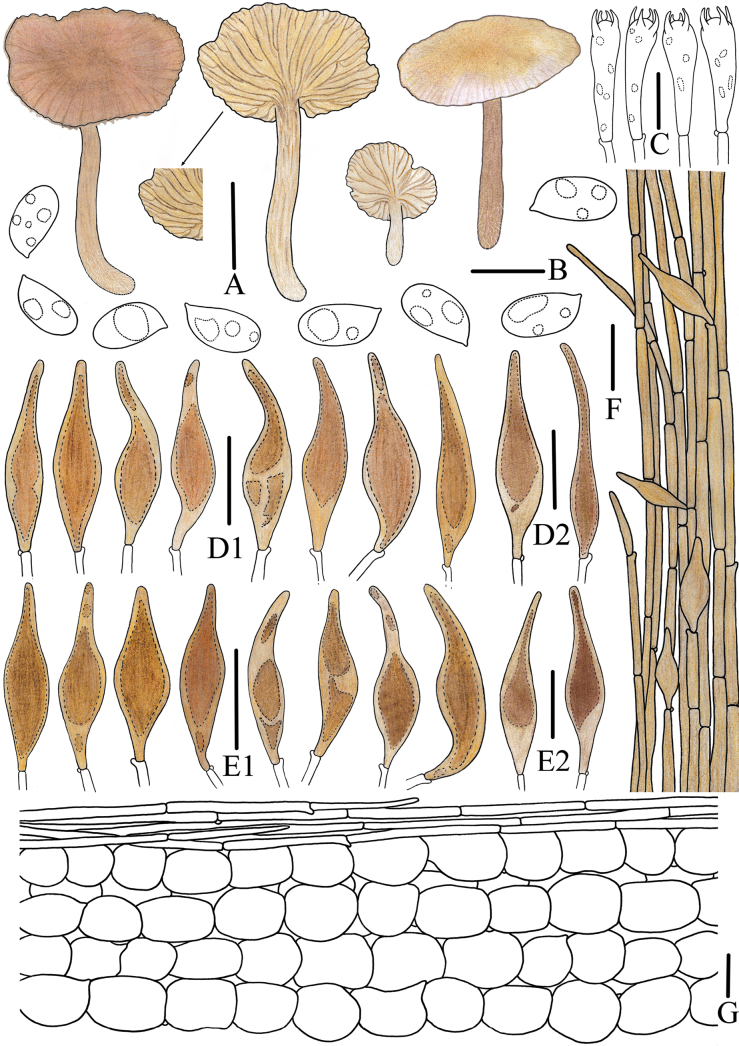 Figure 4
Figure 4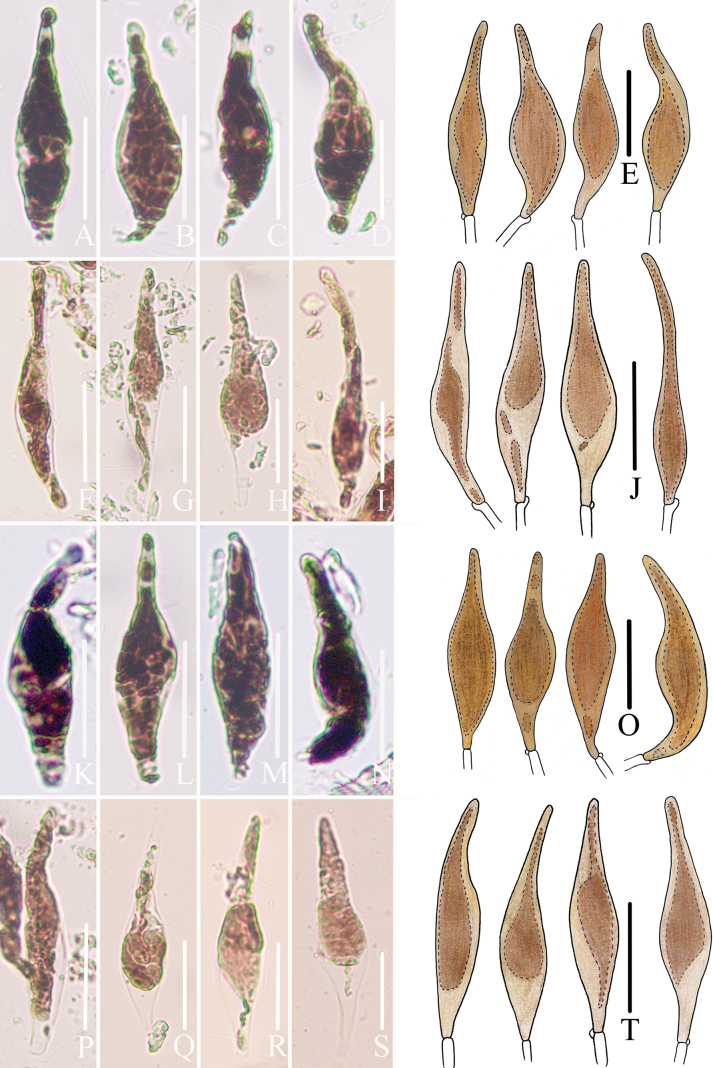 Figure 5
Figure 5 Figure 6
Figure 6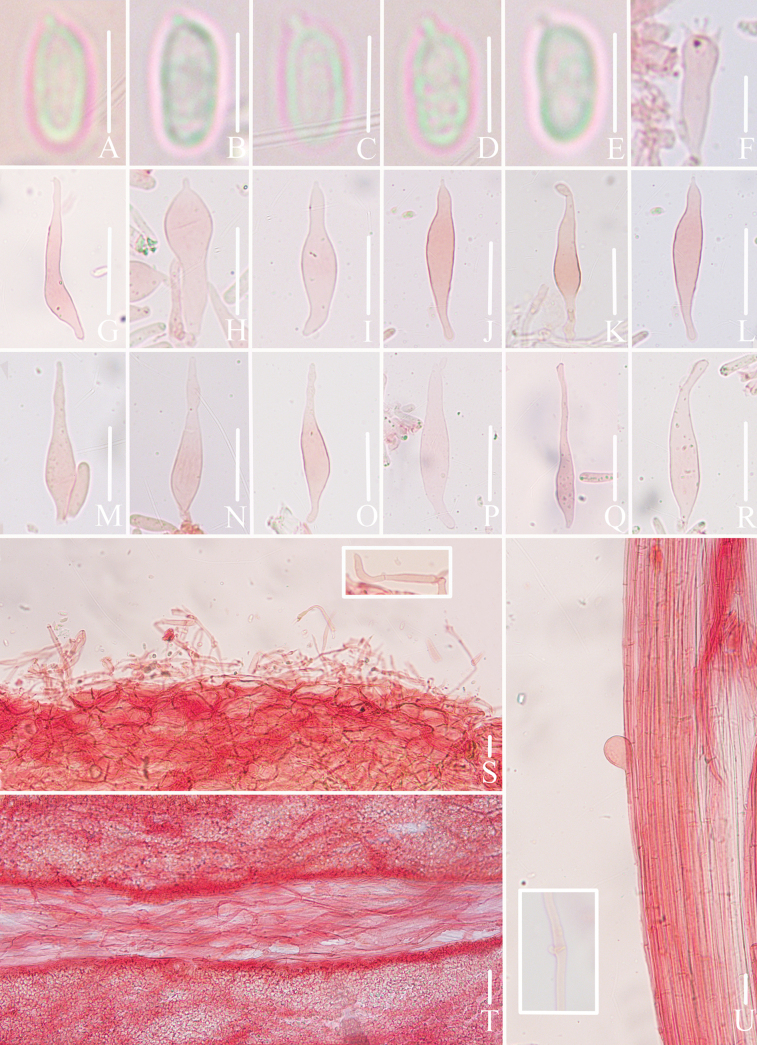 Figure 7
Figure 7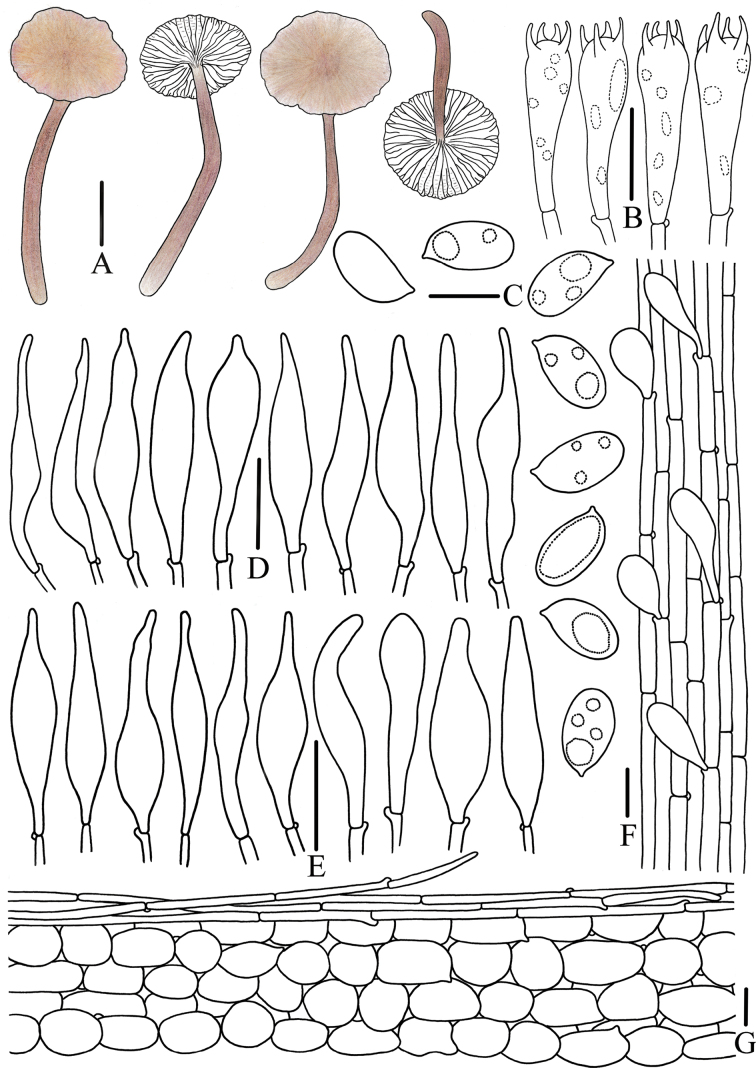 Figure 8
Figure 8 Figure 9
Figure 9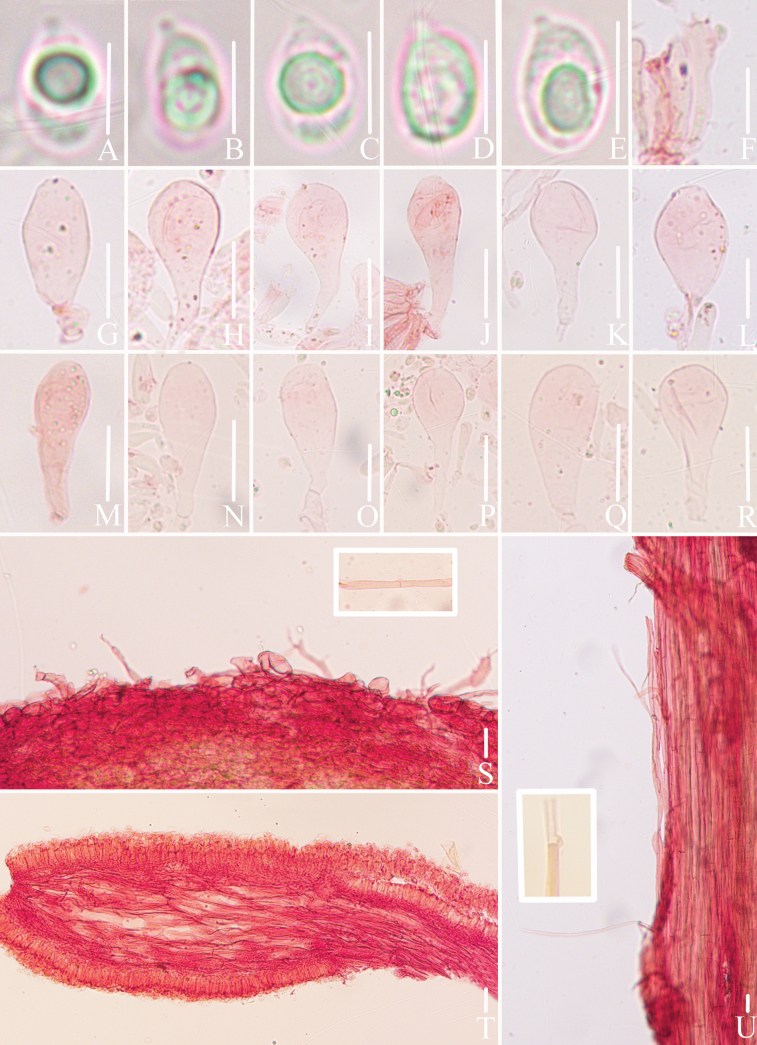 Figure 10
Figure 10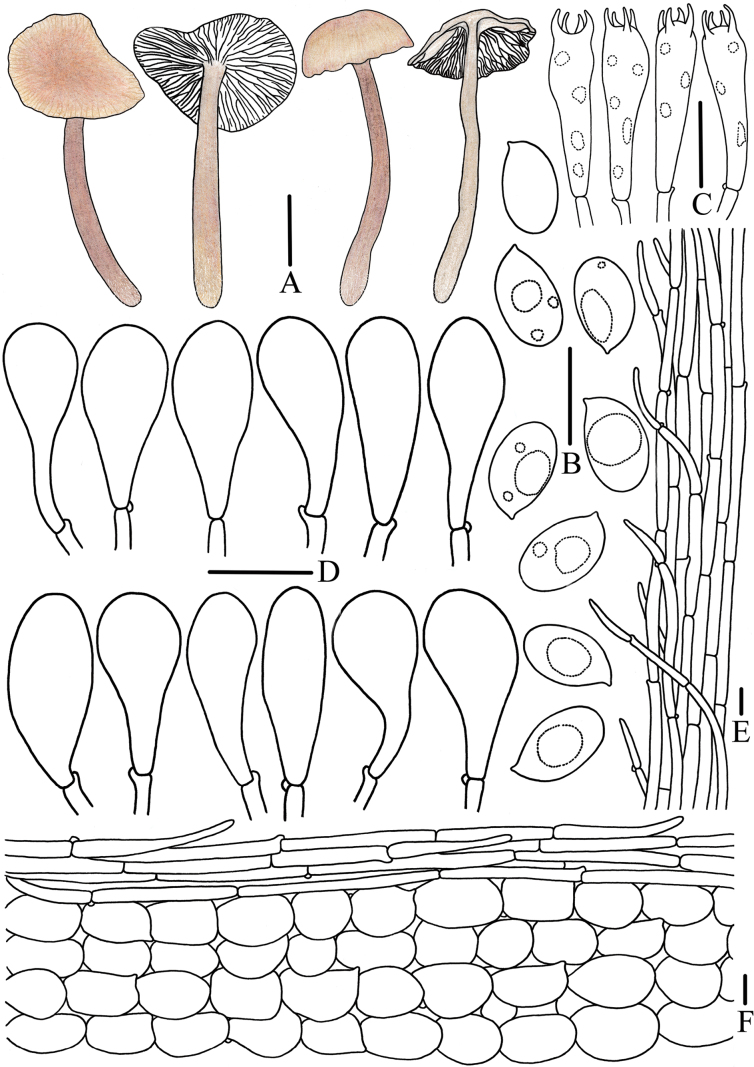 Figure 11
Figure 11| No. | Species | Voucher/Strain No. | GenBank NO. | Locality | Reference | ||
|---|---|---|---|---|---|---|---|
|
|
|
| |||||
| 1 |
| TL8052 |
|
|
| Ecuador | |
| 2 |
| TL9433 |
|
|
| Ecuador | |
| 3 |
| TL9450 |
|
|
| Ecuador | |
| 4 |
| TL9678 |
|
|
| Ecuador | |
| 5 | CBH187 |
|
|
| Denmark | ||
| 6 | CBH386 |
|
|
| Denmark | ||
| 7 |
|
|
|
|
|
|
|
| 8 |
|
|
|
|
|
|
|
| 9 |
| ACL134 |
| – | – | Malaysia |
|
| 10 |
| TUFC 101999 |
|
|
| Japan |
|
| 11 |
| TUMH 65486 |
|
|
| Japan |
|
| 12 |
| TUMH 65482 |
|
|
| Japan |
|
| 13 |
| TNS-F-75029 |
|
|
| Japan |
|
| 14 |
| 10315 |
|
|
| Austria | |
| 15 |
| TUR165927 |
|
|
| Finland | |
| 16 |
| ACP2116 |
|
|
| Mexico |
|
| 17 |
| ACP2126 |
|
|
| Mexico |
|
| 18 |
| ACP2139 |
|
|
| Mexico |
|
| 19 |
| ACP2166 |
|
|
| Mexico |
|
| 20 |
| ACP2352-B |
| – |
| Mexico |
|
| 21 | CBH226 |
|
|
| Denmark | ||
| 22 | DB2005/152 |
| – | – | Denmark |
| |
| 23 | TL5614 |
|
|
| Denmark | ||
| 24 |
| ACP2160 |
|
|
| Mexico |
|
| 25 |
| ACP2159 |
|
|
| Mexico |
|
| 26 |
| LK880/2002 |
|
|
| Germany | |
| 27 |
| CBH068 |
|
|
| Germany | |
| 28 |
| JV06890 |
|
|
| Denmark | |
| 29 |
| CBH015 |
|
|
| Denmark | |
| 30 |
| CBH016 |
|
|
| Denmark | |
| 31 |
| FFAAS0417 |
|
|
| China |
|
| 32 |
| FFAAS0418 |
|
|
| China |
|
| 33 |
| FFAAS0421 |
|
|
| China |
|
| 34 |
| FFAAS0422 |
|
|
| China |
|
| 35 | CBH039 |
|
|
| Denmark | ||
| 36 | CBH105 |
|
|
| Denmark | ||
| 37 | CBH169 |
|
|
| Denmark | ||
| 38 | CBH366 |
|
|
| Denmark | ||
| 39 | CBH404 |
|
|
| Denmark | ||
| 40 | CBH019 |
|
|
| Denmark | ||
| 41 | CBH022 |
|
|
| Denmark | ||
| 42 | KK |
|
|
| Slovakia | ||
| 43 | CBH410 |
|
|
| Denmark | ||
| 44 | JV06979 |
|
|
| Denmark | ||
| 45 | TL4571 |
|
|
| Denmark | ||
| 46 | TL12786 |
|
|
| Sweden | ||
| 47 | BAP132 |
|
|
| USA | ||
| 48 | IS10/11/2000 |
| – | – | USA |
| |
| 49 | CBH216 |
|
|
| Denmark | ||
| 50 | CBH402 |
|
|
| Denmark | ||
| 51 | CBH166 |
|
|
| Denmark | ||
| 52 | CBH358 |
|
|
| Denmark | ||
| 53 | CBH367 |
|
|
| Denmark |
| |
| 54 | CBH371 |
|
|
| Denmark |
| |
| 55 |
| UP2 |
| – | – | UK |
|
| 56 |
| CBH097 |
|
|
| Denmark | |
| 57 |
| CBH383 |
|
|
| Denmark | |
| 58 |
| CBH409 |
|
|
| Germany | |
| 59 |
| TL12393 |
|
|
| Denmark | |
| 60 |
| TL12409 |
|
|
