Two new species of genus Leucoagaricus and Leucocoprinus (Agaricaceae, Agaricales) from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
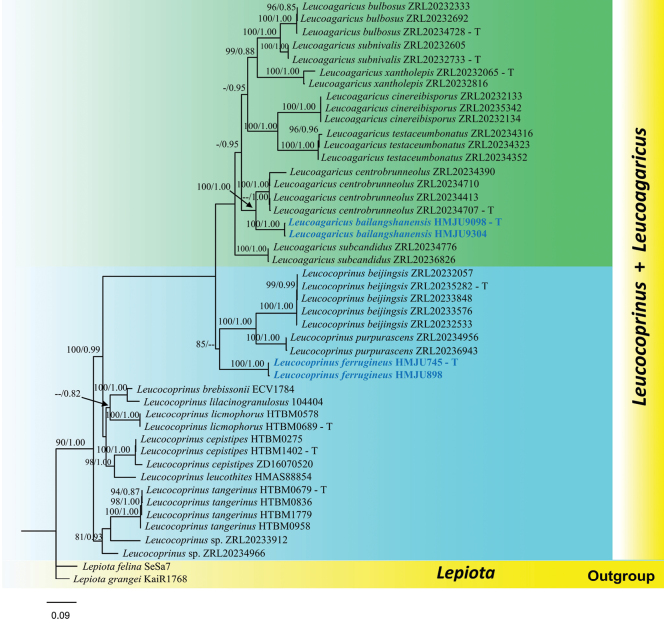 Figure 1
Figure 1 Figure 2
Figure 2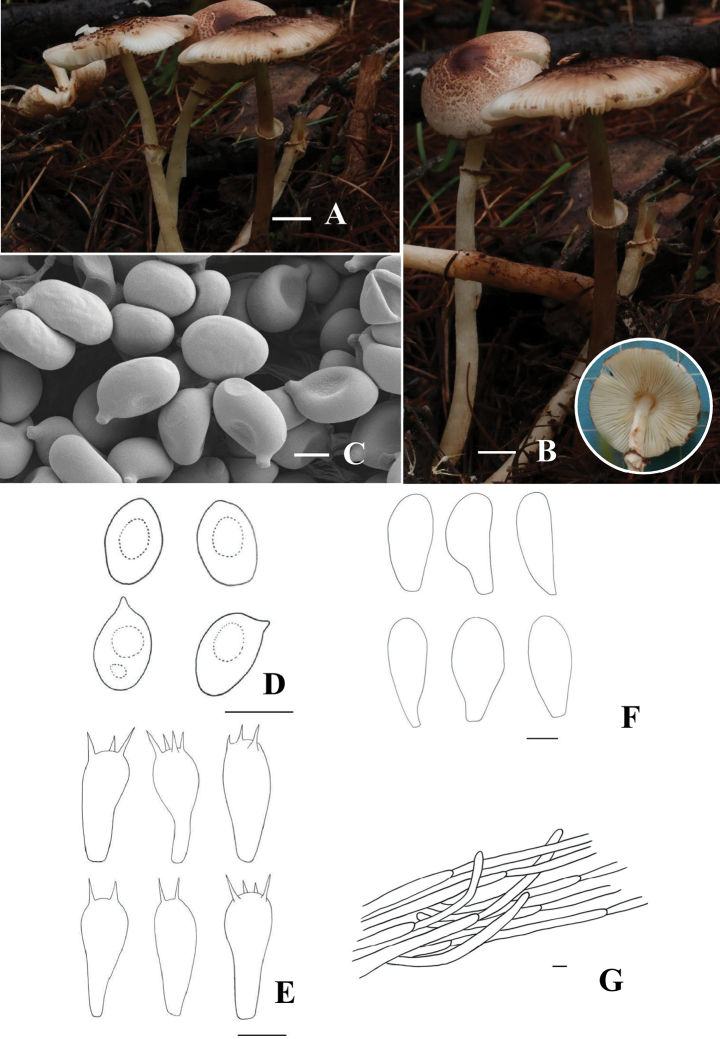 Figure 3
Figure 3| Species | Voucher | GenBank accession number | References | |||
|---|---|---|---|---|---|---|
| ITS | nrLSU |
|
| |||
|
| ZRL20232333 |
|
|
|
| ( |
|
| ZRL20232692 |
|
|
|
| ( |
|
| ZRL20234728 |
|
|
|
| ( |
|
| ZRL20234390 |
|
|
|
| ( |
|
| ZRL20234710 |
|
|
|
| ( |
|
| ZRL20234413 |
|
|
|
| ( |
|
| ZRL20234707 |
|
|
|
| ( |
|
| ZRL20232133 |
|
|
|
| ( |
|
| ZRL20232134 |
|
|
|
| ( |
|
| ZRL20235342 |
|
|
|
| ( |
|
| ZRL20234776 |
|
|
|
| ( |
|
| ZRL20236826 |
|
|
|
| ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| ZRL20232605 |
|
|
|
| ( |
|
| ZRL20232733 |
|
|
|
| ( |
|
| ZRL20234316 |
|
|
|
| ( |
|
| ZRL20234323 |
|
|
|
| ( |
|
| ZRL20234352 |
|
|
|
| ( |
|
| ZRL20232065 |
|
|
|
| ( |
|
| ZRL20232816 |
|
|
|
| ( |
|
| ZRL20232057 |
|
|
|
| ( |
|
| ZRL20235282 |
|
|
|
| ( |
|
| ZRL20233848 |
|
|
|
| ( |
|
| ZRL20232533 |
|
|
|
| ( |
|
| ZRL20233576 |
|
|
|
| ( |
|
| ECV1784 |
|
|
|
| ( |
|
| ZD16070520 |
|
|
| — | (Vellinga. 2011) |
|
| HTBM0275 |
|
| — | — | ( |
|
| HTBM1402 |
|
|
| ( | |
|
| ZRL20232285 |
|
|
|
| ( |
|
| ZRL20235328 |
|
|
|
| ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| HMAS88854 |
|
|
| — | ( |
|
| HTBM0578 |
|
| — | — | ( |
|
| HTBM0689 |
|
|
|
| ( |
|
| 10440 4 |
| — | — | — | (Tian. 2014) |
|
| ZRL20234956 |
|
|
|
| ( |
|
| ZRL20236943 |
|
|
|
| ( |
| ZRL20233912 |
|
| — | — | ( | |
| ZRL20234966 |
|
| — | — | ( | |
|
| HTBM0679 |
|
| — |
| ( |
|
| HTBM0836 |
|
| — | — | ( |
|
| HTBM0958 |
|
| — | — | ( |
|
| HTBM1779 |
|
| — | — | ( |
|
| KaiR1768 |
|
|
| — | ( |
|
| SeSa7 |
|
|
| — | ( |
| 1 | Pileus yellow, with golden droplets exuding |
|
| – | Pileus without droplets exuding |
|
| 2 | Annulus with orange or pink-purplish distinct rim |
|
| – | Annulus white or not white |
|
| 3 | Pileus central dark brown to blackish brown |
|
| – | Pileus central pale yellow, orange, red, brown, or concolorous margin |
|
| 4 | Pileus with depressed dark brownish to grayish brown squamules; stipe covered with purplish brown flocculent fibers; annulus easily detachable |
|
| – | Pileus covered with blackish brown to fuliginous warted or felted squamules; stipe nearly glabrous; annulus white, fugacious |
|
| 5 | Pileus smooth to glabrous, occasionally with radial fibrils |
|
| – | Pileus with squamules or fibrils |
|
| 6 | Pileus with radial streaks towards the margin or fragile |
|
| – | Pile margin entire edges without streaks |
|
| 7 | Pileus centre yellowish orange, margin with radial streaks; basidia clavate |
|
| – | Pileus center slightly yellowish, margin ruptured and fragile; cheilocystidia narrowly clavate |
|
| 8 | Stipe enlarged to subclavate towards base and ranges from cylindrical |
|
| – | Stipe cylindrical or slightly expanded at the base |
|
| 9 | Pileus covered with orange-brown or red-brown floccose squamules; annulus with serrulate margin |
|
