Imperatorin Alleviates Intestinal Fibrosis by Suppressing AIM2-mediated GSDMD Pyroptosis in Macrophages
Sheng Li, Fangqing Zhu, Yao Xie, Teng Ben, Ke Liu, Xinlong Lin, Qian Zhou, Yin Zhang, Xinyue Zhang, Yeling Chen, Yuexin Ren, Xianfei Wang, Fachao Zhi, Gao Tan

TL;DR
Imperatorin reduces intestinal fibrosis in Crohn's disease by blocking AIM2-driven pyroptosis in macrophages.
Contribution
Identifies imperatorin as a novel inhibitor of AIM2-mediated GSDMD pyroptosis in intestinal fibrosis.
Findings
CD patient tissues show increased GSDMD-NT+ CD11b+ macrophages in stenotic ileocecal valves.
Imperatorin reduces intestinal fibrosis and inhibits AIM2, Caspase-1, and GSDMD cleavage in mice.
In vitro, imperatorin suppresses AIM2 inflammasome activity in macrophages.
Abstract
Over-activation of pyroptosis, recently reidentified as Gasdermin D (GSDMD)-mediated proinflammatory cell death, results in severe inflammation-related disorders. Intestinal fibrosis, an inflammation-related disorder, remains one of the most common and intractable complications of Crohn’s disease (CD). However, it is unknown whether excessive pyroptosis contributes to the development of intestinal fibrosis in CD. Immunofluorescence costaining of CD11b and