Polyhydroxyalkanoate synthesis by Sinorhizobium meliloti drives a host-specific collapse in symbiosis with Medicago sativa
Barney Geddes, Garrett Levin, Chinh Luu, Natalie Visich, Scott Hoselton, Anna Lipzen, Shuang Zhao, Liang Li, George diCenzo, Turlough Finan

TL;DR
A gene region in Sinorhizobium meliloti that produces polyhydroxyalkanoates disrupts its ability to fix nitrogen with Medicago sativa, revealing how carbon metabolism affects symbiotic effectiveness.
Contribution
The study identifies a genetic mechanism in rhizobia that causes host-specific symbiotic failure through altered carbon metabolism.
Findings
A gene region from pSymA in Sinorhizobium meliloti HM006 is involved in polyhydroxyalkanoate production in nitrogen-fixing bacteroids.
Transferring this gene region to an effective symbiont strain causes complete loss of nitrogen fixation with Medicago sativa.
The mechanism involves a dehydrogenase (GhbD) diverting succinate semialdehyde to gamma-hydroxybutyrate, causing symbiotic collapse.
Abstract
Naturally occurring root-nodule bacteria (rhizobia) vary substantially in their effectiveness at promoting growth of different plant hosts via symbiotic nitrogen fixation. These variations in rhizobial partner quality have important implications for the productivity of nitrogen-fixing symbioses in natural and agricultural ecosystems, yet we have a limited understanding of the genetic basis for this variation. In a case of host-specific reduction in symbiotic effectiveness (N2-fixation) with Medicago sativa, we identified the causative genetic elements from the pSymA replicon of Sinorhizobum meliloti HM006 and show them to be involved in polyhydroxyalkanoate (PHA) production in nitrogen-fixing bacteroids. Transfer of this gene region to a strain that forms an effective symbiosis with Medicago sativa resulted in a complete loss of symbiotic N2-fixation. We showed the mechanism for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
