Presence and relevance of serum anti-C1P IgG antibodies in relapsing-remitting multiple sclerosis
Justyna Chojdak-Lukasiewicz, Anna Jakubiak-Augustyn, Zdzislaw M. Szulc, Jerzy Gubernator, Anna Pokryszko-Dragan, Slawomir Budrewicz, Maria Podbielska

TL;DR
This study found that certain antibodies against C1P are more common in people with relapsing-remitting multiple sclerosis, suggesting they could help diagnose or understand the disease.
Contribution
The study identifies specific anti-C1P IgG antibodies as potential biomarkers for relapsing-remitting multiple sclerosis.
Findings
Anti-C18:0-C1P and anti-C24:1-C1P IgG levels were significantly higher in RRMS patients compared to healthy subjects.
Anti-C24:1-C1P IgG levels were lower in RRMS patients during relapse compared to remission.
Anti-C1P IgG panels showed potential for differentiating between RRMS, OND, and healthy subjects.
Abstract
There is a growing interest in the role of sphingolipids in the background of multiple sclerosis (MS). The goal of this study was to evaluate the serum levels of antibodies against ceramide-1-phosphate (C1P) subclasses and their relationships with clinical status in MS. The study groups comprised 39 patients with relapsing-remitting MS (RRMS), 26 patients with other neurological diseases (OND) and 12 healthy subjects (HS). Anti-C1P IgG levels in serum were determined using ELISA test. Levels of anti-C18:0-C1P and anti-C24:1-C1P IgG were significantly increased (p = 0.0005; p < 0.0001, respectively) in RRMS compared to HS, while anti-C16:0-C1P and anti-C24:0-C1P IgG – significantly lower (p < 0.0001) in RRMS compared to OND. Validation by ROC and cluster analysis confirmed the ability of these anti-C1P IgG panels to discriminate between the study groups. In addition, level of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
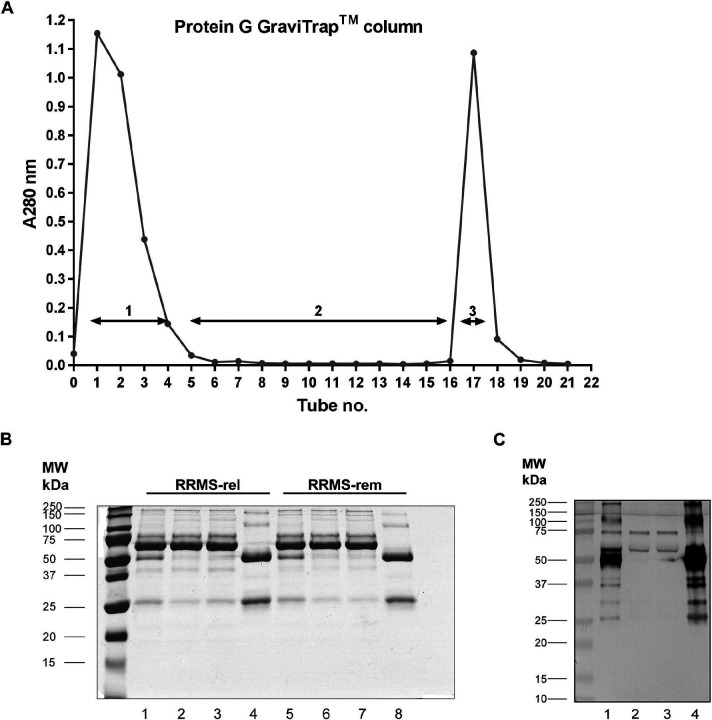 Figure 1
Figure 1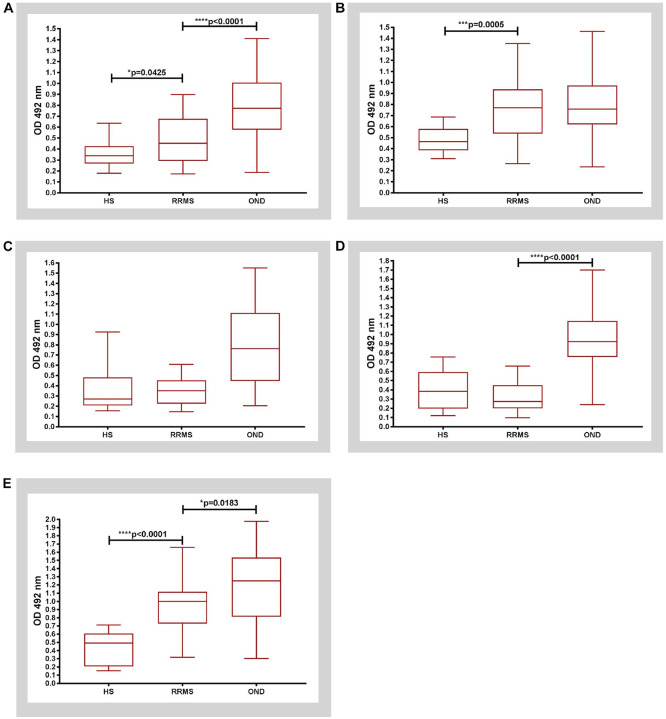 Figure 2
Figure 2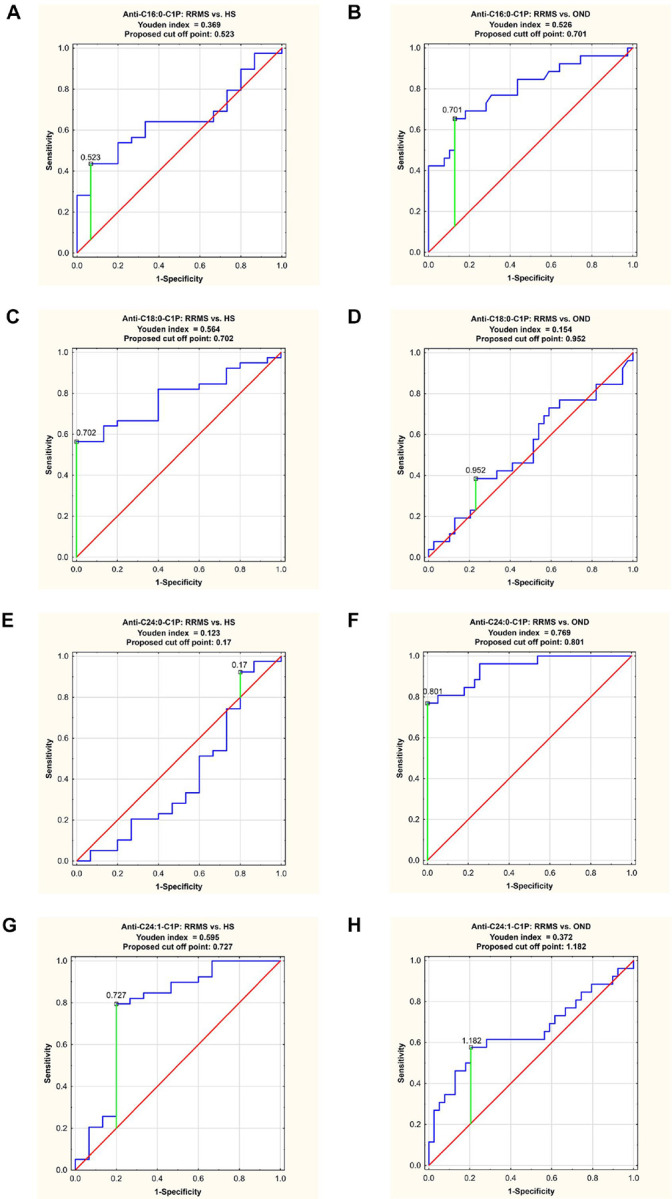 Figure 3