| – | Pileus covered with radially arranged light brownish orange to yellowish-orange fibrils or squamules |
|
| 10 | Pileus umbo becoming radially fissured from the center upon maturation, covered with imbricate squamules and adiating fibrillose vestiges |
|
| – | Pileus umbo entire, not issured |
|
| 11 | Pileus reddish brown to yellowish brown; cheilocystidia narrowly clavate, often wavy in shape |
|
| – | Pileus centre yellowish to light brownish, elsewhere with radially arranged light brownish squamules; cheilocystidia apex often sub-capitate to capitate |
|
| 1 | Basidiocarps small |
|
| – | Basidiocarps large |
|
| 2 | Pileus or context discoloring when bruised |
|
| – | Pileus or context not discoloring when bruised |
|
| 3 | Pileus discoloring dark green when bruised |
|
| – | Pileus discoloring oxide red when bruised |
|
| 4 | Stipe bulbous at base |
|
| – | Stipe not bulbous at base |
|
| 5 | Pileus center dark reddish brown |
|
| – | Pileus center not dark reddish brown |
|
| 6 | Cheilocystidia with refractive contents |
|
| – | Cheilocystidia without refractive contents | 7 |
| 7 | Pileus purplish |
|
| – | Pileus not purplish |
|
| 8 | Context discoloring purplish when drying |
|
| – | Context not discoloring when drying |
|
| 9 | Pileus with dark brown umbo; stipe base faint lilacinous tinge. |
|
| – | Pileus with dark umbo; spores with germ pore |
|
| 10 | Cheilocystidiapyriform; lamellae margin light brownish |
|
| – | Cheilocystidia not pyriform |
|
| 11 | Pileus with farinaceous veil |
|
| – | Pileus without veil |
|
| 12 | Pileus margin ruptured; covered with light brownish scales |
|
| – | Pileus margin not ruptured |
|
| 13 | Cheilocystidia have refractive contents at the apex |
|
| – | Cheilocystidia without refractive contents at the apex |
|
| 14 | Pileus with green-gray scales; cheilocystidia apex with crystals |
|
| – | Pileus with orange brown to red brown floccose scales |
|
| 15 | Spores ovoid |
|
| – | Spores not ovoid |
|
| 16 | Pileus white, margin incurved |
|
| – | Pileus with dark reddish-brown scales |
|
| 17 | Annulus margin brownish black to black |
|
| – | Annulus white, pinkish to pinkish-tan when aging |
|
| 18 | Pileus bright yellow |
|
| – | Pileus not bright yellow |
|
| 19 | Pileus truncated campanulate |
|
| – | Pileus not truncated campanulate |
|
| 20 | Basidia spheropedunculate; cheilocystidiasubfusiform and mucronate |
|
| – | Basidia not spheropedunculate |
|
| 21 | Pileus center dark brown; pileipellis with yellow hyphae |
|
| – | Pileus center not dark brown |
|
| 22 | Pileus smooth; annulus yellowish white |
|
| – | Pileus without adnate scales |
|
| 23 | Annulus peronate |
|
| – | Annulus not peronate |
|
| 24 | Pileus very fragile, grooved from the margin to the center |
|
| – | Pileus not very fragile |
|
| 25 | Cheilocystidia apex with tiny crystals |
|
| – | Cheilocystidia apex without tiny crystals |
|
| 26 | Pileus with golden yellow, reddish brown watery exudates |
|
| – | Pileus without watery exudates |
|
| 27 | Pileus with dirty brown scales; stipe base reddish brown |
|
| – | Pileus without dirty brown scales |
|
| 28 | Cheilocystidia narrowly clavate |