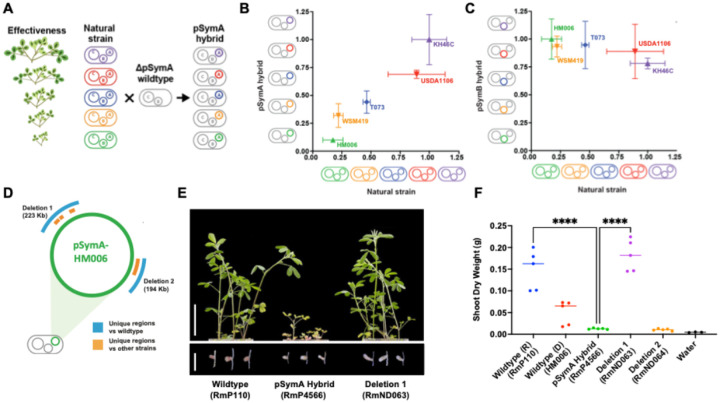 Figure 1
Figure 1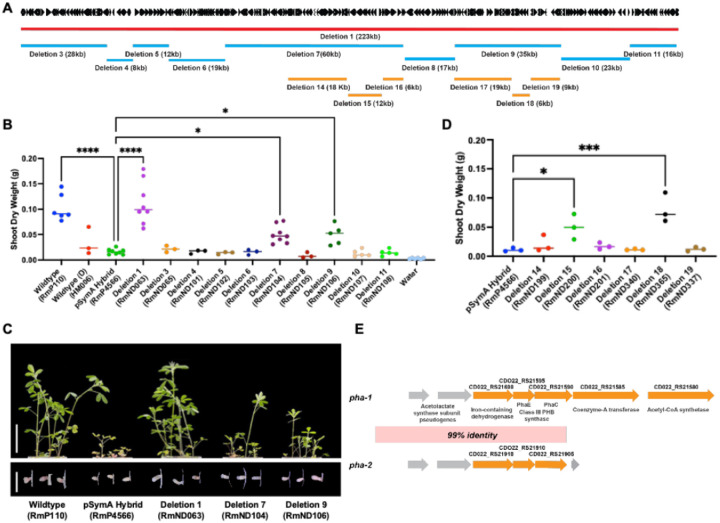 Figure 2
Figure 2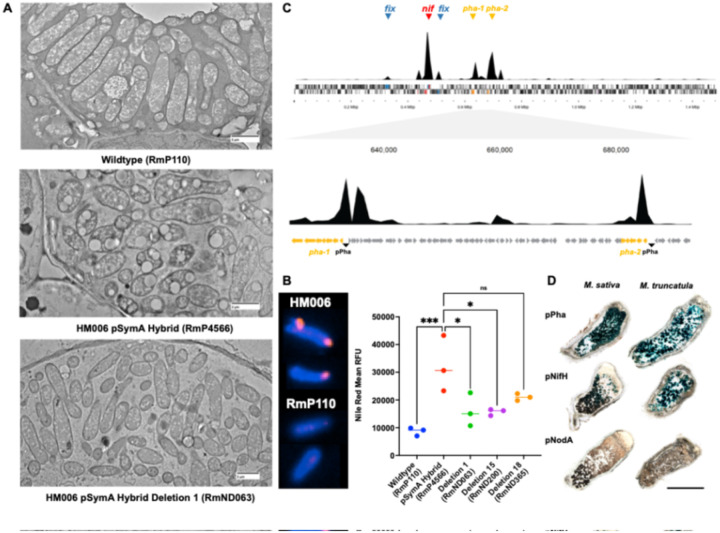 Figure 3
Figure 3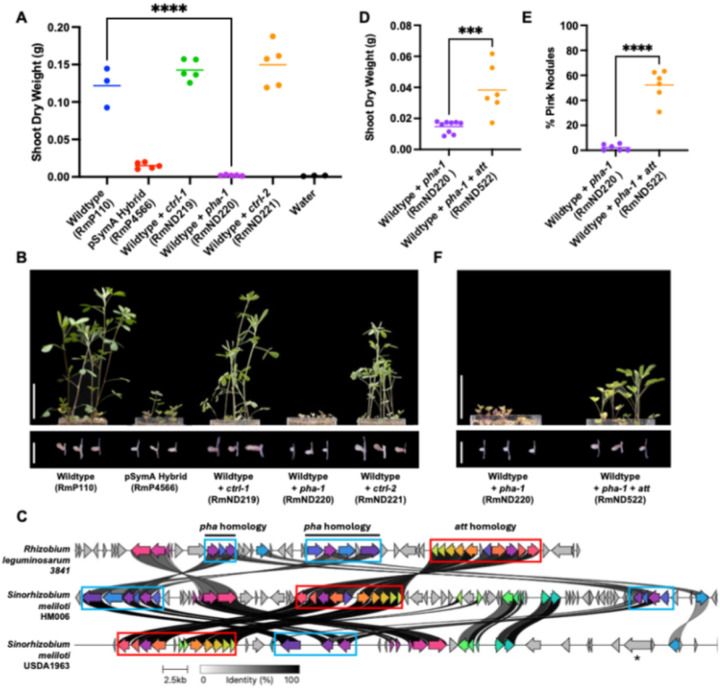 Figure 4
Figure 4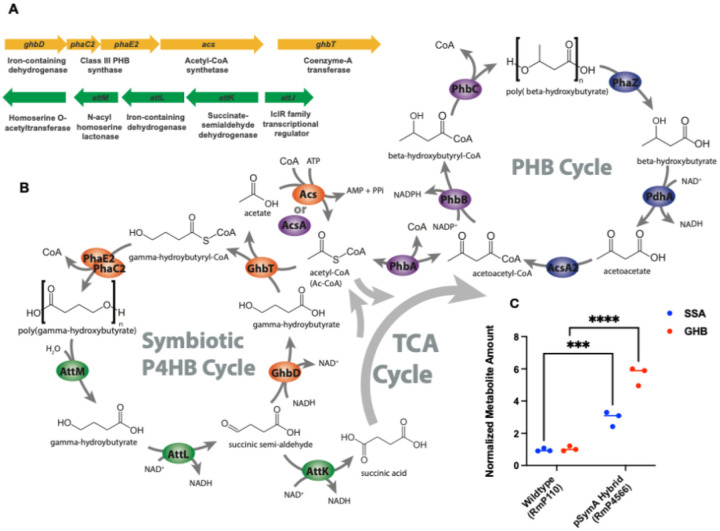 Figure 5
Figure 5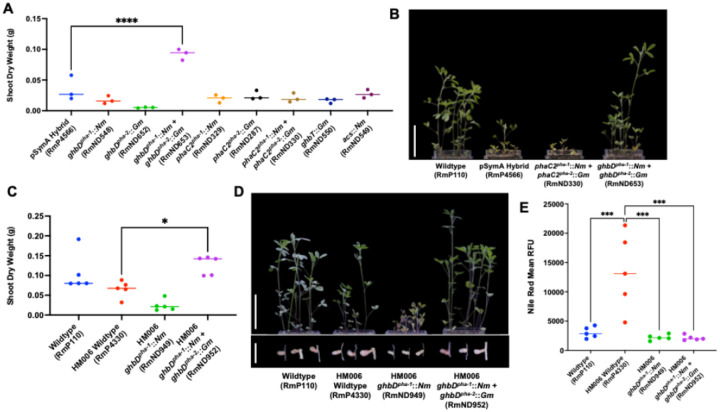 Figure 6
Figure 6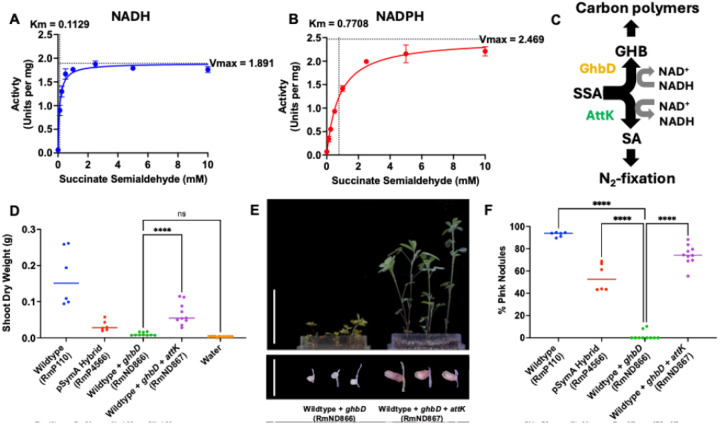 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegume Nitrogen Fixing Symbiosis · biodegradable polymer synthesis and properties · Plant-Microbe Interactions and Immunity
Introduction
As the primary sustainable source of nitrogen nutrition in agricultural and natural ecosystems, the nitrogen-fixing root nodule symbiosis between rhizobia and legumes is a central player in global biogeochemical cycling and sustainable agricultural productivity. Variation in the symbiotic capacity of rhizobia is widespread within symbionts that are able to nodulate and fix nitrogen with a given host legume (1–3). Locally adapted native strains that are less effective at fixing nitrogen for their hosts limit the contribution of the symbiosis to agricultural systems by outcompeting elite inoculant strains for nodulation, dubbed “the rhizobium competition problem” (4–6). Though mechanisms remain elusive, hosts partially overcome this variation by selectively rewarding or punishing (sanctioning) rhizobia based on their productivity (7–10). Uncovering the genetics responsible for variation in microbial services to their