Figure 3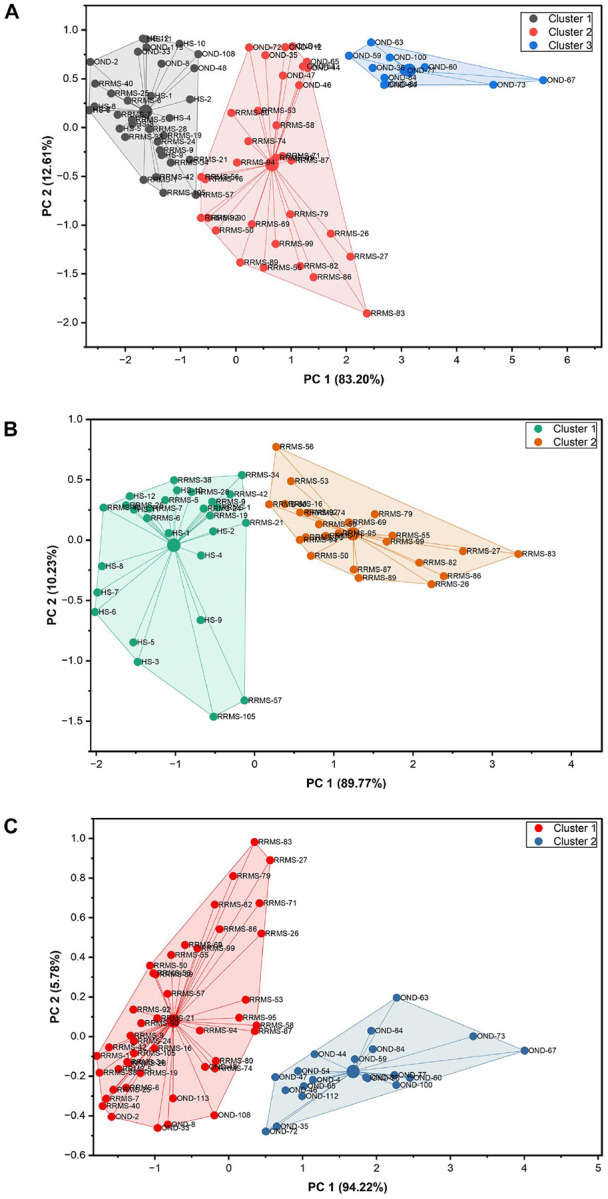 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSphingolipid Metabolism and Signaling · Spondyloarthritis Studies and Treatments · Peripheral Neuropathies and Disorders
Introduction
Multiple sclerosis (MS) is a chronic polyphasic disease of the central nervous system (CNS) which causes multifocal damage to the brain and the spinal cord and results in multiple aspects of disability. The complex background of MS involves two major components: autoimmune-mediated inflammatory injury to the myelin sheath and neurodegeneration with progressive axonal loss. These two processes develop in parallel but with different dynamics throughout the disease and their interplay determines MS-related damage to CNS, with emerging clinical picture of this disorder^1^. Despite a substantial progress in understanding MS pathophysiology, some of its aspects have not been yet elucidated, including key elements of initiating immune-mediated inflammatory cascade in CNS or factors modulating highly variable course of the disease^2^. Thus, reliable markers are being extensively sought, which would allow to identify and predict activity and progression of the disease^3^. Investigations have long been focused mainly upon protein molecules as indices of immune activity or neuronal/axonal damage. However, with the advent of metabolomic approach, lipids have also gained attention in this field^4–7^. Sphingolipids (SL) seem to be promising candidates for the role of such biomarkers, as the main components of CNS myelin sheath^8^, but also relevant players in cellular signaling and cross-talk between inflammation and neurodegeneration^9^. There has been a solid evidence of accumulation of bioactive SL mediators, specifically ceramide (Cer), in different types of MS lesions and microstructurally altered CNS tissues. Cer and its glycosylated derivatives have been also revealed in the cerebrospinal fluid (CSF), serum, plasma and white blood cells of MS patients^7,10–13^.