|
| – | Cheilocystidia not narrowly clavate |
|
| 29 | Lamellae pale yellow, cheilocystidia lageniform |
|
| – | Lamellae not pale yellow |
|
| 30 | Pileus with green radially scales; cheilocystidia with crystals inside |
|
| – | Pileus scales not green |
|
| 31 | Stipe with pale brown scales below anulus |
|
| – | Stipe without scales below anulus |
|
| 32 | Cheilocystidia fusiform |
|
| – | Cheilocystidia ventricose, fusiform, lageniform, and some rostrate |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Fungal Biology and Applications
Introduction
Locquin first classified Leucoagaricus as a subgenus under Leucocoprinus Pat (Locquin 1943). In 1948, Singer elevated Leucoagaricus togenus with a formal Latin diagnosis (Singer 1948). Candusso and Lanzoni (1990) later reclassified the Leucoagaricus into two subgenera viz., subgenus Sericeomyces and subgenus Leucoagaricusm, and six sections viz., sect. Cystidiosi Migl. & Testoni, sect. Intermedii (Bon) Consiglio & Contu, sect. Leucoagaricus Locq. ex Singer, sect. Piloselli Singer, sect. Pseudopiloselli Migl. & Coppola, and sect. Rubrotincti Singer (Bon 1981). Leucoagaricus is characterized by the small to medium-sized, thin or fleshy basidiomata; pileus surfaces range from radially fibrillose, floccose, squamulose to fibrillose-scaly or rarely granulose; entire or very short striated margins; central, equal to bulbous stipe that have membranous, sometimes fugaciousannulus; thin-walled and smooth basidiospores generally lack well-defined germ pores; and the pileipellis is either a trichoderm or a cutis of repent and radially arranged hyphae lacking clamp connections absent; spore print white, sordid cream, or cream-orange; and metachromatic in Cresyl Blue (Singer 1986; Vellinga 2001). The majority of Leucoagaricus species have been documented from temperate regions in North America and Europe, whereas only a limited number have been reported from tropical areas (Liang et al. 2010; Ishaq et al. 2025).
Patouillard segregated Leucocoprinus Pat from Lepiota, but lacked an original circumscription. Locquin (1943) later provided a comprehensive systematic description of Leucocoprinus and subdivided it into three subgenera: Leucocoprinus, Leucoagaricus, Hiatula (Fr.) Mont., and Leucobolbitius J.E. Lange ex Locq. The genus Leucocoprinus is primarily distributed in tropical and subtropical regions (Vellinga 2001, 2004, 2011; Ge et al. 2015; Liang et al. 2010; Vellinga et al. 2010). Leucocoprinus is a highly diverse genus, with the majority of its species occurring in tropical regions (Vellinga 2004). In recent years, numerous new species have been described from these areas (Ge et al. 2015; Liang et al. 2010; Malysheva et al. 2013; Li et al. 2025). In 2001, Vellinga defined the diagnostic characteristics of Leucocoprinus as follows: basidiospores are metachromatic, clamp connections are absent, pseudoparaphyses are present around the basidia, and the pileus has a plicate margin (Vellinga 2001). Subsequently, most researchers have consistently used these characteristics to define species within Leucoagaricus (Justo 2020, 2021).