| Denmark | |
| 61 |
|
|
|
|
|
|
|
| 62 |
|
|
|
|
|
|
|
| 63 |
|
|
|
|
|
|
|
| 64 |
|
|
|
|
|
|
|
| 65 |
|
|
|
|
|
|
|
| 66 |
|
|
|
|
|
|
|
| 67 |
| JV09362 |
|
|
| Denmark | |
| 68 |
| FFAAS0414 |
|
|
| China |
|
| 69 |
| FFAAS0415 |
|
|
| China |
|
| 70 |
| FFAAS0416 |
|
|
| China |
|
| 71 |
| ACL308 |
| – | – | Malaysia |
|
| 72 |
| ACL136 |
| – | – | Malaysia |
|
| 73 |
| FFAAS0424 |
|
|
| China |
|
| 74 |
| FFAAS0425 |
|
|
| China |
|
| 75 |
| ACL092 |
| – | – | Malaysia |
|
| 76 |
| ACL135 |
| – | – | Malaysia |
|
| 77 |
| ACL307 |
| – | – | Malaysia |
|
| 78 |
| ACP2161 |
| – |
| Mexico |
|
| 79 |
| Liu 59 |
|
|
| China |
|
| 80 |
| Liu 265 |
|
|
| China |
|
| 81 |
| Liu 315 |
| – |
| China |
|
| 82 |
| Liu 453 |
|
|
| China |
|
| 83 |
| Liu 10 |
| – |
| China |
|
| 84 |
| Liu 489 |
| – |
| China |
|
| 85 |
| FFAAS0419 |
|
|
| China |
|
| 86 |
| FFAAS0420 |
|
|
| China |
|
| 87 |
| FFAAS0423 |
|
|
| China |
|
| 88 |
| FFAAS0426 |
|
|
| China |
|
| 89 |
| Liu 129 |
| – |
| China |
|
| 90 |
| Liu 369 |
| – |
| China |
|
| 91 |
| Liu 370 |
| – |
| China |
|
| 92 |
| Liu 475 |
| – |
| China |
|
| 93 |
| Liu 477 |
| – |
| China |
|
| 94 |
| FFAAS0344 |
|
|
| China |
|
| 95 |
| FFAAS0345 |
|
|
| China |
|
| 96 |
| FFAAS0346 |
|
|
| China |
|
| 97 |
| FFAAS0347 |
|
|
| China |
|
| 1 | Lamellae edge purplish-brown or brown |
|
| – | Lamellae edge white |
|
| 2 | Caulocystidia present |
|
| – | Caulocystidia absent |
|
| 3 | Pileus pink |
|
| – | Pileus not pink |
|
| 4 | Cheilocystidia thick-walled |
|
| – | Cheilocystidia thin-walled |
|
| 5 | Pileus lilac |
|
| – | Pileus not lilac |
|
| 6 | Lamellae surfaces white |
|
| – | Lamellae surfaces lilac |
|
| 7 | Cheilocystidia acicular to lanceolate |
|
| – | Cheilocystidia clavate |
|
| 8 | Stipe light grayish-brown to purplish-brown |
|
| – | Stipe white to pinkish-yellow |
|
| 9 | Lamellae adnexed |
|
| – | Lamellae subfree |
|
| 10 | Basidiospores inamyloid |
|
| – | Basidiospores amyloid |
|
| 11 | Pileus white to cream-colored |
|
| – | Pileus pinkish-purple to purple |
|
| 12 | Cheilocystidia utriform |
|
| – | Cheilocystidia fusiform or clavate |
|
| 13 | Pleurocystidia present |
|
| – | Pleurocystidia absent |
|
| 14 | Caulocystidia present |
|
| – | Caulocystidia absent |
|
| 15 | Pileus white |
|
| – | Pileus not white |
|
| 16 | Lamellae adnate |
|
| – | Lamellae adnexed |
|
| 17 | Pleurocystidia present |
|
| – | Pleurocystidia absent |
|
| 18 | Stipe purple |
|
| – | Stipe not purple |
|
| 19 | Basidiospores (5.6)5.9–7.4(7.8) × 3.0–3.9(4.1) μm, elongated ellipsoid to cylindrical |
|