Previous research by the members of our team indicated a significant role of Cer derivatives in initiation and shaping of the autoimmune response, targeted at CNS antigens, a key element of MS background. The analysis of the cellular and exosomal profile of bioactive SL mediators in human oligodendroglioma (HOG) cells showed that exosomal Cer subspecies (C16:0-, C24:0- and C24:1-Cer) participate in the apoptosis process induced by pro-inflammatory Th1 cytokines^14^. Comparative assessment of the lipid profile in brain tissues collected during autopsy (demyelinating plaques in the specimens from MS patients vs. unaffected cerebral white matter in the specimens from the control group) allowed to establish specific SL markers that differentiated active and inactive chronic MS lesions^15^. Active lesions were characterized by a significant increase in the main subclasses of dihydroceramide (dhCer), while inactive ones – by decreased levels of dhCer, Cer and sphingomyelin (SM) subclasses and significantly elevated levels of hexosylceramides (HexCer) and ceramide-1-phosphates (C1P). These findings suggested a key element of ‘pathological switch’ in the metabolic pathways of Cer, driving alternately active or inactive phase of the disease.
Furthermore, analysis of anti-Cer immunoglobulin G (IgG) in serum and CSF revealed elevated levels of their particular subclasses (anti-C16:0-, C18:0-, C18:1-, C24:0- and C24:1–Cer) in MS group in comparison with healthy subjects (HS) and those with other CNS disorders^16^. In addition, differences in anti-Cer IgG panels were observed between subgroups with different clinical types of MS [i.a. relapsing-remitting (RRMS) vs. progressive]. For selected anti-Cer IgG subclasses, significant correlations were found between their level in CSF and serum, as well as Link index. The results indicated the relevant role of Cer as a target for autoreactive humoral response, an essential element of MS pathology.
Encouraged by the above findings, which supported the need for further research aimed at identification of SL putative biomarkers for MS activity and progression, we undertook the current study focused on particular Cer derivatives. Our main goal was to evaluate the serum level of IgG antibodies against selected C1P subclasses in MS subjects in comparison with the reference groups. We also aimed at investigating their relationships with clinical status.
Materials and Methods
Human specimens of serum
Participants were recruited among the patients hospitalized at the Department of Neurology, Wroclaw Medical University. The following 3 groups were included in this study: i) patients with RRMS (n = 39), diagnosed according to McDonald’s criteria^17^; ii) patients with other inflammatory disorders of the nervous system (I-OND, n = 13) that comprised chronic inflammatory demyelinative polyneuropathy (CIDP) or immune-mediated vasculitis, as well as iii) patients with non-inflammatory disorders of the nervous system (NI-OND, n = 13), including migraine, cerebrovascular disease and hydrocephalus.
Exclusion criteria in RRMS and I-OND groups comprised: recent (within previous 3 months) short-term treatment with corticosteroids, any kind of long-term immunomodulatory or immunosuppressive treatment during at least previous 6 months, current infection and coexisting systemic diseases, in particular inflammatory ones. In the RRMS group, clinical data, such as duration of disease and degree of disability in Expanded Disability Status Scale (EDSS), have been determined based on medical records. The patients were divided into two subgroups, according to their clinical status: those in relapse – exacerbation of disease (RRMS-rel, n = 23) or in remission – a period of stable condition (RRMS-rem, n = 16).
In addition, the blood samples were obtained from healthy subjects (HS, n = 12) at the Regional Center of Blood Donation and Blood Treatment in Wroclaw.
Participation in the study was anonymous and voluntary. All the subjects gave informed written consent to participate in this study. The project was approved by the Bioethics Committee of Wroclaw Medical University (No. KB 337/2024). The study was conducted in accordance with the Declaration of Helsinki and Good Clinical Practice guidelines.
Approximately 15 ml of peripheral venous blood was collected from each participant. After coagulation the samples were centrifuged at 2000xg at room temperature for 10 min, and afterwards stored at −80°C prior to IgG preparation.
Preparation of IgG antibodies from human sera
The IgG antibodies were isolated from human serum by affinity chromatography on Protein G GraviTrap^™^ columns (GE Healthcare). Prior to chromatographic separations, serum samples (0.5 ml) were centrifuged for 10 min at 10,000xg at 4°C, then diluted 1:1 (v/v) with 20 mM phosphate buffer pH 7.0 and filtered through a 0.45 μm Millex^®^-HV filter. Columns containing 1 ml of packed gel were equilibrated with phosphate buffer and prepared in the described above way sera were loaded. Proteins not bound by protein G were eluted from the columns with phosphate buffer. Next, the column was washed with 12 column volumes of phosphate buffer and the wash fraction was collected. Elution of IgG, bound to the ligand, was performed with 0.1M Gly/HCl pH 2.7 and subsequently neutralized with 1M Tris/HCl pH 9.0. During the fractionation the amount of proteins in all fractions was monitored by measuring the absorbance at 280 nm on 96 well UV half-plate area plates in an Enspire reader. The fractions containing IgG only were concentrated on an Amicon^®^ Ultra100K by ultracentrifugation at 4500xg at 4°C to a final volume of 250 μl. During this process, IgG samples were desalted and albumin residues were removed. The protein concentration of column fractions was measured using bicinchoninic acid (BCA) protein assay kit (Pierce) with bovine serum albumin (BSA) as a standard. Purified IgG antibodies were preserved with 0.02% NaN_3_, aliquoted and stored at −80°C prior to further analysis.