The Leucoagaricus and Leucocoprinus exhibit a cosmopolitan distribution in Europe (Candusso and Lanzoni 1990), Pakistan (Ashraf et al. 2023), and Laos (Sysouphanthong 2018). Currently, Leucoagaricus comprises more than 130 species and Leucocoprinus 250 species globally, based on the legitimate names in the MycoBank (https://www.mycobank.org) and the Species Fungorum database (www.speciesfungorum.org; accessed September 24, 2025). To date, a total of 12 Leucoagaricus and 33 Leucocoprinus species have been reported in China, such as: Leucoagaricus atrosquamulosus Z. W. Ge & Zhu L. Yang (Yang 2017), Leucoagaricus exannulatus Singer (Dong 2013), Leucoagaricus lateritiopurpureus E. F. Malysheva, T. Yu. Svetasheva & E. M. Bulakh (Fan 2022), Leucoagaricus bulbosus R. L. Zhao & J. X. Li, Leucoagaricus xantholepis R. L. Zhao & J. X. Li, Leucoagaricus testaceumbonatusR. L. Zhao & J. X. Li, Leucoagaricus luteocanus R. L. Zhao & J. X. Li, Leucoagaricus cinereibisporusR. L. Zhao & J. X. Li, Leucoagaricus centrobrunneolus R. L. Zhao & J. X. Li, Leucoagaricus subcandidus R. L. Zhao & J. X. Li, Leucoagaricus subnivalis R. L. Zhao & J. X. Li (Li et al. 2025), Leucocoprinus brunneoruberJ. M. Zhang & X. T. Zhu (Zhu 2024), Leucocoprinus coerulescens J. F. Liang, Zhu L. Yang & J. Xu (Liang et al. 2010), Leucocoprinus shixingensis (Z.S. Bi & T.H. Li) Kun L. Yang, Jia Y. Lin & Zhu L. Yang (Yang et al 2024), and Leucocoprinus subcretaceus Bon (Chu 2014).
In the current taxonomic revisions, sous-genus Leucoagaricus and sous-genus Sericeomyces are still placed within Leucoagaricus; the remaining related taxa have been reclassified into Leucocoprinus (Yang et al. 2024). At present, the phylogenetic relationships between Leucocoprinus and Leucoagaricus remain unresolved. Previous studies have primarily focused on ITS and nrLSU analyses, resulting in incomplete molecular data; therefore, this study employs ITS, nrLSU, rpb2, and tef1-α sequences. In this study, two new species of Leucoagaricus and Leucocoprinus from China are described based on both morphological and molecular data. The abbreviations for the generic names referenced in this study are as follows: L. = Lepiota, La. = Leucoagaricus, and Lc. = Leucocoprinus.
Materials and methods
Collection of specimens
The study area is located in the northeastern part of China, where we found two new species. The species of Leucoagaricus found in Liaoning Province, specifically in Huludao City, Bailangshan National Nature Reserve, grows on soil and is solitary in deciduous broadleaf forests in August. The species of Leucocoprinus found in Nei Mongol, specifically in Hinggan League City, Arxan National Forest Park, is scattered on the soil in a coniferous-broadleaf mixed forest in September. Fresh basidiocarps were photographed using a Canon 80D digital single-lens reflex (DSLR) camera, with concurrent documentation of habitat characteristics (Rathnayaka et al. 2025). The color characteristics of the basidiocarps were documented and coded following the methodology of Kornerup and Wanscher (1978). All specimens were dried overnight in an electric blast drying oven at 45 °C (Hu et al. 2022) and then deposited in the Herbarium of Mycology at Jilin Agricultural Science and Technology University (HMJU).
Morphological observation
Detailed macro-morphological descriptions of the collected specimens were accomplished from the fresh specimens. Microstructural features, including basidiospores, basidia, cheilocystidia, pleurocystidia, and pileipellis elements, were observed using the protocols described by Largent (1986). The dried specimen sections were processed through fixation with 3% potassium hydroxide (KOH), 1% Congo red, and Melzer’s reagent, and subsequently observed. [n, p, m] indicates: The data represent n basidiospores measured from p samples within m specimens. Basidiospore dimensions are presented in the format ‘(a–) b–av–c(–d)’, where the range ‘b–c’ encompasses at least 90% of measured values. Extreme values a and d are indicated in parentheses, with ‘av’ representing the average. Q denotes the length-to-width ratio of spores, and Qm represents the mean value of Q for all basidiospores. Electron Microscope (SEM) images of basidiospores were obtained from dried, free-hand sections of lamellae, directly mounted on a double-sided adhesive tape pasted onto a metallic specimen stub, and then scanned at different magnifications in high-vacuum mode (Tarafder et al. 2022, 2025). This work was conducted using a Zeiss EVO 18 electron microscope.