the pyroptosis-inducing fragment GSDMD-N terminal (GSDMD-NT) was performed in stenotic ileocecal valve tissues from patients with CD. A 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced mouse CD model was established. J744a.1 macrophages pretreated with imperatorin (IMP) were transfected with lipopolysaccharides (LPS) plus poly (dA: dT) to explore potential regulatory mechanisms controlling GSDMD-mediated pyroptosis in vitro.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1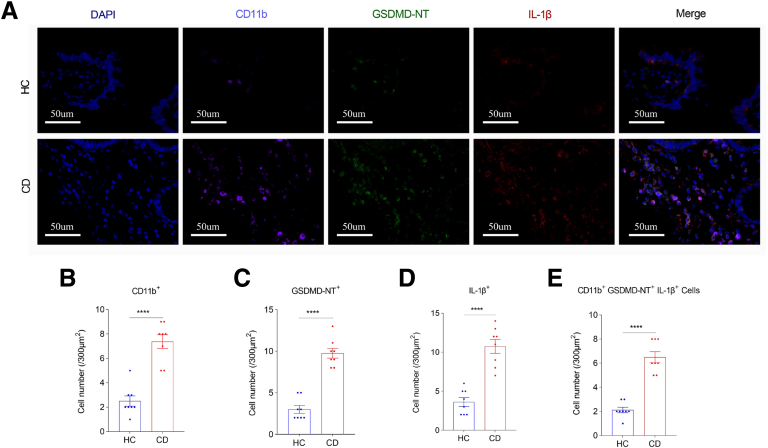 Figure 2
Figure 2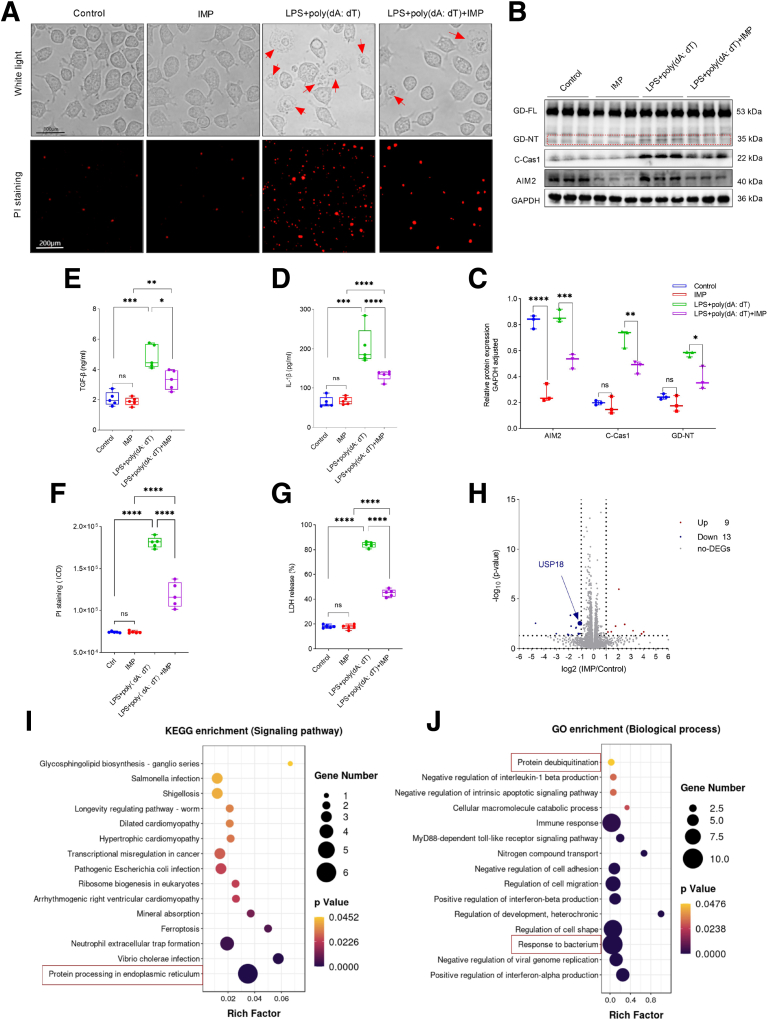 Figure 3
Figure 3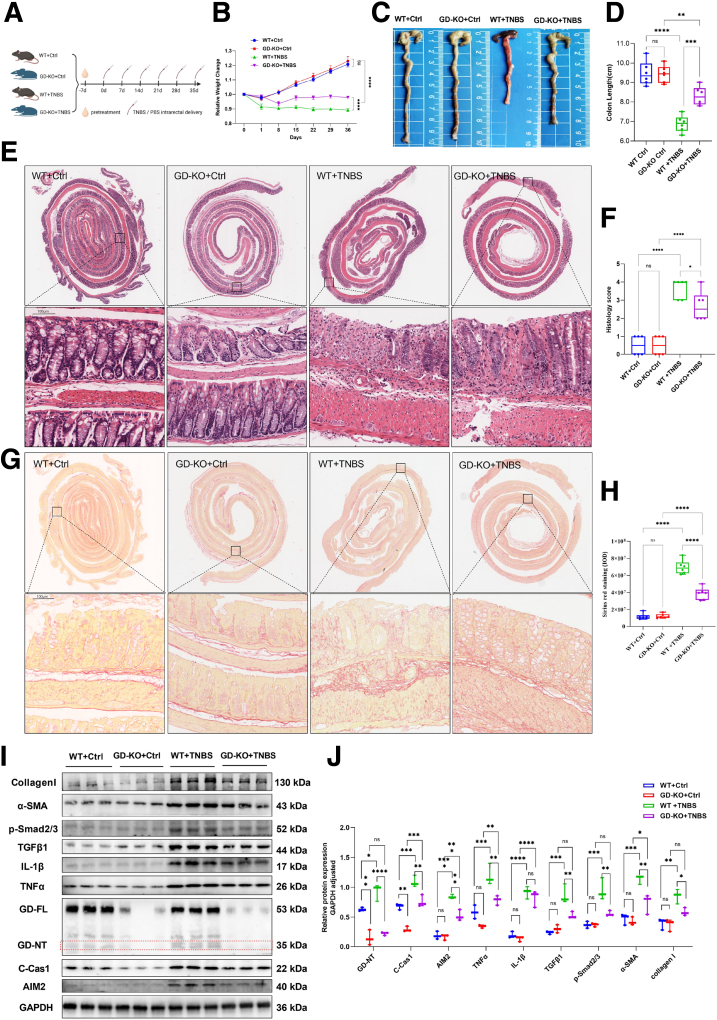 Figure 4
Figure 4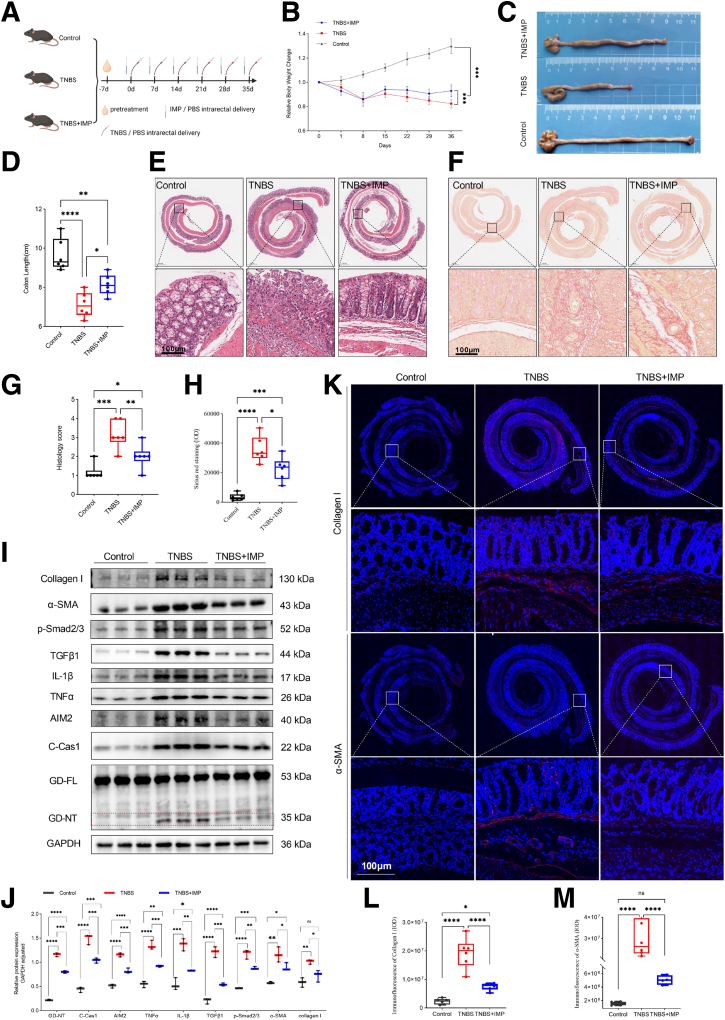 Figure 5
Figure 5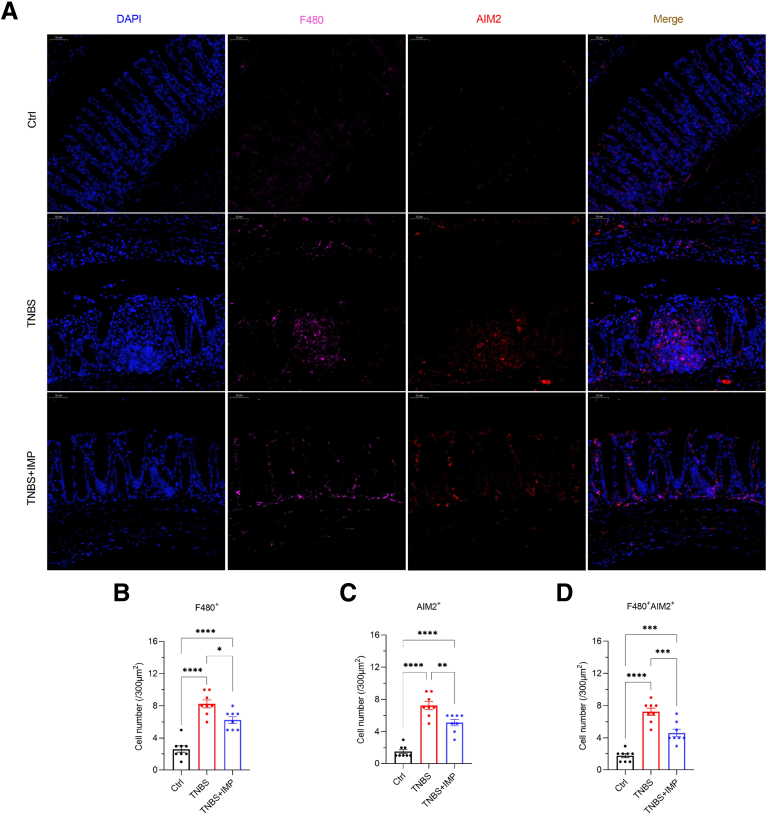 Figure 6
Figure 6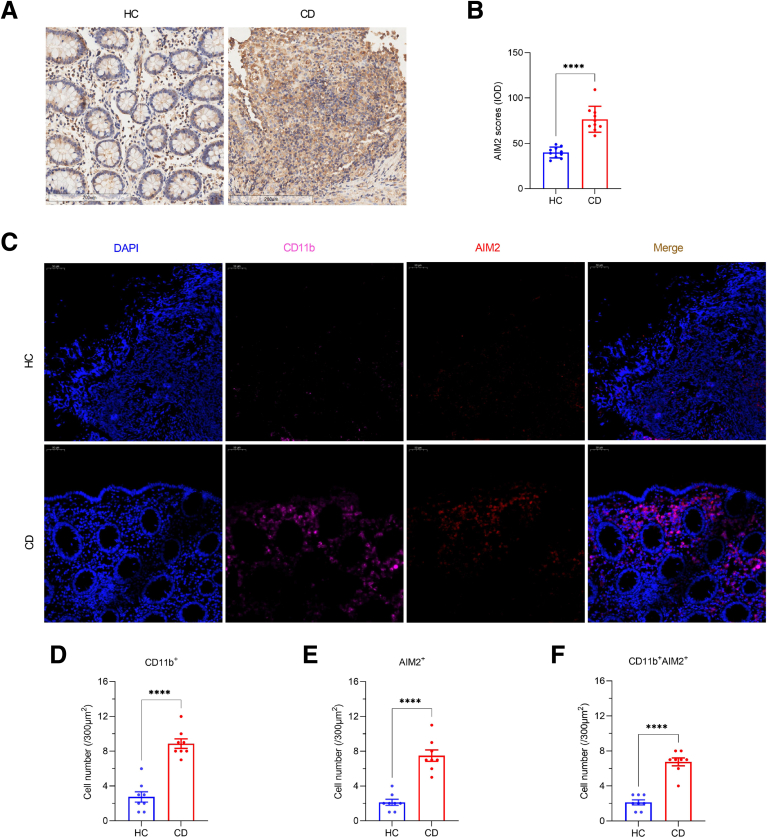 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInflammasome and immune disorders · IL-33, ST2, and ILC Pathways · Eosinophilic Esophagitis
SummaryWe discovered imperatorin (IMP), a natural compound, alleviates Crohn’s intestinal fibrosis by blocking AIM2-mediated pyroptosis in macrophages. Validated in human tissues and animal models, IMP enema offers a novel targeted therapy for stricturing complications.