SDS-PAGE and Western blotting analysis
The electrophoretic separation was performed in a BIO-RAD^®^ Tetra Cell Systems apparatus as described earlier^16^. Briefly, proteins (2 μg) were analyzed on 12% polyacrylamide gel in reducing conditions and either stained with Coomassie Brilliant Blue (CBB) or transferred into 0.45 μm Immobilon P. The separated proteins were detected with, produced in donkey, anti-human IgG (H + L), conjugated with alkaline phosphatase (AP), antibody (1:7500) (Sigma). The reaction was visualized with AP substrates, according to standard procedure.
Synthesis of C1P derivatives
C1P derivatives were synthesized in the Synthesis Unit of the Lipidomics Shared Resources at the Medical University of South Carolina, Charleston, SC, USA, as previously described^18^. Their purity was checked by the high performance liquid chromatography- tandem mass spectrometry (HPLC-MS/MS).
ELISA assay
ELISA plates (Immulon 1B, Thermo Fisher Scientific) were coated with selected C1P solutions in absolute ethanol (10 μg/ml; 0.1 ml /well). The plates were incubated overnight at room temperature to facilitate complete solvent evaporation. Next, the plates were washed three times with phosphate buffered saline (PBS; 0.2 ml/well). Nonspecific binding sites were blocked by adding 100 μl of 1% (w/v) BSA in PBS and incubating the plates for 1 hour at 37°C. After washing 5 times with PBS purified IgG (0.1 mg/ml of protein, 0.1 ml /well) from serum of MS patients and individuals from reference groups were added and the plates were incubated overnight at 4°C. In the next step the plates were washed again for 5 times with PBS and then incubated with horseradish peroxidase (HRP) conjugated anti-human IgG (GE Healthcare, 1:2000, 0.1 ml/well). The enzymatic activity was developed by adding peroxidase substrate (Sigma tablets). The colorimetric reaction was stopped by adding of 100 μl of 3N H_2_SO_4_. and the optical density (OD) was read at 492 nm in an Enspire reader.
Statistical analysis
Ordinary one-way Anova with post-hoc Tukey test was used to estimate differences in respect to age of examined subjects belonging to different groups.
The U Mann-Whitney test was used to compare levels of serum IgG anti-C1P antibodies between patients with RRMS and reference groups (HS, OND) as well as between RRMS-rel and RRMS-rem subgroups. The diagnostic significance of immunoreactivities of these IgG antibodies with appropriate C1P antigens was analyzed using received operating characteristic (ROC) curves with clinical meaning considered as zero (values from 0–0.5); limited (values from 0.5–0.7); moderate (values from 0.7–0.9) and high (> 0.9). The Youden index method was used for the determination of cut off points.
The IgG samples, for which the area under the curve (AUC) values were established as moderate or high, were subjected to a cluster analysis, a multivariate statistical method. Methods belonging to two different categories were used. A hierarchical Ward’s method was applied to determine the number of clusters. A non-hierarchical method, namely k-means clustering (k-means function of base R) from the selected antiC1P IgG dataset was used for further analysis.
The Spearman’s rank correlation was used to check the associations between measured levels of anti-C1P antibodies and clinical parameters such as disease duration, EDSS and Link index.
The ROC analysis was performed using Statistica 13.3 software (StatSoft Inc., Tuls, OK, USA), cluster analysis was executed by OriginPro 2025b (OriginLab Corporation, Northampton, MA, USA). For the remaining statistical analysis GraphPad Prism 7.01 (GraphPad Software Inc., San Diego, CA, USA) was applied.
Results
Analysis of the study cohort
Basic demographic data, including age and sex structure of the patients included in the study are presented in the Table 1. One-way Anova followed by Tukey’s multiple comparisons test indicated no significant age differences between the study groups. In RRMS group, duration of the disease ranged from 1 to 108 months (mean 22 months) and EDSS score – from 1 to 4 (median 1.5) points. All the RRMS patients (n = 39) had magnetic resonance imaging (MRI) of the brain performed, with typical demyelinative lesions. In 32 patients the results of CSF testing were available, with confirmed presence of IgG oligoclonal bands and calculated Link index (increased value ≥ 0.7 in 20 cases).
Analysis of Protein G-isolated and purified from human serum IgG antibodies
The IgG antibodies present in human serum were isolated by affinity chromatography on Protein G Gravi Trap columns. A representative preparation of IgG antibodies from 0.5 ml of serum is shown on Fig. 1. In a typical elution profile obtained during the isolation of IgG from RRMS-rel patient two peaks were visible (Fig. 1A). The first one − Fr. 1 − represented the proteins unbound by protein G. The second one, Fr. 3, corresponded to four isotypes of IgG (IgG_1_, IgG_2_, IgG_3_ and IgG_4_), bound by protein G. The obtained fractions were subjected to qualitative and quantitative analysis. The CBB staining (Fig. 1B) obtained as a result of electrophoretic separation of samples, collected during the isolation of serum IgG from both RRMS-rel and RRMS-rem, indicated that in both cases the products obtained as a Fr. 3 were IgG antibodies (lanes 4 and 8). This is evidenced by two characteristic bands with a molecular mass of 50 and 25 kDa, corresponding to the heavy and light chain of immunoglobulin G, as appeared under these conditions. In the remaining lanes (lanes 1–3 and 5–7), distinct bands with a molecular mass of about 65 kDa were observed, originating from other proteins found in the serum. Presumably, these bands indicate the presence of albumin, which constitutes up to 65% of all serum proteins, and perhaps the heavy chain of immunoglobulin M, constituting about 5 to 10% of serum immunoglobulins. Quantitation of proteins in the various fraction demonstrated high and comparable total protein amounts in Fr. 1 and residual protein amounts in Fr. 2. The preparation resulted in the recovery of 5.6 ± 0.9 mg of purified IgG (Fr. 3), a predominant class of antibody in serum, which corresponded to 14.3% ± 2.4 of all serum proteins (Supplemental Table 1S). The absence of IgG in Fr. 1 and 2 (Fig. 1C; lanes 2 and 3, respectively) and the presence of the purified IgG antibodies was confirmed by Western blotting (Fig. 1C, lane 4). This analysis indicated the high efficiency of the IgG purification method used.