| – | Basidiospores (8.3)8.6–10.8(11.5) × (5.3)5.4–6.3(6.4) μm, ellipsoid to elongated ellipsoid |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Slime Mold and Myxomycetes Research
Introduction
Mycena (Pers.) Roussel is a typical saprobic fungus which plays a pivotal role in forest ecosystems (Smith 1947; Perry 2002; Kirk et al. 2008; Na 2019). Species in this genus are known for their ability to decompose rotten branches and wood, fallen leaves, and various organic substrates, and could be facilitating nutrient cycling and sustaining energy flow (Fukasawa et al. 2009; Baldrian and Kohout 2017; Kyaschenko et al. 2017; Guerreiro et al. 2023). Within the genus, sect. Calodontes (Fr. Ex Berk.) Quél. is recognized for its large numbers of species and relatively large basidiomata, in which the pileus diameter can reach 6 cm, hygrophanous pileus, irregularly intervenose lamellae, mostly amyloid basidiospores, and smooth cystidia, pileipellis, and stipitipellis (Smith 1947; Maas Geesteranus 1992a, 1992b; Maas Geesteranus and de Meijer 1997; Grgurinovic 2003; Robich 2003; Aronsen and Læssøe 2016; Na 2019; Liu 2023). Mycena sect. Calodontes is primarily distributed in the mid- to high- latitude regions of the Northern Hemisphere, including Europe and America, with only a few species known from lower latitudes in the Southern Hemisphere (Maas Geesteranus 1992a, 1992b; Grgurinovic 2003; Chew et al. 2014; Cooper et al. 2018; Liu et al. 2021, 2022, 2024; Cortés-Pérez et al. 2023). In 1821, Fries erected subtri. Calodontes under Agaricus trib. Clitocybe, later Berkeley (1836) elevated Clitocybe (Fr.) Staude to subgenus rank, and sect. Calodontes was raised to the rank of section, and the concept of sect. Calodontes has been widely adopted (Fries 1821; Berkeley 1836; Quéltet 1875; Maas Geesteranus 1992a, 1992b; Grgurinovic 2003; Aronsen and Læssøe 2016; Na 2019; Liu 2023). By the early 21^st^ century, 44 species of sect. Calodontes are known, including 17 newly species recorded from Asia, 5 from North America, 2 from Africa, and 1 from Oceania (Grgurinovic 2003; Chew et al. 2014; Cooper et al. 2018; Liu et al. 2021, 2022, 2024; Qiang and Bai 2022; Cortés-Pérez et al. 2023; Fan et al. 2024; Nagamune et al. 2024; Xiao et al. 2025). Species of sect. Calodontes adapt to various temperature and humidity conditions, widely growing on the humus layer of coniferous forests in North Temperate and Cold Temperate Zones, a few species also reported from tropical and subtropical climates, including India, Malaysia, São Tomé, and southern Australia (Maas Geesteranus 1992a, 1992b; Grgurinovic 2003; Chew et al. 2014; Cooper et al. 2018; Cortés-Pérez et al. 2023; Liu 2023).