Crohn’s disease (CD) is a kind of refractory chronic inflammatory bowel diseases (IBD) of unknown etiology.1 With increasing incidence worldwide, it has become a global public health problem.2 In the early stages, its main manifestation is intestinal inflammation, whereas in the middle and late stages, with repeated episodes of intestinal inflammation, it gradually develops into intestinal strictures.3 After 10-year diagnosis of CD, approximately 70% of patients will develop intestinal strictures, most of which require endoscopic balloon dilation and surgery, despite a high recurrence rate.4 The overall prognosis of patients with CD with intestinal strictures is extremely poor.5 The essence of intestinal strictures is intestinal fibrosis, but specific antifibrotic drugs are currently unavailable.3 Although intestinal inflammation in patients with CD has been somewhat controlled in clinical treatment with the rise of immunosuppressants and biologics, these drugs fail to halt fibrotic progression.6 Thus, elucidating fibrogenic mechanisms and identifying novel therapeutics are urgent clinical priorities.
Intestinal fibrosis is characterized by excessive deposition of the extracellular matrix (ECM) in the intestine.3^,^6 To date, known pathological processes responsible for the development of intestinal fibrosis include fibroblast proliferation, migration, recruitment and activation, fibrocyte recruitment, activation and differentiation of stellate cells, pericyte differentiation, EMT, and EndoMT.7 Recurrent inflammation is regarded to be the main cause of intestinal fibrosis in CD, because it participates in these pathological processes from multiple aspects.7, 8, 9 Certain inflammation and consequent fibroblast activation are necessary for normal physiological repair of damaged tissues, whereas recurrent and excessive inflammation will initiate multiple pathological processes that result in dysregulated repair and tissue fibrosis.10 However, what mechanism induces excessive inflammation contributing to the development of intestinal fibrosis is unclear.
Pyroptosis is initially regarded as Caspase1-dependent cell death in response to certain microbial invasion.11 With the recent identification of Gasdermin D-N terminal (GSDMD-NT) as the pyroptotic executioner, pyroptosis is redefined as Gasdermin-mediated pro-inflammatory programmed cell death, characterized by cell membrane swelling and rupture, and distinct from apoptosis, autophagy, ferroptosis, cuproptosis, and necroptosis in morphological characteristics.12, 13, 14, 15, 16, 17 Inflammasomes, including NLRP1, NLRP3, NLRC4, AIM2, and Pyrin, play a vital role in triggering pyroptosis.16 Among them, the AIM2 inflammasome in macrophages sensing microbial DNA activates Caspase1 and subsequently induces Gasdermin D (GSDMD)-mediated pyroptosis, thereby acting as an anti-microbial host defense, but strict negative regulatory mechanisms are required to avoid excessive activation of AIM2-dependent and GSDMD-mediated pyroptosis, which is harmful to the host.18 Currently, it is a well-known fact that patients with CD have an exaggerated intestinal inflammatory response to invaded microbes.19 However, it is unclear whether it is due to impaired negative regulatory mechanisms controlling the AIM2 inflammasome that invading microbes trigger excessive GSDMD-mediated pyroptosis in macrophages, leading to severe intestinal inflammation and fibrosis in CD.
Natural compounds represent promising anti-fibrotic candidates; among them, imperatorin (IMP)—a bioactive furanocoumarin—has been shown to attenuate fibrosis in extra-intestinal organs20 and modulate inflammatory caspases,21 yet its therapeutic potential for intestinal fibrosis remains unexplored. This study aimed to: (1) determine whether AIM2-GSDMD pyroptosis promotes intestinal fibrosis in CD; and (2) evaluate IMP’s efficacy and mechanism of action targeting the AIM2 inflammasome. Through integrated analysis of clinical specimens, murine models, and in vitro systems, we demonstrate that IMP alleviates fibrosis by suppressing AIM2-mediated pyroptosis in macrophages, thereby proposing a novel therapeutic strategy for CD-associated intestinal fibrosis.
Results
GSDMD-mediated Pyroptosis in Macrophages Is Excessively Activated in Intestinal Stenotic Tissues From Patients With CD
To investigate whether GSDMD-mediated pyroptosis in macrophages is associated with CD-associated intestinal fibrosis, we performed triple immunofluorescence staining of CD11b, GSDMD-NT, and interleukin (IL)-1β in stenotic ileocecal valve tissues from patients with CD and in normal ileocecal valve tissues from healthy controls (HCs), and found that CD11b^+^ cells, GSDMD-NT^+^ cells, IL-1β^+^ cells, and CD11b^+^ GSDMD-NT^+^ IL-1β^+^ cells were significantly increased in stenotic ileocecal valve tissues from patients with CD compared with that in normal ileocecal valve tissues from HCs, suggesting that GSDMD-mediated pyroptosis in CD11b^+^ macrophages of intestinal stricture tissues is excessively activated in CD (Figure 1A–E).Figure 1CD11b^+^ GSDMD-NT^+^ IL-1β^+^ macrophages are increased in stenotic ileocecal valve tissues from patients with CD. (A–E) Triple immunofluorescence costaining of CD11b, GSDMD-NT, and IL-1β in stenotic ileocecal valve tissues from patients with CD and in normal ileocecal valve tissues from HCs. (A) Representative immunofluorescence images. Scale bars:50 μm. (B–E) The number of CD11b^+^, GSDMD-NT^+^, and IL-1β^+^ cells per 300 μm^2^. Pyroptotic macrophages were costaining with CD11b (pink), GSDMD-NT (green), and IL-1β (red). CD11b^+^ GSDMD-NT^+^ IL-1β^+^ macrophages were counted under a microscope. Data represent means ± SEM (n = 8 per group); ∗∗∗∗P < .0001 by Student’s t-test.