Altered levels of serum anti-C1P IgG in RRMS patients
The Protein G-purified IgG antibodies from serum of MS patients (Fr. 3) were evaluated for the presence of selected C1P subclasses binding activity, using an ELISA (Fig. 2 and Supplemental Fig. 1S).
The binding activity of serum IgG to C16:0-C1P was significantly higher in RRMS patients (median value: 0.4535) in comparison to HS group (median value: 0.34) and significantly lower in relation to OND group (median value: 0.7725), with significance of p = 0.0425 and p < 0.0001, respectively (Fig. 2A).
The binding activity of serum IgG to C18:0-C1P in RRMS patients (median value: 0.7715) was significantly increased in comparison to HS group only (median value: 0.4635) with significance of p = 0.0005 (Fig. 2B). In turn, there were no significant differences in the binding activity of serum IgG to C18:1-C1P in RRMS individuals in relation to both reference groups (Fig. 2C).
In contrast, RRMS patients manifested lower level of anti-C24:0-C1P serum IgG (median value: 0.2725) in comparison to OND group (median value: 0.924) with significance of p < 0.0001 (Fig. 2D).
The binding activity of serum IgG to C24:1-C1P represented the pattern similar to C16:0-C1P. Interestingly, serum IgG derived from RRMS patients exhibited the highest reactivity for C24:1-C1P among all C1P subclasses examined (median value: 1.001) in comparison to HS group (median value: 0.4915, p < 0.0001), while the level of anti-C24:1-C1P IgG was significantly decreased in RRMS in comparison to OND group (median value: 1.25; p = 0.0183) (Fig. 2E). The presented above results, obtained by ELISA test, are summarized in Table 2.
Next, we reanalyzed the data considering the division of RRMS patients into subgroups in relapse and in remission (RRMS- rel and RRMS -rem) and OND patients – into subgroups with inflammatory or other etiology of disease (I-OND, NI-OND) (Supplemental Fig. 1S). The Mann Whitney test showed significantly higher levels of serum IgG derived from both RRMS subgroups against most C1P subclasses (except of C24:0-C1P) compared to HS, particularly significant for IgG anti-C18:0-C1P and C24:1-C1P (p = 0.0024 and p < 0.0001, Supplemental Fig. 1S, panel B & E, respectively). The level of IgG anti-C24:1-C1P in RRMS-rel subgroup was significantly lower than in RRMS-rem (p = 0.0448), as well as lower in both RRMS subgroups in relation to NI-OND only (p < 0.0001 and p = 0.0004, Supplemental Fig. 1S, panel E, respectively). The level of IgG anti-C24:0-C1P in both RRMS subgroups was significantly lower than in I-OND and NI-OND (p < 0.0001, Supplemental Fig. 1S, panel D). The above results in details are included in Supplemental Table 1S.
ROC analysis
In order to validate ELISA results, we performed ROC analysis for all serum anti-C1P IgG levels in RRMS patients, as presented in Table 2. The results of ROC curve analysis for anti-C1P IgG data sets with the AUC value higher than 0.700 are presented in Fig. 3. The cut-off points, determined by the Youden index method, are shown in Table 3.
Anti-C16:0-C1P IgG level indicated a moderate value in differentiating RRMS group from OND group (Fig. 3B), with sensitivity 65.4% and specificity of 97.2%, respectively (proposed cut off point: 0.701, AUC = 0.798). Anti-C18:0-C1P IgG level showed moderate discriminating value between RRMS and HS groups (Fig. 3C), with sensitivity and specificity of 56.4% and 100%, respectively (proposed cut off point: 0.702, AUC = 0.781). The level of anti-C24:0-C1P IgG had a high discriminating value between RRMS group and OND group, with sensitivity of 76.9% and specificity of 100%, respectively (proposed cut off point: 0.801, AUC = 0.942) (Fig. 3F). The level of anti-C24:1-C1P IgG exhibited a moderate value in differentiating between RRMS group and HS group, with sensitivity of 79.5% and specificity of 80% (proposed cut off point: 0.727, AUC = 0.769) (Fig. 3G).
Cluster analysis
Based on the results of ROC analysis, serum anti-C1P IgG levels with AUC equal or greater than 0.769 were selected to perform cluster analysis. The number of expected clusters was pre-determined by using the hierarchical Ward’s method (Supplemental Fig. 2S). Relative level of anti-C16:0-, anti-C18:0-, anti-C24:0- and anti-C24:1-C1P serum IgG was subsequently used to perform k-means clustering to categorize the samples examined into 3 clusters (Fig. 4A). Cluster 1 comprised 34 subjects: all healthy ones; 16 patients with RRMS (including 9 ones from RRMS-rel subgroup) and 6 individuals with OND (containing 5 from I-OND subgroup). Cluster 2 included 23 RRMS patients (14 from RRMS-rel, 9 from RRMS-rem subgroup) and 9 OND ones (5 from I-OND and 4 from NI-OND subgroup). Cluster 3 consisted of OND patients only, with the majority belonging to NI-OND subgroup (8 out of 11) (Supplemental Fig. 3S).