Over the past three decades, the color of pileus, and the shape, contents, thickness of the walls of cheilocystidia have been confirmed to be the diagnostic characters of sect. Calodontes (Maas Geesteranus 1992a, 1992b; Grgurinovic 2003; Robich 2003; Harder et al. 2010, 2013; Liu 2023). The color of pileus in sect. Calodontes, ranging from pink, sulfur yellow, white, purple to brown, is easily changed by the temperature, humidity, and growth stage, so the characteristics of cheilocystidia show more effectiveness (Kauserud et al. 2008; Liu 2023). In sect. Calodontes, cheilocystidia are fusiform, clavate, or utriform (sometimes with tapering apices), and apart from their shape, the thickness of their walls can also be used to distinguish some species from closely related taxa (Maas Geesteranus 1992a, 1992b; Grgurinovic 2003; Robich 2003; Liu et al. 2021, 2022; Liu 2023). According to a phylogenetic reconstruction of sect. Calodontes, based on the materials from Europe and North America, derived from an internal transcribed spacer (ITS), RNA polymerase II largest subunit (RPB1), and translation elongation factor-1 alpha (tef1-α) sequence dataset, the results supported the color of pileus and the characteristics of cheilocystidia can be used to delimit species, but the varieties and forms were not supported (Harder et al. 2010, 2013). Additionally, Harder et al. (2010, 2013) also proposed that the RPB1 and tef1-α sequences improved the ability to identify phylogenetic species in the M. pura complex (Harder et al. 2010, 2013).
Materials and methods
Specimen collection and macroscopic characteristics recording
During field investigations, each specimen was assigned a unique collection number. High-resolution photos were taken using a Canon EOS 90D digital camera (Canon, Tokyo, Japan) equipped with an EF-S 60 mm f/2.8 Macro USM lens. Comprehensive ecological data were recorded, including forest type, substrate, elevation, climate, season, GPS coordinates, and macroscopic characteristics such as pileus, lamellae (intervenose), context, stipe (base), odor, and taste. Color codes and notations followed Ridgway (Ridgway 1912). A small fragment of context was excised from each specimen for molecular analyses after macroscopic documentation. Specimens were dried at 40 °C using a Stöckli dehydrator (A. & J. Stöckli AG, Netstal, Switzerland) and stored in self-sealing plastic bags containing color-changing silica gel. All voucher specimens were deposited in the Fungarium of the Fujian Academy of Agricultural Sciences (FFAAS), China.
Microscopic characteristics’ observation and morphological description
Dried specimens were rehydrated in 5% KOH aqueous solution and examined using a Lab A1 light microscope (Carl Zeiss AG, Jena, Germany). The tissues were stained with 1% Congo red aqueous solution when necessary. Microscopic structures were photographed and measured using ZEN 2.3 software. For each specimen, basidiospores were observed in lateral view, and 20 mature basidiospores were randomly measured. The dimensions of basidiospores and Q values are presented as [a/b/c] (d)e–f–g(h) × (i)j–k–l(m) µm [Q = (n)o–p(q), Q_m_ = r ± s]. The notation [a/b/c] refers to the number of basidiospores, basidiomata, and specimens measured, respectively. Spore dimensions are presented as (d)e–f–g(h) × (i)j–k–l(m), where d and h denote the 5% minimum and maximum values, e–g indicate the central 90% range, and f represents the mean value. Q represents the length-to-width ratio, with Q = (n)o–p(q) indicating the range, and Q_m_ = r ± s denoting the mean and standard deviation. For the type specimen, two basidiomata were examined, with at least 20 mature basidiospores measured per individual, following Na et al. (2021), Liu et al. (2021, 2022), and Wei et al. (2024). Amyloid reactions of basidiospores and lamellar trama were tested using Melzer’s reagent (Vizzini et al. 2020). For all other microscopic structures, at least 20 basidia were measured, as well as measurements of the shape of cheilocystidia, pleurocystidia (if present), and caulocystidia (if present), and the hyphae of the pileipellis and stipitipellis were also measured, the contents, and wall thickness of cheilocystidia, pleurocystidia, and caulocystidia were also observed (Liu et al. 2022). Line drawings were prepared based on habitat photographs, field notes, and microscopic observations, scanned using a Canon LiDE120 scanner (Canon, Tokyo, Japan), and finalized using Adobe Photoshop 2023.
Phylogenetic analyses
DNA sequence acquisition
Genomic DNA was extracted using the New Plant Genomic DNA Extraction Kit (Cowin Century, Beijing, China) and stored at -20 °C. Three nuclear loci, comprising the internal transcribed spacer (ITS), RNA polymerase II largest subunit (RPB1), and translation elongation factor-1 alpha (tef1-α), were amplified using the primer pairs ITS1/ITS4, RPB1Mp_f1/RPB1Mp_r1, tEFMp_f2/tEFMp_r2, respectively (Harder et al. 2010, 2013). PCR reactions were performed in 25 μL volumes, containing 12.5 μL of 2 × Utaq PCR MasterMix (ZomanBio, Beijing, China), 1 μL of each primer, 2 μL of DNA template, with ddH_2_O added to reach the final volume (Liu et al. 2022). The PCR protocol for the ITS region consisted of an initial denaturation at 94 °C for 4 min, followed by 34 cycles of 94 °C for 45 s, 52 °C for 45 s, and 72 °C for 1 min, a final extension of 72 °C for 10 min (Harder et al. 2010). The amplification protocol for the RPB1 and tef1-α regions was as follows: 94 °C for 1 min, then 10 cycles of 94 °C for 35 s, 53 °C for 45 s, 72 °C for 45 s, and 25 cycles of 94 °C for 35 s, 56 °C for 45 s, 72 °C for 45 s, ending with a final extension of 72 °C for 10 min (Harder et al. 2013). PCR products were sequenced by the Beijing Genomics Institute (Beijing, China). The PCR products were cloned using the pBLUE-T Kit (Beijing Zoman Biotechnology Co., Beijing, China) to generate high-quality sequences.