To further determine whether GSDMD-mediated pyroptosis in macrophages participates in the development of intestinal fibrosis, we first stimulated J774A.1 macrophages with lipopolysaccharide (LPS) plus poly (dA: dT) and then used their culture supernatants to stimulate CCD-18Co fibroblasts in vitro. We observed that J774A.1 cells treated with LPS plus poly (dA: dT) had obvious pyroptotic bubbles, Caspase-1 activation, and GSDMD cleavage, and in their culture supernatants, the concentration of IL-1β was significantly increased (Figure 2A–D), suggesting that LPS plus poly (dA: dT) can induce GSDMD-mediated pyroptosis in macrophages to secrete lots of cytokines, including IL-1β. In addition, we also found that the concentration of transforming growth factor beta (TGF-β) was significantly increased in the culture supernatants from CCD-18Co cells treated with the supernatants of pyroptotic macrophages compared with that from untreated CCD-18Co cells (Figure 2E). These results suggest that GSDMD-mediated pyroptosis in macrophages secrete lots of cytokines, such as IL-1β, to activate intestinal fibroblasts, thereby promoting the occurrence and development of intestinal fibrosis.Figure 2IMP inhibits GSDMD-mediated pyroptosis in macrophages through downregulating AIM2 expression. (A–G) J774A.1 macrophages were treated with or without 50 μM IMP for 6 hours following transfection with 1 μg/μL LPS plus 2.5 ug/mL poly (dA: dT) for 8 hours. Control represents untreated cells. (B, C) Their whole cell lysates were subjected to Western blot (WB) analysis. (B) Representative WB images. Scale bars: 100 μm. Red arrows: pyroptotic bubbles. (C) The numbers present the intensity ratio of indicated proteins/GAPDH analyzed by ImageJ software. (D, G) Their culture supernatants were collected for LDH and IL-1β assays. (E) Their culture supernatants were collected to stimulate the human colon fibroblast cell line CCD-18Co. After 24 hours, CCD-18Co cells were changed to a new medium and cultured for 12 hours, and subsequently their culture supernatants were collected for TGF-β assays. (F) These cells were stained with PI solution (100 μL/well) at room temperature for 15 minutes in the dark, and then observed under a fluorescence microscope. Quantitative analysis of PI staining by ImageJ software. IOD, integrated optical density. (H–J) J744a.1 macrophages were treated with 50 μM IMP for 6 hours, then total RNA was isolated and DEGs were analyzed by RNA-seq. Control represents untreated cells. (H) Volcano plot showing the DEGs in the IMP-treated group vs the untreated control group. Genes were plotted based on log_2_ fold change against the absolute confidence (−log_10_p value). Green and red dots represent down- and up-regulated genes, respectively. (I) KEGG pathway enrichment analysis of the DEGs in the 2 groups. Dot size represents the gene number of DEGs and the corresponding color represents its P value. (J) GO functional enrichment analysis of the DEGs in the 2 groups. Enriched GO terms according to biological processes. (C–G) Data represent means ± SD from 3 to 5 technical replicates; ns, not significant; ∗P < .05, ∗∗P < .01, ∗∗∗P < .001, ∗∗∗∗P < .0001 by 1-way ANOVA test. All data shown are representative of 3 independent experiments.
Gsdmd Deletion Alleviates 2,4,6-trinitrobenzenesulfonic Acid-induced Colitis and Fibrosis in Mice
To determine the role of GSDMD-mediated pyroptosis in CD-associated fibrosis, Gsdmd^-/-^ mice and their wild-type (WT) littermates were rectally injected with 2,4,6-trinitrobenzenesulfonic acid (TNBS) to induce the chronic CD model according to the methods shown in Figure 3A. Compared with WT littermates, Gsdmd^-/-^ mice were resistant to TNBS-induced colitis and intestinal fibrosis, exhibiting less weight loss, colonic shortening, inflammation-associated histological scores, Sirius red staining, and decreased expression of Collagen I, alpha smooth muscle actin (α-SMA), p-Smad2/3, TGFβ1, and tumor necrosis factor alpha (TNFα) in the TNBS colons (Figure 3B-J). In addition, we found that Gsdmd knockout significantly decreased the levels of AIM2 expression, cleaved-Caspase1, and GSDMD-NT in the TNBS colons (Figure 3I, J). Together, these results suggest that Gsdmd-mediated pyroptosis plays an important role in the development of TNBS-induced colitis and intestinal fibrosis.Figure 3Gsdmd^-/-^ mice are less sensitive to TNBS-induced colitis and fibrosis. (A–J) Gsdmd^-/-^ (KO) mice and WT littermates were intrarectally injected with 2.5% TNBS to induce experimental chronic CD model, or with PBS enema as a negative control (Ctrl). On the 36th dday, all the mice were euthanized, and specimens were taken for testing. (B) One day after TNBS or PBS enema, body weight changes were monitored. (C, D) Representative images of the colons. Colon lengths were measured. (E, F) Colon sections were examined histologically. (E) Representative images of the H&E-stained colon sections; black boxes represent enlarged images. Scale bars: 100 μm. (F) Histology scores for colonic inflammation were measured. (G, H) Representative images of Sirius red staining; black boxes represent enlarged images. Scale bars: 100 μm. Quantitative analysis by ImageJ software. (I, J) The levels of indicated proteins in the colonic tissues were analyzed by Western blot (WB). (I) Representative WB images. (J) The numbers present the intensity ratio of indicated proteins/GAPDH analyzed by ImageJ software. (B, D, F, H) Data represent means ± SEM (n = 6 per group); ns, not significant; ∗P < .05, ∗∗P < .01, ∗∗∗P < .001, ∗∗∗∗P < .0001 by 1-way ANOVA test. All data shown are representative of 3 independent experiments.