hosts and elucidating what forces (e.g., “cheating”) maintain such widespread variation in symbioses despite host sanctioning mechanisms have emerged as key questions in microbial ecology (3). Recent studies point towards much of this variation being associated with mobile modular units such as plasmids and integrative conjugative elements (ICE) that are common features of rhizobium genomes (1, 11, 12).
In the tripartite genome of the model rhizobium Sinorhizobium meliloti, the “symbiotic megaplasmid” pSymA contains many essential genes for symbiosis with its Medicago hosts (nod, nif and fix). These gene sets, which encode the ability to produce the “Nod Factor” that initiates nodulation of the host (the nod genes) and to fix nitrogen in root nodules (the nif and fix genes) (13), account for ~ 5% of its size, yet are sufficient for functional symbiosis in the absence of the remaining ~ 95% of pSymA (14). The chromid pSymB is more ancestral than pSymA and has been implicated in playing a primary role in saprophytic competence (15), while also carrying the dicarboxylate transport genes (dct) essential for fueling nitrogen fixation in the nodule and exopolysaccharide synthesis (exo) genes important for plant invasion (16, 17). Despite decades of genetic study, the vast majority of genes on these replicons remain undefined with respect to their contributions to symbiosis. In part, this is due to the challenging nature of performing genetic screens for more subtle phenotypes (e.g., partner quality or competition for nodule occupancy) than the nodule-minus phenotypes of nod mutants or nitrogen-fixation-minus phenotypes of nif and fix mutants (18, 19), or of genes associated with symbiotic incompatibility (20, 21).