DNA extraction, PCR amplification, and sequencing
Total genomic DNA was extracted using the EZup Column Fungi Genomic DNA Purification Kit (Sangon Biotech Co., Ltd., Shanghai, China) according to the manufacturer’s protocol. During PCR amplification, primers ITS1F and ITS4 were employed to amplify the ITS rDNA region (Gardes 1993; White 1990), primers LR0R and LR5 were used to obtain nrLSU sequences (Vilgalys and Hester 1990), primers rpb2-6F and rpb2-7cR were utilized to target the rpb2 gene (Hofstetter et al. 2007; Matheny 2005), and primers TEF1-983F and TEF1-1567R were applied to amplify the tef1-α region (Qu et al. 2023). The PCR cycling protocol comprised: initial denaturation at 95 °C for 5 min, followed by 28 cycles (ITS) or 32 cycles (nrLSU) or 30 cycles (rpb2, tef1-α) of denaturation at 94 °C for 30 s (ITS, tef1-α) or 94 °C for 40 s (nrLSU) or 94 °C for 50 s (rpb2), annealing at 54 °C for 30 s (ITS) or 52 °C for 45 s (nrLSU) or 59 °C for 1 min (rpb2) or 49 °C for 35 s (tef1-α), and extension at 72 °C for 30 s (ITS, tef1-α) or 72 °C for 40 s (nrLSU) or 72 °C for 1 min (rpb2), with a final extension step at 72 °C for 5 min (ITS) or 72 °C for 4 min (nrLSU, tef1-α) or 72 °C for 10 min (rpb2). The PCR products detected by 1% agarose gel electrophoresis using the JY 600 electrophoresis (Beijing JUNYI Electrophoresis Co., Ltd., Beijing, China) apparatus were sent to BGI Co., Ltd. (Beijing, China) for sequencing.
Sequence acquisition and dataset preparation
Use the BLAST algorithm to compare the sequences obtained in this study against the GenBank nucleotide database (ITS, nrLSU, rpb2, and tef1-α). Accession numbers were obtained from GenBank upon submission of nucleotide sequences to the NCBI database. In the phylogenetic analysis, 16 new sequences were generated from two specimens. Integrate the newly generated sequences with existing data from previous phylogenetic studies (Misra et al. 2011; Yang et al. 2024), using Lepiota grangei (Eyre) Kühner and Lepiota felina (Pers.) P. Karst. as the outgroup (Sarawi 2025) to construct a combined data matrix.
Sequence alignment and phylogenetic analyses
To confirm the taxonomic positions of the new species, ITS and nrLSU, rpb2, and tef1-α, sequences were combined and analyzed with Bayesian inference (BI) and maximum likelihood (ML) methods. Sequence alignment was performed using MAFFT v7.0, (Katoh and Standley 2013), and low-quality regions were trimmed with MEGA v7.0 (Kumar et al. 2016) and then combined by Phylosuite v1.2.3 (Zhang et al. 2020; Xiang et al. 2023). The BI analysis was conducted using MrBayes v3.2.7a (Ronquist et al. 2012) in Phylosuite v1.2.3, which employs a Markov chain Monte Carlo (MCMC) algorithm. Nucleotide substitution models were determined by ModelFinder v 2.2.0 (Kalyaanamoorthy et al. 2017). Four Markov chains were run simultaneously for 1,000,000 generations, with the trees sampled every 1000 generations. A 75% majority rule consensus tree was computed after excluding the first 25% trees as “burn-in”. Bayesian inference posterior probability (BIPP) was determined from the remaining trees. The ML analysis was performed using IQ-Tree v2.2.0 (Nguyen et al. 2015) in Phylosuite v1.2.3, with the best model selected for each locus according to ModelFinder. The model selection was performed using ModelFinder under the Bayesian Information Criterion (BI) (Kalyaanamoorthy et al. 2017), with the best model being GTR+F+I+G4.
The phylogenetic position of the new species was inferred using Maximum Likelihood (ML) and Bayesian Inference (BI) methods. The phylogenetic analysis was performed using IQ-TREE for Maximum Likelihood (Minh et al. 2020) and Markov Chain Monte Carlo (MCMC) methods, with MrBayes 3.2.2 (Ronquist et al. 2012). Bootstrap values ≥ 70%in Maximum Likelihood (ML) analysis and Bayesian posterior probabilities (PP) ≥ 0.90 are considered to provide significant support for phylogenetic nodes. The sequences used in this study are listed in Table 1.