IMP Alleviates TNBS-induced Colitis and Fibrosis Through Inhibiting GSDMD-mediated Pyroptosis
Traditional Chinese medicine (TCM) is a great treasure house of medicine, in which searching for natural compounds that may suppress excessive pyroptosis has become a potent and valuable approach for inflammation-related disorders, such as fibrosis and cancer. A recent finding shows that IMP, a natural furocoumarin, can alleviate pulmonary fibrosis induced by bleomycin in vivo.20 Another study displays that it can inhibit obesity-induced excessive activation of Caspase-1 in stellate sympathetic ganglion,22 whereas activated Caspase-1 can cleave GSDMD into C- and N-terminal, in which GSDMD-NT, as the pyroptotic executioner, induces proinflammatory cell death.16 Thus, we speculated that IMP may alleviate CD colitis and fibrosis via inhibiting excessive activation of Caspase-1-dependent and GSDMD-mediated pyroptosis. To test it, we rectally injected IMP or phosphate buffered saline (PBS) as a control in the TNBS-induced chronic CD model according to the methods shown in Figure 4A. The IMP-administered WT mice were weakly responsive to TNBS-induced colitis and intestinal fibrosis compared with the controls, displaying reduced weight loss, colonic shortening, inflammation-associated histological scores, Sirius red staining, and downregulated colonic expression of Collagen I, α-SMA, p-Smad2/3, TGFβ1, IL-1β, and TNFα, as well as decreased immunofluorescence staining of Collagen I and α-SMA in the colons (Figure 4B–M). These results suggest that IMP plays a protective role against CD-associated inflammation and fibrosis.Figure 4IMP alleviates TNBS-induced colitis and fibrosis in mice. (A–M) Littermate WT mice were divided into 3 groups. In the Control group, the mice were intrarectally injected with PBS alone. One day before TNBS enema, the mice in the TNBS group were intrarectally injected with 100 μL PBS, whereas the mice in the TNBS + IMP group were injected with 100 μL 3 mM IMP. On the 36th day, all mice were euthanized, and specimens were taken for testing. (B) One day after TNBS or PBS enema, body weight changes were monitored. (C, D) Representative images of the colons. Colon lengths were measured. (E, G) Colon sections were examined histologically. (E) Representative images of the H&E-stained colon sections; black boxes represent enlarged images. Scale bars: 100 μm. (G) Histology scores for colonic inflammation were measured. (F, H) Representative images of Sirius red staining; black boxes represent enlarged images. Scale bars: 100 μm. Quantitative analysis by ImageJ software. (I, J) The levels of indicated proteins in the colonic tissues were analyzed by Western blot (WB). (I) Representative WB images. (J) The numbers present the intensity ratio of indicated proteins/GAPDH analyzed by ImageJ software. (K–M) Representative immunofluorescence images of Collagen Ⅰ and α-SMA immunostaining in the colonic tissues; white boxes represent enlarged images. Scale bars: 100 μm. Quantitative analysis by ImageJ software. (B, D, G, H, L, M) Data represent means ± SEM (n = 6 per group); ns, not significant; ∗P < .05, ∗∗P < .01, ∗∗∗P < .001, ∗∗∗∗P < .0001 by 1-way ANOVA test. All data shown are representative of 3 independent experiments.
Our previous finding showed that disruption of the intestinal epithelial cell (IEC) barrier causes an increase of intestinal mucosal bacteria, thus activating abnormal inflammatory responses in the gut,21 in line with a generally accepted view that an excessive intestinal immune response to the gut microbe in CD is due to disruption of the IEC barrier.19 The AIM2 inflammasome sensing bacterial DNA triggers GSDMD-mediated pyroptosis in macrophages against bacterial invasion, but a strict regulatory mechanism is required to avoid its excessive activation by bacterial DNA, which is harmful to the host.18 Here, we found that the TNBS-challenged mice had obvious epithelium damage and crypt loss, and increased AIM2 expression in the colon (Figure 4E, I, J), suggesting that TNBS caused excessive AIM2 activation upon various intestinal luminal bacteria translocation into the mucosa from disrupted IEC barrier. In addition, TNBS also caused obvious Caspase1 activation and Gsdmd cleavage to produce the pyroptosis-inducing fragment GSDMD-NT (Figure 4I, J). However, IMP significantly decreased the levels of AIM2 expression, cleaved Caspase1, and GSDMD-NT in the TNBS colons (Figure 4I, J). Together, these results indicate that IMP acts as a negative regulatory factor controlling excessive AIM2 and Caspase1 activation and subsequent GSDMD-mediated pyroptosis.
IMP Negatively Controls GSDMD-mediated Pyroptosis in Macrophages Through Inhibiting the AIM2 Pathway
To confirm the role of IMP in GSDMD-mediated pyroptosis, J774A.1 macrophages were pretreated with or without IMP, then transfected with LPS plus poly (dA: dT) in vitro. We observed that the cells transfected with LPS plus poly (dA: dT) had obvious pyroptotic bubbles with disrupted cytomembranes as reflected by increased lactate dehydrogenase (LDH) release and propidium iodide (PI) staining compared with the untransfected cells (Figure 2A, F, G). Moreover, this treatment also caused Caspase1 activation and GSDMD cleavage (Figure 2B, C), indicating that it induces GSDMD-mediated pyroptosis. However, IMP inhibited GSDMD-mediated pyroptosis induced by LPS plus poly (dA: dT), displaying less pyroptotic bubbles, LDH release, PI staining, AIM2 expression, cleaved-Caspase1, and GSDMD-NT in the cells with IMP pretreatment compared with the cells without IMP pretreatment (Figure 2A–C, F, G). Furthermore, IMP alone downregulated AIM2 expression, but not GSDMD or Caspase1 (Figure 2B, C). These results suggest that IMP inhibits GSDMD-mediated pyroptosis through downregulating AIM2 expression in macrophages.
To further validate the mechanism by which IMP alleviates GSDMD-mediated macrophage pyroptosis via AIM2 in mice, we performed double immunofluorescence staining for F4/80 and AIM2 on colon tissues from IMP-treated TNBS-induced colitis (CD) mice. The results demonstrated that, compared with the control group, the TNBS group exhibited a significant increase in F4/80^+^ macrophage density along with elevated AIM2^+^ cell density. Double colocalization analysis revealed a marked increase in F4/80^+^ AIM2^+^ double-positive cells in the TNBS group relative to controls (Figure 5A–D). In contrast, the TNBS + IMP cotreatment group showed reduced F4/80^+^ macrophage density, decreased AIM2^+^ cell density, and fewer F4/80^+^ AIM2^+^ double-positive cells compared with the TNBS group, indicating that IMP mitigates GSDMD-mediated pyroptosis in vivo by downregulating AIM2 expression in macrophages. Taken together, these results suggest that the AIM2 inflammasome emerges as the pivotal protein through which IMP regulates macrophage pyroptosis.Figure 5IMP alleviates GSDMD-mediated pyroptosis through downregulating AIM2 expression in colonic macrophages. (A–D) Double immunofluorescence staining of F4/80 and AIM2 in colon tissues from TNBS-induced colitis mice with/without IMP intervention and control groups. (A) Representative confocal micrographs showing costaining patterns. Scale bar: 50 μm. (B–D) Quantitative analysis of immunopositive cells within 300 μm^2^ regions. (B–E) Data represent means ± SEM (n = 10 per group); ∗P < .05, ∗∗P < .01, ∗∗∗P < .001, ∗∗∗∗P < .0001 by 1-way ANOVA test.