Phylogenetic analyses
All newly generated sequences from the collected specimens were compared using BLAST in the NCBI database (https://www.ncbi.nlm.nih.gov/). Homologous sequences showing nucleotide identity greater than 90% were downloaded from GenBank (https://www.ncbi.nlm.nih.gov/genbank). Mycena rubromarginata (Fr.) P. Kumm was selected as the outgroup for phylogenetic analyses (Harder et al. 2010, 2013; Liu et al. 2022). Sequence alignment was performed using MAFFT v.7.110, with gaps treated as missing data. Peak profiles of the newly generated sequences were examined in BioEdit to detect insertion and deletion sites, and regions containing multiple copies were encoded using degenerate bases (Hall 1999; Katoh et al. 2002, 2019; Alzohairy 2011). Phylogenetic analyses were conducted using both Bayesian Inference (BI) and Maximum Likelihood (ML) methods in MrBayes v3.2.6 and raxmGUI 2.0.10 (Posada and Crandall 1998; Nylander 2004; Edler et al. 2021). The sequence matrix of 3 nuclear loci were divided into 6 partitions: ITS1, 5.8S, ITS2, RPB1 exons, tef1-α exons, and the combined introns regions of RPB1 and tef1-α. MCMC was run with 6 chains, and conducted sampling at intervals of 10,000 generations until the Average Deviation of Split Frequencies was below 0.01; the first 25% of trees were discarded as burn-in, using the ‘sump’ and ‘sumt burnin’ commands to generate the results (Ronquist and Huelsenbeck 2003; Ronquist et al. 2012). Tracer v.1.7.2 was used to evaluate the Effective Sample Size (ESS) and Average Potential Scale Reduction Factor (PSRF) values as indicators of Bayesian inference (BI) analysis (Rambaut et al. 2018; Fabreti and Höhna 2022). For the Maximum Likelihood (ML) analysis, default parameters in RAxML were used with 1,000 rapid bootstrap replicates to assess branch support (Edler et al. 2021). The resulting phylogenetic trees were visualized with FigTree v.1.4.3.
Results
Phylogenetic analyses
The dataset comprised 263 sequences, containing 24 newly generated sequences (8 ITS, 8 RPB1, and 8 tef1-α) and 239 sequences downloaded from GenBank (89 ITS, 70 RPB1, and 80 tef1-α). Detailed information for all sequences was provided in Table 1. The aligned dataset contained 1,538 nucleotide sites (including gaps), with 216 bp for ITS1 region, 159 bp for 5.8S region, 247 bp for ITS2 region, 55 bp for RPB1 exons region, 307 bp for tef1-α exons region, 433 bp for RPB1 introns region, and 121 bp for tef1-α introns region. Among the 24 newly generated sequences, 29 insertion sites and 23 deletion sites were identified, along with 2 degenerate bases, specifically R (1519 bp of FFAAS3406) and W (944 bp of FFAAS3407). For BI analysis, the best-fitting models for each partition of the concatenated dataset were selected as follows: GTR+G for ITS1 and RPB1 introns+tef1-α introns, JC for 5.8S, GTR+I+G for ITS2 and tef1-α exons, and HKY for RPB1 exons. The BI analysis, after 15,000,000 generations, yielded an average deviation of split frequencies of 0.006965, an effective sample size (ESS) of 1160.2, and a potential scale reduction factor (PSRF) ranging from 1.000 to 1.002. For ML analysis, the substitution models were as follows: JC for ITS1, 5.8S, and RPB1 exons, K80+G for ITS2, TVMef+G for tef1-α exons, and HKY+G for RPB1 and tef1-α introns regions. The final log-likelihood score was -8333.401114. The BI and ML analyses tree showed similar topologies, and the ML topology was selected to present the final phylogenetic tree (Fig. 1).
Table 1.: Sequences of Mycena sect. Calodontes used in the phylogenetic analyses.
Bayesian Inference tree based on concatenated ITS+RPB1+tef1-α dataset. Only branch nodes with both Maximum Likelihood bootstrap support values (BS) above 75% and Bayesian posterior probabilities (BPP) exceeding 0.95 are indicated. New taxonomic groups are marked in red.
According to the phylogenetic tree in Fig. 1 10 well-supported clades were identified. The three new species were located in Clade 3, Clade 5, and Clade 10, respectively, each forming an independent lineage with strong statistical support (BS/BPP = 100/1.00). Among the three clades, Clade 3 was morphologically characterized by the absence of pleurocystidia, and 6 species included in the clade: M. rubrofusca, M. shengshanensis Z.W. Liu, Y.P. Ge & Q. Na, M. pearsoniana Dennis ex Singer, M. violocea-ardesiaca Shun Liu & Biao Zhu, M. subulata Z.W. Liu, Y.P. Ge & Q. Na, and M. lucisnieblae Cortés-Pérez, Racm.-Cruz & Guzm.-Dáv. In Clade 3, M. rubrofusca and M. violocea-ardesiaca were identified as the two most closely related taxa but with low support (BS/BPP = 81/--). There were 3 species in Clade 5, M. brunneocystidiata, M. pelianthina (Fr.) Quél., and M. lammiensis Harmaja, the species in the clade have colored lamellae margins and cystidia with colored contents, M. brunneocystidiata showed a close phylogenetic relationship with M. pelianthina (BS/BPP = 93/0.98) than M. lammiensis. Clade 10 merely contained 2 species, M. roseopurpurea and M. yuezhuoi Z.W. Liu, Y.P. Ge & Q. Na, each formed a distinct lineage with statistical support (BS/BPP = 100/1.00). Furthermore, Clades 1, 2, 4, 6, 7, 8, and 9 also each formed separate lineages with high bootstrap and posterior probability values.
Taxonomy
Mycena
brunneocystidiata
Taxon classificationFungiAgaricalesMycenaceae
J.W. Guo, Z.W. Liu, Y.P. Ge & Q. Na sp. nov.
9D5CF2E7-A99B-584C-95DD-68BE40CBA634
860072
Diagnosis.
Pileus brown. Lamellae densely covered with dark brown dots, margin brown. Cheilocystidia, pleurocystidia, caulocystidia and terminal cells of stipitipellis with brownish contents. Differ from M. lammiensis by wider basidiospores (width > 4 μm) and fusiform caulocystidia.
Basidiomata of Mycena brunneocystidiata. A–D. Collection FFAAS3400, holotype; E–I. Collection FFAAS3401. Scale bars: 20 mm (A–B, E, G–H); 1 mm (C, F); 5 mm (D, I). Photographs (A–I) by Jingwen Guo and Yupeng Ge.
Holotype.
China • Heilongjiang Province, Mudanjiang City, Mudanfeng National Forest Park, 42°45'74"N, 128°14'41"E, 22 August 2024, Jingwen Guo, Tian Wang, Qin Na, Zengcai Liu, Ruipeng Liu, Pengyu Du, Ying Yu, and Yupeng Ge leg., FFAAS3400 (collection no. NJ 6538).
Etymology.
Name refers to the cheilocystidia, pleurocystidia, and caulocystidia with brown contents.
Description.
Pileus 10–36 mm in diam., plano-convex, with slightly umbo at center, margin revolute, wavy, cracked at mature; *Drab (XLVI17′′′′) at center, gradually towards margin to PaMid Vinaceous-Drab (XLV5′′′′f), Pale Drab-Gray (XLVI17′′′′f) to *Drab-Gray (XLVI17′′′′d), margin *Drab (XLVI17′′′′); striate *Hair Brown (XLVI17′′′′i), towards the center up to 1/2–2/3 diam.. Context White (LIII), 1.0 mm thick, fragile. Lamellae subdecurrent, 23–27 reaching the stipe, 1–3 tiers of lamellulae, White (LIII), densely covered with Deep Brownish Drab (XLV9′′′′i) dots, irregularly intervenose, stretching downward to 2/3–3/4 of the width of lamellae, edge entirely Deep Brownish Drab (XLV9′′′′i), wavy. Stipe 24–44 × 2–4 mm, central, cylindrical; apex to middle *Smoke Gray (XLVI21′′′′d) to Pale Smoke Gray (XLVI21′′′′f), base *Ecru-Drab (XLVI13′′′′d) to *Drab-Gray (XLVI17′′′′d), hollow, fragile, apex with Light Drab (XLVI17′′′′b) to *Drab (XLVI17′′′′) striates, sparse White (LIII) pubescent at base. Odor and taste not distinctive.