Significant differences in anti-C1P IgG panels between the study groups (RRMS vs. HS: anti-C18:0- and anti-C24:1-C1P IgG; RRMS vs. OND: anti-C16:0- and anti-C24:0-C1P IgG) (Fig. 2), with discriminatory ability of these IgG subsets, confirmed by ROC analysis (Fig. 3, Table 3), together with similar patterns of relative levels of anti-C16:0- / anti-C24:0-C1P, as well as anti-C18:0- /anti-C24:1-C1P IgG (Supplemental Fig. 4S), supported the notion that anti-C1P IgG panels might differentiate pathological from physiological condition but also reflect distinct biological processes (i. e. neuroinflammation vs. neurodegeneration) with different degrees of sensitivity. To check this hypothesis, a k-means analysis was subsequently performed to divide the appropriate IgG samples into 2 clusters. Relative levels of anti-C18:0- and anti-C24:1C1P serum IgG were subjected to k-means clustering to differentiate RRMS from HS group (Fig. 4B). Cluster 1 comprised 28 subjects (12 HS and 16 RRMS, almost equally distributed between RRMS-rel and RRMS-rem subgroups) whereas cluster 2 was represented by 23 RRMS patients (mainly RRMS-rel subgroup) (Supplemental Fig. 5S). In turn, relative levels of anti-C16:0- and anti-C24:0-C1P serum IgG were subjected to k-means clustering to differentiate RRMS from OND group (Fig. 4C). Cluster 1 comprised 45 subjects: all RRMS patients (n = 39) and 6 OND ones (mainly from I-OND subgroup), whereas cluster 2 was composed of 20 OND subjects, mainly from NI-OND subgroup (Supplemental Fig. 6S).
Correlation analysis between selected serum anti-C1P IgG derived from RRMS patients and clinical parameters
Next, we examined whether there are any correlations between anti-C1P IgG titers (as reflected by OD at 492 nm) in RRMS group and MS-related clinical parameters. The levels of anti-C16:0-C1P, anti-C18:0-C1P, anti-C18:1-C1P, anti-C24:0-C1P and anti-C24:1-C1P IgG in RRMS did not correlate with disease duration (Supplemental Fig. S7, panels A, D, G, J and M) or EDSS score (panels B, E, H, K and N), accordingly. No associations were found between the levels of these antibodies and Link index, either (Supplementary Fig. S7, panels C, F, I, L and O, respectively). The summary of this analysis is displayed in Table 4.
Discussion
Cer products have been suggested to be involved in the main mechanisms underlying MS pathology, including those - recently highlighted - driven by B cells. Activation of B cells together with cytokine stimulation promote tissue penetration and destabilization of membrane lipids in CNS, resulting in damage to myelin sheath. SL (in particular Cer derivatives) are essential for maintenance of lipid membranes integrity, so changes in their metabolism and proportion correspond with dynamic processes of demyelination and repair and their pathophysiological consequences^19^. Although in the aforementioned studies there is convincing evidence for Cer abnormalities in MS, it relies mainly on preclinical models and their translation into human clinical context, with clear identification of Cer related mechanisms of action and potentially relevant MS biomarkers, still remains a challenge^20 4–7^. Based on results of our previous studies^14–16^ and scarce literature data^10,12,13,21,22^, here we focused on particular Cer derivative – C1P, aiming to investigate its putative role as a target and/or mediator of autoimmune response in MS.
C1P, a major metabolite of Cer, can be formed through a stereospecific reaction of phosphorylation, due to direct action of Cer kinase (CerK), while the reverse reaction (conversion of C1P to Cer) is mediated by C1P phosphatase. An appropriate balance between these two molecules is required to ensure cells and tissues homeostasis through regulation of relevant metabolic or signaling pathways. Thus, coordinated action of CerK and C1P phosphatases may have crucial importance for maintaining this balance. Its disturbances with a shift towards accumulation either Cer or C1P may result in metabolic dysregulation, supposedly involved in the background of autoimmune diseases^23^, neurodegenerative disorders^24^, cardiovascular illnesses^25^ or cancer^26^.
It has been demonstrated that C1P exerts pro-inflammatory properties; however, the underlying mechanisms have not been yet fully elucidated. One of the major function of C1P is its capability to activate cytosolic phospholipase PLA_2_ (cPLA_2_), which mediates release of arachidonic acid^27^ - the substrate used for production of eicosanoids, such as prostaglandin E_2_ (PGE_2_). This is aligned with the fact that MS patients exhibit higher PGE_2_ expression/activity in CSF^28^, lymphocytes^29^ as well as demyelinative lesions in brain tissue^30^. A sharp increase of this pro-inflammatory mediator was observed in active stage of MS^31^, and experimental studies with the use of cyclooxygenase (COX) inhibitors, which oppose C1P action by preventing PGE_2_ synthesis, suggested their beneficial effect as potential options in MS therapy^32^.
Other pro-inflammatory activities affected by C1P include: activation of degranulation in mast cells^33^, stimulation of phagocytosis in neutrophils^34^, as well as stimulation of macrophage migration^35^. C1P also increases transport of P-glycoprotein, which regulates the permeability of the blood brain barrier via COX-2/PGE_2_ signaling^36^.