Results
Phylogenetic analyses
The final dataset included 16 newly generated sequences and 153 sequences retrieved from the GenBank database. Two sequences of Lepiota grangei and L. felina from GenBank were selected as outgroups. The ML and BI analyses yielded the same topologies, with maximum likelihood bootstrap percentage (MLBP) values on the left and Bayesian posterior probability (PP) values on the right; thus, only the ML tree is presented (Fig. 1). In the resulting phylogeny, the Chinese specimens clustered into two distinct clades with moderate to strong statistical support, indicating that they represent two undescribed species. Within Leucoagaricus, the newly generated ITS, nrLSU, rpb2, and tef1-α sequences of La. bailangshanensis (HMJU 9098) formed a sister relationship to La. centrobrunneolus (ZRL20234707) with 100% ML and 1.00 BYPP statistical support (Fig. 1). Within Leucocoprinus, the ITS, nrLSU, rpb2, and tef1-α sequences of Lc. ferrugineus (HMJU 745) grouped as sisters to Leucocoprinus beijingensis R. L. Zhao & J. X. Li (ZRL20235282) and Leucocoprinus purpurascens T. Guo & Z. W. Ge (ZRL20234956), though this relationship was supported only by moderate ML values of 85% and unsupported in the Bayesian analysis.
Phylogram generated from maximum likelihood analysis based on ITS, LSU, rpb2, and tef1-α sequences data representing Leucoagaricus and Leucocoprinus species. Related sequences are obtained following Li et al. (2025) and GenBank. The tree topology of the ML analysis is similar to the Bayesian analysis. Bootstrap values for ML equal to or greater than 70% and clade credibility values greater than 0.90 (rounded to 2 decimal places) from BYPP analysis are labeled on the nodes. A newly generated sequence is in blue bold, and the type specimen sequences are represented in T.
Taxonomy
Leucoagaricus
bailangshanensis
Taxon classificationFungiAgaricalesAgaricaceae
J.Z. Xu sp. nov.
B8B0CBFD-01DD-563B-833D-F928E496A5AC
Fungal Names: FN 573018
Diagnosis.
Leucoagaricus bailangshanensis differs from La. centrobrunneolus by pileus margin with floccules, longer sterigmata (3.5 µm), and distinctive ITS, nrLSU, rpb2, and tef1-α sequences and position in the phylogram.
Leucoagaricus bailangshanensis (HMJU 9098, holotype). A, B. Basidiocarps; C. SEM images of basidiospores; D. Basidiospores; E. Basidia; F. Cheilocystidia; G. Pileipellis. Scale bar: 2 cm (A, B); 2 µm (C); 5 µm (D, E); 10 µm (F, G).
Holotype.
China • Liaoning Province, Huludao City, Bailangshan National Nature Reserve, on soil, 8 August 2023, J. Z. Xu (HMJU 9098, holotype).
Etymology.
The epithet “bailangshanensis” refers to the location ‘Bailang shan of Liaoning Province, where the holotype was collected.
Description.
Pileus 33–48 mm diam, ranging from plano-convex to convex, pale grayish-white to snow-white (28B1-28A1), with an irregular margin exhibiting remnants of the partial veil, surface arid, bearing imbricate squamules and radiating white fibrillose vestiges; umbo slightly obtuse, vinaceous yellow (4B4) centrally positioned, becoming radially fissured from the pileus center upon maturation. Context white, non-discoloring when bruised, thin. Lamellae free, cream-white (1A2) to white (1A1), crowded, less than 1 mm in width, with 1–3 tiers of lamellulae; edge entire. Stipe 41–54 × 3–5 mm, birch bark (6B2), hollow, covering white fibrillose. Annulus superior and single-layered white (1A1) on the upper stipe. Odor not distinctive. Spore print white.
Basidiospores [40/4/2] (3.6–)4.7–5.6–6.4(–6.9) × (2.8–)3.1–3.9–4.5(–4.8) µm, Q=1.2–1.8, Qm=1.40, broadly ellipsoid to slightly elongated, without a germ pore, transparent in 3% KOH, dextrinoid, with 1 or 2 guttules. Basidia (12–)13–14.8–17(–17.7) × (5.0–)6.5–7.3–8.1(–8.7) µm, 2–4 spored, broadly clavate, hyaline in KOH; sterigmata up to 3.5 µm long. Cheilocystidia (23.1–)23.4–29.4–37.4(–39.4) × (7.3–)9.0–11.4–13.7(–15.0) μm, subfusiform to fusiform, smooth and hyaline KOH. Pleurocystidia absent. Lamellatrama regular, made up of parallel to subparallel, 2.6–8.1 μm wide hyphae in KOH. Pileipellis a cutis of repent, subcylindrical, radially arranged, occasionally branched, trichodermal, hyphae 3.6–14.2 wide. Clamp connections absent.
Habitat.
Solitary on soil in deciduous broadleaf forests.
Known distribution.
Known only from north-eastern China.
Additional material examined.