Basidiospores (60/3/2) (5.4)5.6–6.3–6.9(7.1) × (2.6)2.9–3.2–3.5(4.0) μm [Q = (1.72)1.77–2.16(2.19), Q_m_ = 1.97 ± 0.09] [holotype (40/2/1) 6.0–6.5–6.9(7.1) × 3.0–3.2–3.5 μm [Q = (1.83)1.89–2.15(2.18), Q_m_ = 2.00 ± 0.08], narrowly ellipsoid to cylindrical, colorless, smooth (1000×), thin-walled, amyloid. Basidia clavate, 16–21 × 5–7 μm, hyaline, thin-walled, 4-spored, sterigmata 2–3 μm in length. Cheilocystidia fusiform with tapered apices, 36–62 × 9–14 μm (Fig. 3G–J, Fig. 4D1), acicular to lanceolate, 46–84 × 10–17 μm (Fig. 3M–P, Fig. 4D2), with brownish contents, thin-walled, smooth. Pleurocystidia similar to cheilocystidia, 39–72 × 8–15 μm, with brownish contents, thin-walled, smooth. Pileipellis a cutis composed of cylindrical cells, 32–121 × 3–6 μm, smooth, thin-walled; terminal cells cylindrical, apex tapering, 34–87 μm in length, apex 1–3 μm, base 3–7 μm, thin-walled, hyaline. Hypodermium formed by fusiform to subglobose hyphae, 20–72 × 12–39 μm, thin-walled, hyaline. Lamellar trama subregular, dextrinoid. Stipitipellis a cutis composed of cylindrical hyphae, 9–18 μm in diam., smooth, thin-walled; terminal cells fusiform or cylindrical, apex tapering, 31–72 × 4–8 μm, with pale brownish contents, thin-walled, smooth; caulocystidia fusiform, 33–70 × 7–13 μm, with pale brownish contents, thin-walled, smooth. Clamps present in all tissues.
Microscopic features of Mycena brunneocystidiata (FFAAS3400, holotype). A–E. Basidiospores; F. Basidia; G–J. Fusiform cheilocystidia; K–L. Acicular to lanceolate cheilocystidia; M–P. Fusiform pleurocystidia; Q–R. Acicular to lanceolate pleurocystidia; S. Pileipellis and hypodermium; T. Lamellar trama; U. Stipitipellis and caulocystidia. Scale bars: 5 μm (A–E); 10 μm (F); 25 μm (G–R); 20 μm (S–U). Structures (A–E) were rehydrated in 5% KOH aqueous solution, (G–R, U) were rehydrated in sterile water and (F, S–T) were stained in 1% Congo red aqueous solution.
Morphological features of Mycena brunneocystidiata (FFAAS3400, holotype). A. Basidiomata; B. Basidiospores; C. Basidia; D1. Fusiform cheilocystidia; D2. Acicular to lanceolate cheilocystidia; E1. Fusiform pleurocystidia; E2. Acicular to lanceolate pleurocystidia; F. Stipitipellis and caulocystidia; G. Pileipellis and hypodermium. Scale bars: 20 mm (A); 5 μm (B); 10 μm (C); 25 μm (D1–G). Drawings by Jingwen Guo.
Habit and habitat.
Scattered on the litter layer in Acer mono Maxim., Larix gmelinii (Ruprecht) Kuzeneva, Pinus koraiensis Siebold et Zuccarini, and Quercus mongolica Fischer ex Ledebour mixed forests during summer and autumn.
Known distribution.
Heilongjiang Province, Jilin Province, China.
Additional material examined.
China • Jilin Province, Yanbian Korean Autonomous Prefecture, Antu County, Erdaobaihe Town, Back Mountain of Changbai Mountain Natural History Museum, 42°46'43"N, 128°14'49"E, 17 August 2021, Zewei Liu, Qin Na, Shixin Wang, and Yupeng Ge leg., FFAAS3401 (collection no. MY 0611).
Notes.
Mycena brunneocystidiata is considered to be a distinct species of sect. Calodontes subsect. Marginatae J. E. Lange on account of its lamellae margins brown and cheilocystidia, pleurocystidia, and caulocystidia with brown contents (Maas Geesteranus 1992a, 1992b). In the subsection, M. lammiensis and M. pelianthina share the same colored lamellae margins, cheilocystidia, and pleurocystidia with purple-brown contents, but M. lammiensis differs in having larger basidiospores (7.5–9.0 × 4.0–5.0 μm) and cylindrical caulocystidia, while M. pelianthina is identified by purple-brown pileus and lacking caulocystidia (Harmaja 1985; Robich 2003; Harder et al. 2010; Aronsen and Læssøe 2016). Mycena shengshanensis resembles M. brunneocystidiata in growing on the humus layer of Larix gmelinii and having light purple-brown to brown pileus, but differs in colorless, clavate, and thick-walled cheilocystidia (Liu et al. 2022).
The main shape of cheilocystidia and pleurocystidia in FFAAS3401 is fusiform with tapered apices (Fig. 5A–E, K–O), but in specimen FFAAS3400, acicular to lanceolate cheilocystidia and pleurocystidia can be observed occasionally, which are larger than the fusiform ones (Fig. 5F–J, P–T).
Morphological features of the Cheilocystidia and pleurocystidia of Mycena brunneocystidiata. A–E. Cheilocystidia of FFAAS3401; F–J. Cheilocystidia of FFAAS3400, holotype; K–O. Pleurocystidia of FFAAS3401; P–T. Pleurocystidia of FFAAS3400, holotype. Scale bars: 25 μm (A–E, K–O); 30 μm (F–J, P–T). Drawings by Jingwen Guo.
Mycena
roseopurpurea
Taxon classificationFungiAgaricalesMycenaceae
J.W. Guo, Z.W. Liu, Y.P. Ge & Q. Na sp. nov.
07E18D40-9E3A-5152-B03F-A94798B95305
860104
Diagnosis.
Pileus light pinkish-purple, light brown at center, hygrophanous when old. Cheilocystidia and pleurocystidia fusiform with tapered apices, thin-walled. Differ from M. subulata by lacking pleurocystidia and having acicular to lanceolate, thick-walled cheilocystidia.
Basidiomata of Mycena roseopurpurea. A–C. Collection FFAAS3405; D–G. Collection FFAAS3404, holotype; H–J. Collection FFAAS3403; K–L. Collection FFAAS3402. Scale bars: 10 mm (A–E, G, I–J); 5 mm (F, H, K–L). Photographs (A–L) by Qin Na.
Holotype.
China • Heilongjiang Province, Yichun City, Liangshui National Nature Reserve, 47°12'74"N, 128°52'86"E, 21 August 2021, Zewei Liu, Qin Na, Shixin Wang, and Yupeng Ge leg., FFAAS3404 (collection no. MY 0660).
Microscopic features of Mycena roseopurpurea (FFAAS3404, holotype). A–E. Basidiospores; F. Basidia; G–L. Cheilocystidia; M–R. Pleurocystidia; S. Pileipellis and hypodermium; T. Lamellar trama; U. Stipitipellis and caulocystidia. Scale bars: 5 μm (A–E); 15 μm (F); 30 μm (G–U). Structures (A–E) were rehydrated in 5% KOH aqueous solution and (F–U) were stained in 1% Congo red aqueous solution.
Etymology.
Name refers to light pinkish-purple pileus.
Morphological features of Mycena roseopurpurea (FFAAS3404, holotype). A. Basidiomata; B. Basidia; C. Basidiospores; D. Cheilocystidia; E. Pleurocystidia; F. Stipitipellis and caulocystidia; G. Pileipellis and hypodermium. Scale bars: 10 mm (A); 15 μm (B); 5 μm (C); 30 μm (D–G). Drawings by Jingwen Guo.
Description.
Pileus 13–22 mm in diam., oblate hemispherical to plano-convex, with slightly depressed at center, margin wavy, occasionally cracked at mature; *Ecru-Drab (XLVI13′′′′d), Light Drab (XLVI17′′′′b) to *Drab-Gray (XLVI17′′′′d) at center, gradually towards margin to Pale Verbena Violet (XXXVI55′′f), Light Pinkish Lilac (XXXVII65′′f), Pale Grayish Vinaceous (XXXIX9′′′f), margin White (LIII); striate Pale Smoke Gray (XLVI21′′′′f), Pale Ecru-Drab (XLVI13′′′′f) to *Ecru-Drab (XLVI13′′′′d), towards the center up to 1/3 diam.; surface dry, hygrophanous when old. Context White (LIII), 1.0 mm thick, fragile. Lamellae sinuate to subdecurrent, 21–28 reaching the stipe, 1–3 tiers of lamellulae, White (LIII), irregularly intervenose, stretching downward to 1/3–3/4 of the width of lamellae, edge concolorous, wavy, serrated. Stipe 32–57 × 1–4 mm, central, cylindrical; apex Pale Drab-Gray (XLVI17′′′′f), Pale Vinaceous-Drab (XLV5′′′′d), Pale Brownish Drab (XLV9′′′′d), lower part Pale Purple-Drab (XLV1′′′′d), Light Purple-Drab (XLV1′′′′b) to *Ecru-Drab (XLVI13′′′′d), hollow, fragile; sparse White (LIII) pubescent at base. Odor and taste raphanoid.