AIM2 Upregulation in Macrophages Associates With Intestinal Fibrosis in CD
To further elucidate the role of AIM2 in the pathogenesis of CD-associated intestinal fibrosis, we performed immunohistochemical (IHC) and immunofluorescence staining of AIM2 in stenotic ileocecal valve tissues from patients with CD and in normal ileocecal valve tissues from HCs. We found that AIM2 expression in stenotic ileocecal valve tissues from patients with CD is significantly increased compared with that in normal ileocecal valve tissues from HCs (Figure 6A–B). In addition, we found significantly increased density of CD11b^+^ macrophages and elevated AIM2^+^ cell density in CD specimens compared with the HC group. Dual colocalization analysis revealed a marked increase in CD11b^+^ AIM2^+^ double-positive cells in patients with CD vs HCs (Figure 6C–F). Integrated with prior findings (Figure 1), these results indicate that pathogenic stimuli trigger macrophage AIM2 overexpression in humans, which subsequently drives excessive GSDMD-mediated pyroptosis, ultimately exacerbating intestinal fibrosis and stenosis in CD.Figure 6AIM2 expression is upregulated in stenotic ileocecal valve tissues from patients with CD. (A–B) IHC analysis of AIM2 levels in stenotic ileocecal valve tissues from patients with CD and in normal ileocecal valve tissues from HCs. (A) Representative IHC images. Scale bars: 200 μm. (B) Quantitative analysis by Image-Pro Plus software. IOD, integrated optical density. (C–F) Double immunofluorescence costaining of CD11b and AIM2 in stenotic ileocecal valve tissues from patients with CD and in normal ileocecal valve tissues from HCs. (C) Representative immunofluorescence images. Scale bars: 50 μm. (D–F) The number of CD11b^+^ and AIM2^+^ cells per 300 μm^2^. CD11b^+^ AIM2^+^ macrophages were counted under a microscope. (B, D–F) Data represent means ± SEM (n = 8–10 per group); ∗∗∗∗P < .0001 by Student’s t-test.
Discussion
This study elucidates a novel mechanism by which IMP alleviates intestinal fibrosis in CD through suppression of AIM2-mediated pyroptosis in macrophages. We provide multi-level evidence demonstrating that excessive GSDMD-dependent pyroptosis drives fibrotic progression in CD and establish AIM2 inflammasome inhibition as the primary therapeutic mechanism of IMP. Critically, clinical specimens from patients with CD with ileocecal stenosis revealed significant enrichment of CD11b^+^ GSDMD-NT^+^ macrophages (Figure 1), implicating pyroptotic hyperactivity in human disease pathogenesis. This observation was functionally validated in vivo, where genetic ablation of Gsdmd markedly attenuated intestinal fibrosis in a TNBS-induced chronic colitis model (Figure 3). The anti-fibrotic efficacy of IMP was consistently demonstrated through its ability to restore colon architecture, downregulate fibrotic markers (Collagen I, α-SMA), and suppress inflammatory cytokines (IL-1β, TNF-α) (Figure 4). Crucially, IMP administration significantly reduced AIM2 protein expression, Caspase-1 activation, and GSDMD cleavage in colonic tissues (Figure 4I–J), whereas in vitro studies confirmed direct inhibition of AIM2-mediated pyroptosis in macrophages (Figure 2).
The centrality of AIM2 as a key regulatory node is further underscored by spatial analyses. In TNBS-challenged mice, IMP treatment reduced F4/80^+^ AIM2^+^ macrophage infiltration (Figure 5), paralleling reduced pyroptosis and fibrosis. Importantly, human stenotic tissues exhibited elevated CD11b^+^ AIM2^+^ macrophages compared with HCs (Figure 6), establishing clinical relevance for this pathway. To explore the potential mechanism underlying AIM2 downregulation by IMP in macrophages, RNA sequencing of J744a.1 cells with or without IMP stimulation was performed to uncover differentially expressed genes (DEGs). There were 9 upregulated DEGs in the IMP group compared with the control group, and 13 downregulated DEGs (Figure 2H). Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis revealed the functional enrichment of protein processing in endoplasmic reticulum, a post-transcriptional modification (Figure 2I). Moreover, Gene Ontology (GO) analysis revealed the functional enrichment of protein deubiquitination (Figure 2J). Among the DEGs, the USP18 protein is a deubiquitinating enzyme (Figure 2H). Although these results suggest that IMP may regulate AIM2 protein stability through the deubiquitinating enzyme pathway, such as an USP18-dependent post-transcriptional manner, the precise molecular mechanism warrants further investigation.
Our results reshape understanding of macrophage-driven fibrogenesis. Conventionally, fibrosis is attributed to M2-polarized macrophages via TGF-β1 secretion.23 However, we demonstrate that pyroptosis-associated cytokine release (eg, IL-1β) directly activates fibroblasts to produce TGF-β (Figure 2E),24 creating a feed-forward loop independent of polarization states.25 This aligns with observations that pyroptotic macrophages release copious IL-1β and TNF-α,21 which are known fibroblast mitogens and inducers of endothelial-mesenchymal transition.26 Thus, dysregulated pyroptosis intensity, rather than macrophage polarization per se, emerges as a critical determinant of fibrotic outcomes.27 IMP’s ability to fine-tune this axis (reducing pathologic pyroptosis while preserving microbial defense) offers a strategic advantage over global immunosuppressants.
Several limitations merit consideration. First, the absence of macrophage-specific AIM2 knockout data limits mechanistic depth, though IMP’s efficacy across models supports target engagement.28 Second, although we established AIM2 as IMP’s functional target, its direct molecular binder (whether AIM2 itself or upstream regulators) remains undefined.29 Third, cell-type-specific contributions (eg, IECs or fibroblasts) to IMP’s anti-fibrotic effects require elucidation using conditional knockout models. Four, the translational potential of IMP enema necessitates further pharmacokinetic and safety studies. Nevertheless, our work provides foundational insights: (1) GSDMD-mediated pyroptosis is mechanistically linked to CD-associated intestinal fibrosis; (2) AIM2 inflammasome activation is a pathogenic trigger; and (3) IMP represents a promising therapeutic agent acting through AIM2 suppression.
7In conclusion, we identify hyperactivated AIM2-GSDMD pyroptosis as a key driver of intestinal fibrogenesis in CD. By demonstrating IMP’s efficacy in targeting this pathway—evidenced by reduced AIM2 expression, diminished pyroptosis, and attenuated fibrosis across human, murine, and cellular models—our study nominates AIM2 inhibition as a viable strategy for treating CD-associated intestinal fibrosis. Future efforts should focus on delineating the molecular interface of IMP-AIM2 interaction and advancing localized IMP delivery for clinical translation.