The rapid increase in genome sequencing of rhizobium isolates in recent decades has revealed that rhizobia, such as S. meliloti, contain large, open pangenomes (11, 22), further demonstrating the unexplored genetics that could contribute to variation in symbiosis outcomes. This wealth of genomic data, coupled with next-generation sequencing approaches to screen rhizobium symbiosis traits en masse (23, 24), enable the prediction of candidate genes that may impact the mutualism using quantitative genetics such as genome-wide association studies (GWAS) (12, 25–27). However, thus far, few candidates identified by these studies have been empirically validated for their role in symbiosis.
Here, we used strains wherein extrachromosomal replicons have been cured (15, 28) as surrogate hosts to identify genetic determinants that define differences observed in the vast diversity of effectiveness of functional symbioses between Sinorhizobium strains and their Medicago hosts. First, we investigated the co-transfer of partner quality with extrachromosomal replicons from five natural strains. The pSymA replicons were found to have the greatest impact on symbiotic performance and in the most extreme case, the transfer of pSymA from strain HM006 resulted in a substantial reduction of symbiotic performance with Medicago sativa. Next, through a series of refined deletions and gain-of-function experiments, the locus responsible for limiting symbiotic nitrogen fixation was identified and shown to be dominant. Genes within the locus were found to be involved in carbon polymer synthesis during symbiosis, and their impact on symbiotic N_2_-fixation revealed an unanticipated aspect to carbon metabolism in root nodules and its impact on partner quality.
Results
Co-transfer of host-specific symbiotic effectiveness phenotypes with the Sinorhizobium replicons.
We hypothesized that most genes contributing to variance in partner quality would be present on the extrachromosomal replicons in sinorhizobia and reasoned we could identify key genes involved in partner quality by transferring the extrachromosomal replicons (~ 40% of genome) to a surrogate S. meliloti background in which pSymA, pSymB, or both pSymA and pSymB were removed (14, 15, 29, 30). For replicon transfer, we used four genome-sequenced S. meliloti strains: KH46, HM006, USDA1021, and T073 and one Sinorhizobium medicae strain, WSM419 (31, 32). These genome-sequenced strains showed significant variation in partner quality (shoot-dry weight accumulation under nitrogen-limiting conditions) with Medicago sativa and Medicago truncatula (33) (Supplemental Figure S1).
The pSymA or pSymB replicons from the five strains were transferred independently or together to derivatives of the long-studied wildtype chromosomal background SU47 (Rm1021-derived background strain RmP110, or Rm5000) (33, 34) (Fig. 1A). The symbiotic productivity of the resulting hybrid strains with M. sativa revealed a clear positive correlation between the abilities of the natural strains to promote plant growth, and the relative abilities of hybrid strains containing their respective pSymA replicons (Fig. 1B, Supplemental Figures S2A and S3A). In contrast, no significant difference in plant growth promotion was observed for pSymB transfer (Fig. 1C, Supplemental Figures S2A and S3A). With M. truncatula as a host, increased growth relative to the wildtype was also observed by transfer of pSymAs from more effective strains (2- to 3-fold increase in SDW) and was enhanced with co-transfer of pSymB (~ 6-fold increase in SDW) (Supplemental Figures S2B and S3B).
A genetic determinant on the S. meliloti HM006 pSymA leads to a collapse of symbiotic efficiency with M. sativa.
As a platform to investigate genetics that underly variation in pSymA partner quality, we chose the pSymA that conferred the most dramatic phenotype; a collapse in the productivity of the symbiosis with M. sativa conferred by the pSymA from S. meliloti HM006 when transferred to the RmP110 background, which forms an effective symbiosis with M. sativa when bearing its native pSymA. We first compared the pSymA contents of HM006 and RmP110. While known symbiosis gene sets in the pSymAs were nearly identical (Supplemental Fig. 4), two large regions from HM006 pSymA were found to be absent from the RmP110 pSymA (Fig. 1D, Supplemental Figure S5). Deletion of these regions from the HM006 pSymA-hybrid strain revealed that the loss of 223 kb in one region (Deletion 1, D1) completely restored symbiotic effectiveness with M. sativa, whereas the deletion of the second region (Deletion 2, D2) had no effect (Figs. 1E and 1F). Neither deletion significantly impacted symbiosis with M. truncatula (Supplemental Figure S6). We concluded that dominant genetic determinant(s) within the D1 region of the HM006 pSymA led to the collapse in effective symbiosis with M. sativa.