China • Liaoning Province, Huludao City, Bailangshan National Nature Reserve, on soil, 8 August 2023, J.Z. Xu (HMJU 9304); China • Jilin Province, Jilin City, Zuojia Nature Reserve on soil, 22 September 2023, J.Z. Xu (HMJU 9842).
Notes.
The species is characterized by a pileus with a vinaceous, yellow umbo and white squamules radially arranged over a white background, crowded lamellae, a pileus margin with floccules, and cheilocystidia subfusiform to fusiform; and ITS, nrLSU, rpb2, tef1-α sequence analyses, the present species clusters with members of the La. centrobrunneolus, signifying its position. Regarding overall morphology, the present specimen is quite similar to La. centrobrunneolus, La. lateritiopurpureus, La. Goossensiae Heinem, Leucoagaricus fuligineus Pegler, Leucoagaricus griseus Heinem, and La. luteocanus. However, La. centrobrunneolus has a much smaller pileus (20 mm) with its surface covered without floccules; smaller sterigmata (1.5 µm) (Li et al. 2025). The pileus of La. lateritiopurpureus (10–20 mm) is covered with pink-brownish, brightly orange-brown, or terracotta scales, and much bigger basidia (20–25 × 7–10 µm) (Malysheva et al. 2013). La. goossensiae and La. bailangshanensis differ in that the pileus is reddish-brown, smoother, and with radiating grooves; and the stipe is longer (Heinemann 1973). Leucoagaricus fuligineus has a pileus the center of which is grayish-brown; the hyphae of the pileipellis are grayish-brown; basidia are bigger; and spores are concave (Pegler 1977). Leucoagaricus griseus has a pileus, the center of which is dark gray, with much larger basidia and lanceolate cheilocystidia (Heinemann 1979). Leucoagaricus luteocanus has a pileus the center of which is light brown, with radial light-brown to pale-yellow squamulose or fibrillose; the stipe has uneven brown coloration.
Leucocoprinus
ferrugineus
Taxon classificationFungiAgaricalesAgaricaceae
J.Z. Xu sp. nov.
1151C4BA-1D5C-5CA9-B2CC-0F37AE06F658
Fungal Names: FN 573019
Diagnosis.
Leucocoprinus ferrugineus differs from Lc. purpurascens by the absence of cheilocystidia, basidiospores amygdaliform, smaller basidia (14.7– 21.5 × 6.4–9.5 µm) (Guo et al. 2023) and distinctive ITS, nrLSU, rpb2, and tef1-α sequences and position in the phylogram.
Leucocoprinus ferrugineus (HMJU 745, holotype). A, B. Basidiocarps; C. SEM images of basidiospores; D. Basidiospores; E. Basidia; F. Cheilocystidia; G. Pileipellis. Scale bar: 2 cm (A, B); 2 µm (C); 5 µm (D, E); 10 µm (F, G).
Holotype.
China • Nei Mongol, Hinggan League City, Arxan National Forest Park, on soil, 1 September 2020, J. Z. Xu (HMJU 745, holotype).
Etymology.
The species epithet “ferrugineus” is derived from the Latin word “ferruginous”, referring to the ferruginous red pileus at the center of the taxon.
Description.
Pileus 12–35 mm diam, convex-lens, mahogany-red (8E7) blunt umbo at center and small mahogany-red (8E7) squamules in white base color; margin uneven oxide-red (8E8). Context white, discoloring to oxide red (8E8) when bruised, thick. Lamellae free, white with densely radiating tomato-red (8C8) spots, less than 1 mm in width with 1–3 tiers of lamellulae; edge entire. Stipe 35–50 × 2–3 mm, cylindrical with a swollen base, white in the lower part, gradually deepening to fox (8D7). Annulus at the upper middle portion of the stipe, white, with a mahogany-red (8E7) margin.
Basidiospores [40/4/2] (5.9–)6.8–7.5–8.5(–8.9) × (4.1–)4.3–4.9–5.6(–6.2) µm, Q=1.4–1.9, Qm=1.54, broadly ellipsoid to slightly elongated, without germpore, transparent in 3% KOH, dextrinoid, with 1 or 2 guttules. Basidia (13.1–)14.7–18.1–21.5(–21.8) × (6.3–)6.4–7.9–9.5(–10.4) µm, 2–4 spored, broadly clavate; sterigmata up to 2.5 µm long, hyaline in KOH. Cheilocystidia (28.0–)34.0–44.7–59.8(–71.2) × (11.1–)12.2–16.5–21.3(–28.2) µm, capitate to subcapitate, smooth and hyaline in KOH. Pleurocystidia absent. Lamellatrama regular, made up of parallel to subparallel, 3.26–12.53 μm wide, hyaline in KOH. Pileipellis a cutis of repent, subcylindrical, radially arranged, occasionally branched, trichodermal, seen hyphae 3.0–13.5 μm wide, transparent in 3% KOH. Clamp connections absent.