Basidiospores (100/5/4) (5.6)5.9–6.7–7.4(7.8) × 3.0–3.5–3.9(4.1) μm [Q = (1.70)1.72–2.10, Q_m_ = 1.90 ± 0.09] [holotype (40/2/1) (6.0)6.2–6.7–7.4(7.7) × 3.0–3.5–3.9(4.1) μm, Q = (1.71)1.77–2.12, Q_m_ = 1.93 ± 0.11], narrowly ellipsoid to cylindrical, colourless, smooth (1000×), thin-walled, amyloid. Basidia clavate, 18–31 × 4–7 μm, hyaline, thin-walled, 4-spored, sterigmata 2–4 μm in length. Cheilocystidia fusiform, subfusiform, with apices tapered, 35–97 × 7–18 μm, thin-walled, smooth. Pleurocystidia similar to cheilocystidia, 30–80 × 6–18 μm, thin-walled, smooth. Pileipellis a cutis composed of cylindrical cells, 29–81 × 3–11 μm, smooth, thin-walled; terminal cells cylindrical, apex tapering, 20–99 μm in length, apex 2–4 μm, base 2–9 μm, thin-walled, hyaline. Hypodermium formed by fusiform to subglobose hyphae, 20–76 × 6–37 μm, thin-walled, hyaline. Lamellar trama subregular, dextrinoid. Stipitipellis a cutis composed of cylindrical hyphae, 4–17 μm in diam., smooth, thin-walled; caulocystidia fusiform, clavate, sometimes utriform, 23–66 × 6–20 μm, thin-walled, smooth. Clamps present in all tissues.
Habit and habitat.
Scattered on the litter layers in Betula platyphylla Suk., Larix gmelinii, Pinus koraiensis, and P. syluestriformis (Takenouchi) T.Wang ex Cheng mixed forests during summer and autumn.
Known distribution.
Heilongjiang Province, Jilin Province, China.
Additional material examined.
China • Heilongjiang Province, Yichun City, Liangshui National Nature Reserve, 47°12'74"N, 128°52'56"E, 20 August 2021, Zewei Liu, Qin Na, Shixin Wang, and Yupeng Ge leg., FFAAS3403 (collection no. MY 0635); • same location, 21 August 2021, Zewei Liu, Qin Na, Shixin Wang, and Yupeng Ge leg., FFAAS3405 (collection no. MY 0668). • Jilin Province, Yanbian Korean Autonomous Prefecture, Antu County, Erdaobaihe Town Beauty Pine Forest, 42°45'74"N, 128°14'41"E, 18 August 2021, Zewei Liu, Qin Na, Shixin Wang, and Yupeng Ge leg., FFAAS3402 (collection no. MY 0625).
Notes.
Mycena subulata, originally described from Heilongjiang province in China, can be easily mistaken for M. roseopurpurea due to morphological similarity, and growing on the humus layer of mixed forests of Larix gmelinii and Pinus koraiensis, but M. subulata is distinguished by acicular to lanceolate and thick-walled cheilocystidia (Liu et al. 2022). Mycena pearsoniana, while having similar stipe color and lacking pleurocystidia, differs by subfusiform or clavate cheilocystidia and inamyloid basidiospores (Smith 1947; Dennis 1970; Kost 2002; Harder et al. 2012; Aronsen and Læssøe 2016; Na 2019; Kwon et al. 2020). Mycena dura Maas Geesteranus & Hauskn. and M. subcorticalis (Cooke & Massee) Sacc., reported from Europe and southern Australia, resemble M. roseopurpurea; however, M. dura is distinctively characterized by its growth in grasslands and white stipe, and M. subcorticalis is distinguished by inamyloid basidiospores and gelatinized pileipellis (Saccardo 1891; Maas Geesteranus and Hausknecht 1994; Grgurinovic 2003; Olariaga et al. 2015). Mycena pura differs by having light purple to purplish-red pileus and pinkish-purple to purple stipe (Maas Geesteranus 1992a, 1992b; Perry 2002; Robich 2003; Thormann et al. 2006; Aronsen and Læssøe 2016; Na 2019; Okon et al. 2022; Liu 2023).
Mycena
rubrofusca
Taxon classificationFungiAgaricalesMycenaceae
J.W. Guo, Z.W. Liu, Y.P. Ge & Q. Na, sp. nov.,
FBB06453-D7BB-5059-B93C-E1AA68BD7753
860105
Diagnosis.
Pileus light reddish-brown to light grayish-brown, near margin hygrophanous. Pleurocystidia and caulocystidia absent. Differ from M. polycystidiata Z.W. Liu, Y.P. Ge, L. Zou & Q. Na by having pleurocystidia and caulocystidia.
Basidiomata of Mycena rubrofusca. A–E. Collection FFAAS3406, holotype; F–J. Collection FFAAS3407. Scale bars: 15 mm (A–B, F–G); 1 mm (C–D); 5 mm (E, H–J). Photographs (A–J) by Jingwen Guo and Yupeng Ge.
Holotype.
China • Heilongjiang Province, Mudanjiang City, Mudanfeng National Forest Park, 42°45'74"N, 128°14'41"E, 22 August 2024, Jingwen Guo, Tian Wang, Qin Na, Zengcai Liu, Ruipeng Liu, Pengyu Du, Ying Yu, and Yupeng Ge leg., FFAAS3406 (collection no. NJ 6508).
Microscopic features of Mycena rubrofusca (FFAAS3406, holotype). A–E. Basidiospores; F. Basidia; G–R. Cheilocystidia; S. Pileipellis and hypodermium; T. Lamellar trama; U. Stipitipellis. Bars: 5 μm (A–E); 15 μm (F, T, U); 25 μm (G–R); 30 μm (S). Structures (A–E) were rehydrated in 5% KOH aqueous solution and (F–U) were stained in 1% Congo red aqueous solution.
Etymology.
Name refers to the light reddish-brown, light grayish brown to brown pileus.
Morphological features of Mycena rubrofusca (FFAAS3406, holotype). A. Basidiomata; B. Basidiospores; C. Basidia; D. Cheilocystidia; E. Stipitipellis; F. Pileipellis and hypodermium. Scale bars: 15 mm (A, C, E–F); 5 μm (B); 25 μm (D). Drawings by Jingwen Guo.
Description.
Pileus 17–26 mm in diam., hemispherical, plano-convex at mature, margin wavy; Light Drab (XLVI17′′′′b) at center, gradually towards margin to Light Vinaceous-Fawn (XL13′′′d), Pale Ecru-Drab (XLVI13′′′′f) to *Drab-Gray (XLVI17′′′′d), margin Light Drab (XLVI17′′′′b) to *Drab (XLVI17′′′′); striate *Drab (XLVI17′′′′), towards the center up to 1/8–1/7 diam.; surface moist, near margin hygrophanous. Context White (LIII), 1.0 mm thick, fragile. Lamellae adnexed or subdecurrent, approximately 38 reaching the stipe, 1–3 tiers of lamellulae, White (LIII), irregularly intervenose, stretching downward to 2/3–3/4 of the width of lamellae, edge concolorous, wavy. Stipe 44–45 × 2–4 mm, central, cylindrical; apex to lower part Vinaceous-Drab (XLV5′′′′), Light Brownish Drab (XLV9′′′′b) to *Hair brown (XLVI17′′′′i), base Light Grayish Olive (XLVI21′′′′b), hollow, fragile; base swollen, sparse White (LIII) pubescent at base. Odor and taste faintly raphanoid.
Basidiospores (40/2/2) (5.1)6.2–7.4–8.5(8.8) × (3.8)4.0–4.6–5.5(5.7) μm [Q = (1.52)1.55–1.84(1.89), Q_m_ = 1.69 ± 0.10] [holotype (20/1/1) (6.8)7.2–7.6–8.3(8.5) × (3.8)4.1–4.6–4.8(5.0) μm [Q = 1.60–1.81, Q_m_ = 1.70 ± 0.08], ellipsoid to narrowly ellipsoid, colourless, smooth (1000×), thin-walled, amyloid. Basidia clavate, 16–28 × 5–8 μm, hyaline, thin-walled, 4-spored, sterigmata 2–4 μm in length. Cheilocystidia utriform, clavate, 24–67 × 8–23 μm, thin-walled, smooth. Pleurocystidia absent. Pileipellis a cutis composed of cylindrical cells, 28–76 × 5–15 μm, smooth, thin-walled; terminal cells cylindrical, apex tapering, 33–75 μm in length, apex 2–9 μm, base 3–11 μm, thin-walled, hyaline. Hypodermium formed by fusiform to subglobose hyphae, 26–84 × 11–34 μm, thin-walled, hyaline. Lamellar trama subregular, dextrinoid. Stipitipellis a cutis composed of cylindrical hyphae, 6–16 μm in diam, smooth, thin-walled; projecting terminal cells cylindrical or fusiform, 31–73 × 4–9 μm, thin-walled, smooth, hyaline; caulocystidia absent. Clamps present in all tissues.