Materials and Methods
Antibodies, Reagents, and Resources
The antibodies, reagents, and resources used in this study are provided in Table 1.Table 1. Key Antibodies, Reagents, and ResourcesReagent or resourceSourceIdentifierAntibodies Rabbit Anti-AIM2 polyclonal antibodyAbsinCat.# abs125828 Phospho-Smad2/3 rabbit polyclonal antibodyAbsinCat.# abs130992 Rabbit anti-TNFα polyclonal antibodyAbsinCat.# abs131997 Anti-GSDMD antibodyAbcamCat.# ab209845 Rabbit anti-mouse IgGAbcamCat.# ab6728 CD11b rabbit mAbAbcamCat.# ab8878 GAPDH rabbit mAbAbclonalCat.# a19056 Cleaved caspase-1 rabbit mAbCell Signaling TechnologyCat.# 89332S IL-1β rabbit mAbCell Signaling TechnologyCat.# 31202 Smooth muscle actin polyclonal antibodyProteintechCat.# 14395 Collagen type I polyclonal antibodyProteintechCat.# 14695 FITC conjugated donkey anti-rabbit IgG (H+L)ServicebioCat.# GB22403 Cy3 conjugated goat anti-rabbit IgG (H+L)ServicebioCat.# GB21303 Anti-collagen I rabbit polyclonal antibodyServicebioCat.#GB11022 Anti-alpha smooth muscle actin rabbit polyclonal antibodyServicebioCat.#GB111364Chemicals and recombinant proteins RNase-free DTTAnsiangCat.# 5-750 FormalinAnsiangCat.# 5-171 Fetal bovine serumAusGenXCat.# No.FBS500-S PVDF membranesEMD MilliporeCat.#: C3117 RIPA bufferFUDECat.# FD008 HRP substrateEMD MilliporeCat.#: 18-160 2,4,6-trinitrobenzenesulfonic acid (TNBS)SigmaCat.# P2297 Maxpar fix and perm bufferFluidigmCat.# S00092 HE dye solutionServicebioCat.# G1005 Sirius Red staining solutionServicebioCat.#G1018 Hematoxylin staining solutionServicebioCat.# G1004 Eagle’s medium (DMEM)GIBCOCat.# 30030 DMEMGLPBIOCat.# 11320033 CollagenaseGLPBIOCat.# 17104019 transfection reagent RNAi-MateGenePharmaCat.# G04001 TrypsinHARVEYBIOCat.# 9002-07-7 Physiological salineNucleoTechCat.# NS23032-10 Protein-free rapid blocking bufferEpiZymeCat.# PS108P Ethanol anhydrousMerdaCat.# M155707-25L IsofluraneRWDCat.# R510-22 Bovine serum albuminServicebioCat.#GC305006 Citric acid antigen-retrieval solutionServicebioCat.#G1202 Ethylenediaminetetraacetic acid disodium salt dihydrateSigmaCat.#: E5134-500G DNase ISigma-AldrichCat.# AMPD1 ParaformaldehydeSigma-AldrichCat.# P6148 Phosphate buffered salineSolarbioCat.# P1010 Propidium iodideSolarbioCat.# P8080 Gentle cell dissociation reagentStemcell TechnologiesCat.# 7174 EasySep Releasable RapidSpheresStemcell TechnologiesCat.# 50201 ImperatorinMacklinCat. # No. I875280 Penicilin-streptomycin liquidSolarbioCat.# P1400 TSV405 TSnanofect V2TSINGKECat. # No. TSV405 Protein G immunoprecipitation magnetic beadsSelleckCat. # B23201 SDS-PAGE loading bufferYEASENCat. # 20315ES05 Poly(deoxyadenylic-thymidylic) acid sodium salt (poly(dA:dT))SigmaCat. # P0883 LipopolysaccharideMacklinCat. # L861706 CELLSAVINGNCM BiotechCat. # C40100 RNase-free waterTakaraCat.# No.9012 XyleneTCICat.# D0429 DAPIThermo Fisher ScientificCat.# 62248 AgaroseYeasenCat.# 10208ES60 5× HieffCanace PCR Master MixYeasenCat.# 10137ES08 Phosphatase inhibitor cocktailYeasenCat.# 20109ES20Experimental models: cell lines J774A.1FUHENGCat.# FH0329Experimental models: organisms/strains C57BL/6 miceThis paperN/A C57BL/7 mice (Gsdmd^-/-^)This paperN/AE.Z.N.A. soil DNA KitOmega Bio-tekCat.# M1768BCA kitBeyotimeCat.# P0009RNAiso PlusTakaraCat.# No. 9108IL-1β ELISA kitEIAABCat.# E0563mTGF-β ELISA kitEIAABCat.# E0124hRT-PCR kitTakaraCat.# RR014PAGE Gel quick preparation kitYeasenCat.# 20324ES62Mouse tissue lysis componentYeasenCat.# 19697ES50Hifair Ⅲ reverse transcriptaseYeasenCat.# 14601ES10Deposited data RNA-seq (J744A.1, Ctrl and IMP)This manuscriptPRJCA020332Software and algorithms ImageJNIHhttps://imagej.nih.gov/ij/ Prism software v9.0GraphPad Softwarehttps://www.graphpad.com/
Human Samples
Mucosal tissues of ileocecal valves were collected from patients with CD and HCs attending the Department of Gastroenterology of Nanfang Hospital. Disease diagnoses were assessed by a standard combination of clinical, endoscopic, and histological criteria. Mucosal tissue samples were collected from consenting individuals during routine endoscopy according to the protocols approved by the Ethics Committee of Nanfang Hospital of Southern Medical University. Demographic characteristics are shown in Table 2.Table 2. Demographic Characteristics of the Study PopulationCDControlsNo.1515Male, n (%)7 (46.6)8 (53.3)Age, years26.25 ± 8.75 (14–56)28.75 ± 9.25 (20–58)Disease duration, months30.6 ± 12.8 (2–60)_Age at diagnosis, years20.00 ± 6.5 (14–32)_NOTE. Data are presented as number (%) or mean ± standard deviation (range).CD, Crohn’s disease.
Mouse Models
WT and Gsdmd^-/-^mice (C57BL/6 strain) were bred and maintained in a pathogen-free facility, receiving standard chow and water ad libitum. The Gsdmd^-/-^ mice were kindly provided by Professor Feng Shao (National Institute of Biological Sciences, Beijing, China). The Gsdmd^-/-^ mice were conducted using cohoused WT littermates as controls. All animal-related experimental protocols were approval by the Institutional Animal Care and Use Committee of Southern Medical University (Approval NO. L2018231).