Habitat.
Solitary on the soil in coniferous-broadleaf mixed forest.
Known distribution.
Known only from north-eastern China.
Additional material examined.
Nei Mongol • Hinggan League City, Arxan National Forest Park, on soil, 1 September 2020, J. Z. Xu. (HMJU 898)
Notes.
The species is characterized mainly by mahogany-colored squamules at the center, stipe fox-brown in median to lower portions, deepening in color toward the base, annulus white with a mahogany-colored margin, context turns oxide red upon injury, and the spores are ellipsoid and ITS, nrLSU, rpb2, tef1-α sequence analyses, the present species cluster with members of the Lc. beijingensis and Lc. purpurascens signifies its position. Among morphologically related species, the present specimen is somewhat similar to species such as Lc. purpurascens, Leucocoprinus lahorensis Asif, Saba & Vellinga, Leucocoprinus antillarum Justo, Bizzi, Angelini, Leucocoprinus brunneocanus, Leucocoprinus brunneus. Among morphologically related taxa, Lc. purpurascens differs in lacking cheilocystidia and pileipellis hyphae, which are typically yellowish-brown (Guo et al. 2023). Leucocoprinus antillarum has larger spores, cheilocystidia that are subfusiform, and basidiomata that are uniformly white. Leucocoprinus brunneocanus Asif, Saba & Vellinga has gray squamules of pileus; context un-discoloring when bruised; and cheilocystidia sometimes constricted or curved (Asif 2024). Leucocoprinus brunneus differs by having a thicker stipe; cheilocystidia are smaller and clavate.
Discussion
Phylogenetic analyses based on the combined ITS, nrLSU, rpb2, and tef1-α dataset show a monophyletic clade of Leucoagaricus and Leucocoprinus taxa, including Lepiota as outgroup (Fig. 1). Detailed morphological features, along with phylogenetic studies, support the La. bailangshanensis and Lc. ferrugineus as new species. Leucoagaricus bailangshanensis shows a close phylogenetic affinity with La. centrobrunneolus, originally described from China (ITS: GenBank PP816117); however, La. centrobrunneolus is characterized by a much smaller pileus, larger basidia, and smaller sterigmata. Leucocoprinus ferrugineus exhibits the closest phylogenetic affinity to Lc. beijingensis (ITS: GenBank PP816074; Li et al. 2025), and Lc. purpurascens described from China (ITS: GenBank OM987458; Yang et al. 2024). Leucocoprinus beijingensis differs from Lc. ferrugineus by possessing pyriform cheilocystidia and a context that does not change color when damaged (Zhang et al. 2020). Leucocoprinis purpurascens differs in lacking cheilocystidia and possessing yellowish-brown hyphae in the pileipellis.
To date, a total of 12 species of Leucoagaricus and 33 species of Leucocoprinus have been reported in China. Based on the observations of characteristic keys and literature, we prepared a simple taxonomic key including the newly described species and the other Leucoagaricus and Leucocoprinus species from China.
An artificial key to the species of Leucoagaricus reported from China
**: **
An artificial key to the species of Leucocoprinus reported from China
**: **
Supplementary Material
XML Treatment for Leucoagaricus bailangshanensis
XML Treatment for Leucocoprinus ferrugineus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ashraf S Naseer A Usman M Khalid AN (2023) Two new species of genus Leucoagaricus (Agaricaceae, Agaricales) from Pakistan.Myco Keys 96: 159–171. 10.3897/mycokeys.96.10174537214178 PMC 10196934 · doi ↗ · pubmed ↗
- 2Asif M Saba M Raza M Vellinga EC (2024) Molecular insights into fungal diversity reveal three novel species of Leucocoprinus from southern Punjab, Pakistan.Mycologia 116(4): 601–620. 10.1080/00275514.2024.235176938847769 · doi ↗ · pubmed ↗
- 3Bon M (1981) Clé monographique des “Lépiotes” d’Europe.Documents Mycologiques 11(44): 1–77.
- 4Candusso M Lanzoni G (1990) Fungi Europaei 4. Lepiota s.l..Giovanna Biella, Saronno, 743 pp.
- 5Chu HL (2014) Taxonomy and Molecular Phylogeny of Macrofungi and Related Fungi in Hainan and Guizhou. Ph D Thesis. Hainan University, Hainan, Haikou.
- 6Dong D (2013) Researches on resources of Leucocalocybe mongolicum and its genetic diversity. Jilin Agricultural University, Jilin, Changchun.
- 7Fan D (2022) Studies on Macrofungi Diversity and Floristic Characteristics in Yuhe Nature Reserve. Northwest Normal University, Gansu, Lanzhou.
- 8Gardes M Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes‐application to the identification of mycorrhizae and rusts.Molecular Ecology 2(2): 113–118. 10.1111/j.1365-294X.1993.tb 00005.x 8180733 · doi ↗ · pubmed ↗