Habit and habitat.
Scattered on the litter layers in Fraxinus mandschurica Rupr., Pinus koraiensis, and Tilia amurensis Rupr. mixed forests during summer and autumn.
Known distribution.
Heilongjiang Province, China.
Additional material examined.
China • Heilongjiang Province, Hegang City, Taipinggou National Nature Reserve, 48°12'43"N, 130°66'04"E, 3 September 2021, Zewei Liu, Qin Na, Shixin Wang, and Yupeng Ge leg., FFAAS3407 (collection no. MY 0793).
Notes.
Mycena polycystidiata is the closest species to M. rubrofusca because it has a light grayish-brown to brown pileus and stipe, and utriform cheilocystidia, but M. polycystidiata has pleurocystidia and caulocystidia (Liu et al. 2022). Mycena pura, a widely distributed species in the North temperate zone, differs from M. rubrofusca by its fusiform, clavate, or subglobose cheilocystidia and pleurocystidia, and clavate or conical caulocystidia (Maas Geesteranus 1992a, 1992b; Perry 2002; Robich 2003; Thormann et al. 2006; Aronsen and Læssøe 2016; Na 2019; Okon et al. 2022; Liu 2023). Due to the light gray-brown to light brown pileus, M. variispora Shun Liu & Biao Zhu and M. subbrunnea Shun Liu & Biao Zhu are difficult to distinguish from M. rubrofusca, but they can be distinguished by the occurrence in mixed forests of Larix gmelinii and Betula platyphylla, purple lamellae, and clavate, thick-walled cheilocystidia (Liu et al. 2024). Mycena pearsoniana differs from M. rubrofusca in having purple pileus and clavate or fusiform caulocystidia (Smith 1947; Dennis 1970; Kost 2002; Harder et al. 2012; Aronsen and Læssøe 2016; Na 2019; Kwon et al. 2020). Mycena rufobrunnea Z.W. Liu, Y.P. Ge & Q. Na and M. vinacea Cleland, which were reported from northeastern China and southern Australia, respectively, both resemble M. rubrofusca in having utriform cheilocystidia (Cleland 1931; Liu et al. 2022). However, both of them also have clavate cheilocystidia; M. rufobrunnea is distinguished by a light brown to reddish-brown pileus, whereas M. vinacea is characterized by cylindrical pleurocystidia and basidiospores (Cleland 1931; Liu et al. 2022).
Key to 20 species belonging to Mycena
sect. Calodontes in China
**: **
Discussion
The comprehensive morphological characteristics of sect. Calodontes cannot be currently shown by the taxonomic systems (Harder et al. 2010, 2013; Liu 2023). Three subsections were proposed based on the amyloid reaction of basidiospores, the presence of colored contents in cheilocystidia and pleurocystidia, and the presence of pleurocystidia, namely subsect. Purae Konrad & Maubl., subsect. Marginatae, and subsect. Violacellae Singer ex Maas Geest. (Maas Geesteranus 1992a, 1992b). In this study, M. brunneocystidiata was assigned to subsect. Marginatae due to its amyloid basidiospores and cheilocystidia, pleurocystidia, and caulocystidia with brown contents; M. roseopurpurea was assigned to subsect. Purae owing to its amyloid basidiospores and having colorless cheilocystidia and pleurocystidia. However, M. rubrofusca, which was assigned to subsect. Violacellae due to its lack of pleurocystidia, cannot be placed in this subsection on account of its amyloid basidiospores. Notably, Harder et al. (2012) and Pérez-De-Gregorio (2024) reported that M. pearsoniana was assigned to subsect. Violacellae, which was defined by Maas Geesteranus based on inamyloid basidiospores, but its basidiospores showed inamyloid to weakly amyloid basidiospores after 40 minutes (Maas Geesteranus 1992a, 1992b; Harder et al. 2012; Pérez-De-Gregorio 2024). The subsections and species were also not supported by the multi-locus (ITS+RPB1+tef1-α) analyses, subsect. Marginatae was monophyletic, and subsect. Purae and subsect. Violacellae were polyphyletic (Harder et al. 2010, 2012, 2013; Chew et al. 2014; Liu 2023). Species clustered into a single clade with high support in phylogenetic analyses showed significant morphological differences. Mycena subbrunnea, M. variispora, M. subpura Shun Liu & Biao Zhu, and 7 phylogenetic species from the M. pura complex were gathered in the same clade, but the shape and wall thickness of cheilocystidia, and the presence or absence of pleurocystidia were different (Maas Geesteranus 1992a, 1992b; Perry 2002; Robich 2003; Thormann et al. 2006; Harder et al. 2010, 2013; Liu 2023; Liu et al. 2024).
According to previous literature, species of sect. Calodontes predominantly inhabit coniferous forests or coniferous-broadleaved mixed forests in Europe, North America, Asia, and Northern Africa, mainly within the Northern Hemisphere (Maas Geesteranus 1992a, 1992b; Robich 2003; Harder et al. 2010, 2013; Chew et al. 2014; Aronsen and Læssøe 2016; Cooper et al. 2018; Cortés-Pérez et al. 2023). Only a few occur in grasslands or in broadleaved forests of the Southern Hemisphere, southern Oceania (Grgurinovic 2003). In China, species of sect. Calodontes are predominantly found in the litter layer of coniferous-broadleaved mixed forests in Northeast China, and in coniferous forests in East, Southwest, Northwest, and Southeast China (Bau et al. 2019; Na 2019; Liu et al. 2021, 2022, 2024; Liu 2023; Fan et al. 2024; Xiao et al. 2025). Most species of sect. Calodontes, including the three new species in this study, are distributed in low-altitude regions (< 1000 m), with a few recorded in mid-altitude areas (1000–3500 m), and no species have been reported from high-altitude regions (3500–5000 m) (Maas Geesteranus 1992a, 1992b; Maas Geesteranus and de Meijer 1997; Grgurinovic 2003; Cooper et al. 2018; Bau et al. 2019; Na 2019; Liu 2023; Gao et al. 2024).
Supplementary Material
XML Treatment for Mycena brunneocystidiata
XML Treatment for Mycena roseopurpurea
XML Treatment for Mycena rubrofusca
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alzohairy AM (2011) Bio Edit: An Important Software for Molecular Biology.GERF Bulletin of Biosciences 2(1): 60–61.
- 2Aronsen ALæssøe T (2016) The Genus Mycena s.l. Fungi of Northern Europe Vol. 5. Gylling: Narayana Press, 1–373.
- 3Baldrian P Kohout P (2017) Interactions of saprotrophic fungi with tree roots: Can we observe the emergence of novel ectomycorrhizal fungi? The New Phytologist 215(2): 511–513. 10.1111/nph.1466528631323 · doi ↗ · pubmed ↗
- 4Bau T Wang XS Zhang P (2019) Floristic of agarics and boletus in the Greater and Lesser Khinggan Mountains.Biodiversity Science 27(8): 867–873. 10.17520/biods.2019048 · doi ↗
- 5Berkeley MJ (1836) The English flora (Vol. 5, Part 2). Longman, Hurst, Rees, Orme, Brown, and Green, London, 55–64. 10.5962/bhl.title.6340 · doi ↗
- 6Chew ALC Tan YS Desjardin DE Musa MY Sabaratnam V (2014) Four new bioluminescent taxa of Mycena Sect. Calodontes from Peninsula’r Malaysia.Mycologia 106(5): 976–988. 10.3852/13-27424891424 · doi ↗ · pubmed ↗
- 7Cleland JB (1931) Australian Fungi. notes and descriptions. No.8.Transactions and Proceedings of the Royal Society of South Australia (Incorporated)55: 152–160.
- 8Cooper A Desjardin DE Perry BA (2018) The genus Mycena (Basidiomycota, Agaricales, Mycenaceae) and allied genera from Republic of São Tomé and Príncipe, West Africa.Phytotaxa 383(1): 1–47. 10.11646/phytotaxa.383.1.1 · doi ↗