On the other hand, it has been shown that C1P might also act as an anti-inflammatory agent, under specific conditions. Anti-apoptotic effects of C1P, promoting cell survival, are related with its ability to block activity of serine palmitoyltransferase^37^, acid sphingomyelinase^38^ and TNF-α^39^. Furthermore, it promotes macrophage chemoattractant protein-1 (MCP-1) release in different types of cells^35^. Taken together, C1P seems to act on its own rights, executing regulatory effects and modulating cell functions depending on circumstances.
In one of the previous reports by our team members, an increase in C1P subspecies (mainly C18:1-C1P and C24:1-C1P) was demonstrated in inactive MS lesions in comparison with specimens representing other CNS disorders or normal brain tissue. On the contrary, active MS plaques were characterized by a significant decrease of C1P subspecies^15^. The studied autopsy samples from MS subjects represented advanced stage of disease, associated with chronic inflammation and presumably extensive neurodegenerative component. In the present study we focused on earlier phase of the disease (i. e. RRMS), when active immune-mediated inflammation is a predominating mechanism of CNS injury - therefore antibodies as markers of autoimmune humoral response were chosen as the subject of investigation. The main findings presented here included elevated levels of nearly all analyzed anti-C1P IgG in serum of RRMS patients in comparison to HS, with the highest significance for anti-C24:1-C1P and antiC18:0-C1P (Fig. 2E and B). This striking consistency in the results from both studies, despite differences in MS stages and studied material (brain tissues vs. serum), indeed suggests the relevant role of C1P in MS pathology. It can be hypothesized that each stage of MS is associated with the concerted action of Cer derivatives, with their changing interactions in this pathological cross-talk.
It should be mentioned that the level of IgG anti-C24:1-C1P was significantly lower in both RRMS subgroups in relation to NI-OND only (Supplemental Fig. 1S, panel E), whereas the level of IgG anti-C24:0-C1P in both RRMS subgroups was significantly lower than in both OND subgroups (Supplemental Fig. 1S, panel D). Overall, these results suggest that increased humoral response against long-chain C1P subspecies is particularly specific for MS and it does not just reflect overall neuronal injury or dysregulated immune-mediated inflammation in CNS. Importantly, the reliability of the above findings was supported by eliminated impact of immunosuppressive/immunomodulatory drugs, as the patients were included in the study prior to such treatment or after appropriate wash-out period.
There is an evidence that fatty acid chain length of Cer can determine biophysical properties of this SL mediator^40^. In physiological conditions, medium chain Cer (C16 and C18) are known to be enriched in cerebral gray matter, while long chain ones (C24) - in white matter and myelin sheath^41^. Considering distribution of Cer derivatives in the brain, presence of the medium chain ones would reflect neuronal injury^42^, while the long chain ones – oligodendrocytes degradation^14^. The long chain Cer (C24) are also known to be the major Cer subspecies present in serum/plasma whereas C1P constitute only 0.15% of all SL components^43^. However, it should be taken into account that the pattern of anti-Cer IgG in body fluids, revealed by our previous study^16^ reflected metabolic perturbations of Cer, supposed to occur due to processes involved in MS background. The anti-Cer IgG alterations in serum (unlike in the CSF) were observed only for Cer containing medium chains (C16:0-, C18:0- and C18:1-Cer) with no change for the long chain ones^16^. Current findings revealed altered patterns of IgG against C1P in serum of RRMS patients. Here we found that anti-C16:0-C1P (Fig. 2A), anti-C18:0-C1P (Fig. 2B) and anti-C24:1-C1P (Fig. 2E) serum IgG levels were increased in RRMS patients in comparison with HS group. In turn, levels of IgG against C16:0-C1P (Fig. 2A), C24:0-C1P (Fig. 2D) and C24:1-C1P (Fig. 2E) were decreased in RRMS in relation to OND (including distinguished subgroup of inflammatory ones). In addition, the results of ROC analysis indicated that the determination of anti-C18:0-C1P and anti-C24:1-C1P serum IgG levels might be useful for discriminating RRMS from HS group (Fig. 3, panel C and G), whereas anti-C16:0-C1P and anti-C24:0-C1P serum IgG levels – for discriminating RRMS from OND group (Fig. 3, panel B and F). Furthermore, the usefulness of the above anti-C1P IgG subsets in distinguishing pathological from physiological conditions, as well as disorders with different etiology, was confirmed by means of multivariate statistical method of cluster analysis (Supplemental Fig. 5S and 6S). Despite somehow subjective selection of variables for clustering and the number of clusters (justified by the results of initial analysis), the findings from this multivariate technique support the concept that panels/sets of anti-C1P IgG may be of greater discriminating value than single immunological parameters (Fig. 4). Such a panel including anti-C16:0-C1P, anti-C18:0-C1P, anti-C24:0-C1P and anti-C24:1-C1P IgG might be considered for further investigation towards potential clinical implications - the use of anti-C1P IgG as biomarkers in differential diagnosis at MS onset, especially in atypical cases.
As a result of more precise analysis within RRMS group, the level of anti-C24:1-C1P IgG was found to be significantly lower in those with relapse than those in remission (Supplemental Fig. 1S, panel E). These findings may indicate that alterations of Cer metabolism (revealed by presence of corresponding IgG) are associated with changing phases of disease activity.