Our study examined male mice because male animals exhibited less variability in phenotype. A TNBS-induced colitis and fibrosis model was established according to the published procedures.30 Briefly, 8- to 10-week-old male Gsdmd^-/-^ mice and their WT littermates were pretreated with 100 μL TNBS presensitization solution (1% TNBS dissolved in 4:1acetone: olive oil solution), which was applied to shaved back skin on day −7. These mice were lightly anesthetized with isoflurane. Next, they were intrarectally injected with 100 μL 2.5% TNBS (controls were injected with 100 μL PBS) through a flexible catheter inserted 3.5 cm into the rectum on days 0, 7, 14, 21, 28, and 35. In the IMP-treated groups, WT mice were intrarectally injected with 100 μL 3 mM IMP 1 day before TNBS enema. During experiments, body weight was monitored daily. Colon tissues were embedded in paraffin and stained with hematoxylin and eosin (H&E), and histologic scores were assessed as in the published procedures.30 Sirius red staining was performed and calculated by ImageJ software to quantify the fibrosis degree of colon tissues.
Cell Culture and Transfection
J774A.1 cells were from Fuheng Biology Corporation and were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco) supplemented with 10% fetal bovine serum (FBS) in 5% CO_2_ at 37°C. These cells were transfected with 1 μg/μL LPS plus 2.5 ug/mL poly (dA: dT) for 8 hours to induce pyroptosis.
Western Blotting
Western blotting procedures were performed as previously described.31 The density of protein bands was analyzed and quantified by ImageJ software. The specific antibodies used in this study are provided in Table 1.
IHC Analysis
IHC analysis was conducted to evaluate the expression levels of AIM2 in paraffin-embedded tissues using specific anti-AIM2 antibodies. Protein expression levels were analyzed and scored by professional pathologists with Image-Pro Plus software.
Immunofluorescence Staining
The tissue sections were fixed in 4% neutral buffered formalin for 5 minutes and subjected to immunofluorescence staining using primary antibodies targeting CD11b, F480, GSDMD-NT, IL-1β, AIM2, Collagen Ⅰ, and α-SMA. Subsequently, the sections were incubated with a biotinylated secondary antibody against immunoglobulin (Ig) and streptavidin-fluorescein isothiocyanate (FITC). Nuclei were counterstained with 4',6-diamidino-2-phenylindole (DAPI). Ten randomly selected fields of view were captured from each slide using a fluorescent microscope. The fluorescence intensities of each field were quantified using ImageJ software. The specific antibodies used for immunofluorescence staining are listed in Table 1.
RNA Sequencing
RNA sequencing (RNA-seq) was employed to evaluate the mRNA expression levels in J744A.1 cells, with and without IMP treatment. For RNA extraction, cells were lysed in 1000 μL of Buffer RLT (with the addition of 10 μL β-ME per 1 mL of Buffer RLT prior to use) and thoroughly mixed by vortexing or pipetting. The selection of the sample quality control protocol was based on sample types and specific requirements. Reverse transcription was conducted to generate first-strand cDNA using SMART technology. Polymerase chain reaction (PCR) amplification was subsequently performed to preamplify the cDNAs. Qualified amplified cDNAs were then utilized for library construction using a transposon-based method.
LDH Release Assay
J774A.1 cells were seeded in 6-well plates and were stimulated with 50 μM IMP or control medium for 6 hours, and then transfected with LPS (1 μg/μL) plus poly (dA: dT) (2.5 ug/mL) for 8 hours to induce pyroptosis. The culture supernatants were collected, and the levels of LDH in the supernatants were quantified using an LDH assay kit (Solarbio).
PI Staining
PI staining technique was used to observe the morphological pattern of IMP improving pyroptosis. Properly treated J774A.1 cells were seeded into 6-well plates and then stained with 100 μL PI (P8080-10mg, Solarbio) at room temperature for 15 minutes in the dark and then washed twice with PBS. The results were visualized under a fluorescence microscope (BX63; Olympus) at 20× magnification. The excitation and emission wavelengths were 535 nm and 614 nm.
Enzyme-linked Immunosorbent Assay
J774A.1 macrophages were treated with or without 50 μM IMP for 6 hours following transfection with 1 μg/μL LPS plus 2.5 ug/mL poly (dA: dT) for 8 hours, and then their culture supernatants were collected for IL-1β assay. In addition, their culture supernatants were collected to stimulate the human colon fibroblast cell line CCD-18Co. After 24 hours, CCD-18Co cells were changed to a new medium and cultured for 12 hours, and subsequently, their culture supernatants were collected for TGF-β assay.
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 9 software. All variables are expressed as means ± standard error of the mean (SEM) or means ± standard deviation (SD), as noted in the figure legends. One-way analysis of variance (ANOVA) test was used for comparisons among 3 or more groups, and unpaired Student’s t-test for comparison between 2 groups. Statistical significance was defined as ∗P < .05, ∗∗P < .01, ∗∗∗P < .001, and ∗∗∗∗P < .0001.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Torres J.Mehandru S.Colombel J.F.Peyrin-Biroulet L.Crohn’s disease Lancet 3892017174117552791465510.1016/S 0140-6736(16)31711-1 · doi ↗ · pubmed ↗
- 2Roda G.Chien Ng S.Kotze P.G.Crohn’s disease Nat Rev Dis Primers 62020223224202810.1038/s 41572-020-0156-2 · doi ↗ · pubmed ↗
- 3D’Alessio S.Ungaro F.Noviello D.Revisiting fibrosis in inflammatory bowel disease: the gut thickens Nat Rev Gastroenterol Hepatol 1920221691843487668010.1038/s 41575-021-00543-0 · doi ↗ · pubmed ↗
- 4Lin X.Wang Y.Liu Z.Intestinal strictures in Crohn’s disease: a 2021 update Therap Adv Gastroenterol 1520221756284822110495110.1177/17562848221104951 PMC 921844135757383 · doi ↗ · pubmed ↗
- 5Latella G.Rogler G.Bamias G.Results of the 4th scientific workshop of the ECCO (I): pathophysiology of intestinal fibrosis in IBDJ Crohns Colitis 82014114711652473183810.1016/j.crohns.2014.03.008 · doi ↗ · pubmed ↗
- 6D’Haens G.Rieder F.Feagan B.G.International Organization for Inflammatory Bowel Disease Fibrosis Working Group Challenges in the pathophysiology, diagnosis, and management of intestinal fibrosis in inflammatory bowel disease Gastroenterology 162202226313125450210.1053/j.gastro.2019.05.072PMC 7680655 · doi ↗ · pubmed ↗
- 7Rieder F.Fiocchi C.Intestinal fibrosis in inflammatory bowel disease - current knowledge and future perspectives J Crohns Colitis 220082792902117222510.1016/j.crohns.2008.05.009 · doi ↗ · pubmed ↗
- 8Fiocchi C.Lund P.K.Themes in fibrosis and gastrointestinal inflammation Am J Physiol Gastrointest Liver Physiol 3002011 G 677G 6832141541110.1152/ajpgi.00104.2011 PMC 3094134 · doi ↗ · pubmed ↗