Few attempts have been made to use anti-Cer antibodies as biomarkers of activity in other disorders, including those of the nervous system. In leprosy, presence of anti-Cer antibodies was found to be related to extent of nerve sheath damage and particular bacterial strains as pathogenic factors^44^. In the rare autoimmune condition, encephalomyeloradiculoneuropathy (EMRN), accumulation of species-specific Cer (mainly long chain ones) was demonstrated in CSF and presence of anti-neutral glycolipids antibodies – in CSF and serum. Profile of these antibodies allowed to differentiate patients with ERMN from the reference group with Parkinson disease, and their titers decreased after effective immunomodulatory treatment. Dysregulation of glycolipids metabolism and its impact upon invariant natural killer T cell development, as well as abnormal complement activation, were suggested to play a relevant role in EMRN pathophysiology^41,45^. Furthermore, in non-small-cell lung cancer (NSCLC) anti-Cer antibodies were supposed to be produced by the tumor and its microenvironment, and to neutralize pro-apoptotic effect of Cer. Significantly elevated levels of anti-Cer antibodies were revealed in serum and bronchial wash fluid samples from NSCLC group (in comparison with HS) and they showed a tendency (though not significant) for prolonged overall survival^46^.
Evaluation of activity in MS has been so far based on clinical assessment and radiological indices (new and/or contrast-enhanced lesions in MRI). However, there is an ongoing search for specific and reliable body fluid biomarkers of disease activity^8^. Their clinical applications would include i.a. differentiating relapse from pseudorelapse (temporary aggravation of symptoms) or defining active phase in progressive stage of the disease, and in long term follow-up – monitoring of MS course and response to treatment. In view of our previous and current findings, Cer derivatives and antibodies against them seem indeed promising candidates for the role of such biomarkers.
Although no relationships were found in our study group between immunological measures and MS-related clinical data (Supplemental Fig. S7), potential clinical utility of these findings should not be discredited. The group was characterized by a relatively short MS duration and mild disability, and marked individual differences in disease course in MS populations must be taken into account. Perhaps in a larger group of patients and in prospective observation these relationships would appear more relevant.
Indeed, a small sample size and focusing only on RRMS should be considered as limitations of our study, as its results may not accurately reflect abnormalities in SL metabolic pathways in different types and stages of the disease. Another limitation is associated with the fact that anti-C1P IgG presence was determined only in serum and not in CSF. Nevertheless, in view of long-term course of MS and need for regular follow-up, serum/plasma biomarkers are much desired due to their better availability. Based on experiences with other body fluid biomarkers, their serum level may adequately reflect their production in CNS^8^. There is also some evidence that peripheral blood may contribute to formation of immune complexes due to selective transport and the blood-brain barrier (BBB) leakage^47^.
To our best knowledge, this study for the first time focuses on the presence and meaning of anti-C1P antibodies in serum of MS patients and provides evidence of their potential relevance. The results shed some light on the perturbation on SL metabolism (especially significance of balance between Cer and C1P) in the background of neurological diseases, specifically MS. Encouraged by these findings, further investigation is planned with the use of IgG against particular C1P subclasses, analyzed parallel in blood and CSF, in larger groups of MS patients and with prospective observation of immunological and clinical parameters. Hopefully, these studies will lead to a better understanding of the role of this intriguing phosphosphingolipid in pathophysiology of MS, with emerging clinical implications.
Conclusions
The serum levels of IgG anti-C1P subspecies (especially long-chain ones) differed significantly between the patients with RRMS and healthy subjects, as well as the patients with other neurological diseases. Differences were also found between subgroups of RRMS patients with relapse or with remission. These findings indicate the relevant role of C1P as a target and/or mediator of autoimmune response in MS. Potential value of anti-C1P antibodies as biomarkers in differential diagnosis of MS and monitoring disease activity warrants further investigations.
Supplementary Material
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
AntiC1PAbsinMSSupplementalMaterialSciRepformatted.docx
Supplementary information
Supplemental material for this article can be found in the online version of this article.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dobson R. & Giovannoni G. Multiple sclerosis - a review. Eur. J. Neurol. 26, 27–40. 10.1111/ene.13819 (2019).30300457 · doi ↗ · pubmed ↗
- 2Lassmann H. Pathogenic Mechanisms Associated With Different Clinical Courses of Multiple Sclerosis. Front. Immunol. 9, 3116. 10.3389/fimmu.2018.03116 (2018).30687321 PMC 6335289 · doi ↗ · pubmed ↗
- 3Ciubotaru A. Neurodegenerative Biomarkers in Multiple Sclerosis: At the Interface Between Research and Clinical Practice. Diagnostics (Basel). 15 10.3390/diagnostics 15091178 (2025). · doi ↗
- 4Pousinis P. Recent Advances in Metabolomics and Lipidomics Studies in Human and Animal Models of Multiple Sclerosis. Metabolites 14, 545 (2024).39452926 10.3390/metabo 14100545 PMC 11509141 · doi ↗ · pubmed ↗
- 5Smusz J. Metabolomics in Multiple Sclerosis: Advances, Challenges, and Clinical Perspectives-A Systematic Review. Int. J. Mol. Sci. 26 10.3390/ijms 26189207 (2025). · doi ↗
- 6Lattau S. S. J. Plasma Lipidomic Profiling Using Mass Spectrometry for Multiple Sclerosis Diagnosis and Disease Activity Stratification (Lipid MS). Int. J. Mol. Sci. 25 10.3390/ijms 25052483 (2024). · doi ↗
- 7Amatruda M. Retrospective unbiased plasma lipidomic of progressive multiple sclerosis patients-identifies lipids discriminating those with faster clinical deterioration. Sci. Rep. 10, 15644. 10.1038/s 41598-020-72654-8 (2020).32973249 PMC 7515876 · doi ↗ · pubmed ↗
- 8Chertcoff A., Schneider R., Azevedo C. J., Sicotte N. & Oh J. Recent Advances in Diagnostic, Prognostic, and Disease-Monitoring Biomarkers in Multiple Sclerosis. Neurol. Clin. 42, 15–38. 10.1016/j.ncl.2023.06.008 (2024).37980112 · doi ↗ · pubmed ↗