Additive inhibition of symbiotic effectiveness by a duplicated gene cluster on HM006 pSymA.
To identify genes within the 223 kb region removed in D1 responsible for reduced symbiotic N_2_-fixation with M. sativa, we generated nine sub-deletions and screened them for their symbiotic productivity with M. sativa (Figs. 2A and 2B). While none of the deletions fully restored symbiotic productivity, two sub-deletions, D7 and D9, each resulted in a ~ 50% recovery in shoot dry weight production (Figs. 2A, 2B, and 2C). Further sub-deletions within the 60 kb region removed in D7 (D14-D16) and the 35 kb region removed in D9 (D17-D19) localized the genes responsible for the partial recovery of M. sativa shoot dry weight production to two gene sets deleted in D15 and D18 (Figs. 2A and 2D). Both genetic loci, which we name pha-1 (removed in D15, locus tags CDO22_RS21580-CDO22_RS21610) and pha-2 (removed in D18, locus tags CDO22_RS21900-CDO22_RS21925), show an operon-like configuration with a 99.9% identity to one-another over a majority of their length (Fig. 2E). Both clusters included genes annotated as encoding an iron-containing alcohol dehydrogenase (TIGR02638 E-value 1.70e-60) and two subunits of a Class III polyhydroxyalkanoic acid synthase: PhaE (TIGR01834 E-value 1.33e-08) and PhaC (TIGR01836 E-value 2.94e-113/3.25e-85) in an operon-like configuration. Homology between the loci extends 2783 bp upstream of these gene sets and contains two annotated pseudogenes along with, presumably, the promoter. A putative operon in pha-1 extends beyond the duplicated region and included genes predicted to encode a Coenzyme A transferase (TIGR02428 E-value 8.25e-11) and an acetyl-CoA synthetase (TIGR02188 E-value 0).
Symbiotic carbon polymer synthesis leads to a host-specific inhibition of productive symbiosis with M. sativa.
Based on their annotation, we hypothesized that the gene sets in pha-1 and pha-2 may be involved in the production of a polyhydroxyalkanoate (PHA) carbon polymer such as polyhydroxybutyrate (PHB) during symbiosis. PHB is visualized by transmission electron microscopy (TEM) as large granules within bacteroids of rhizobia that synthesize PHB during symbiosis (35). TEM of M. sativa and M. truncatula nodules revealed the presence of large granules in the strains bearing pSymA-HM006. These granules were absent from bacteroids of the wild type or the D1 strain lacking both pha-1 and pha-2 (Fig. 3A, Supplemental Figure S7). While the small inefficient M. sativa nodules restricted further characterization, we also measured PHA production in effective M. truncatula nodules using flow cytometry, using PHA-bound Nile Red for quantitation (36) (Fig. 3B). A significant increase in Nile Red fluorescence of bacteroids was observed in wildtype HM006 and the HM006-pSymA hybrid strains relative to wildtype RmP110 (Fig. 3B). The increased fluorescence was significantly reduced in D1, which lacked both pha-1 and pha-2, as well as D15 which lacked the expanded gene set of pha-1 (Fig. 3B).
To examine the expression of the PHA synthesis genes during symbiosis, we analyzed the transcriptome of the HM006 pSymA hybrid strain in M. sativa and M. truncatula root nodules. RNAseq showed a high level of transcription across the regions that include the PHA synthesis genes in both hosts (Fig. 3C, Supplemental Figure S8). Several genes within them were among the top 50 most highly expressed genes from HM006 pSymA during symbiosis (Supplemental Table S10). We further observed robust pha cluster gene expression in nitrogen-fixing bacteroids of nodules using gusA promoter fusion constructs (Fig. 3D, Supplemental Figure S9). Altogether, these data indicate that the production of carbon polymers during symbiosis by bacteroids containing the newly discovered PHA genes drives a host-specific symbiotic incompatibility between S. meliloti and M. sativa.
Expression of the pha genes alone leads to a complete symbiotic collapse in a naïve background that is relieved by local catabolism genes.
To independently verify that the genes present in the pha loci are the sole components of the HM006 pSymA replicon responsible for the reduced symbiotic N_2_-fixation in M. sativa nodules, we applied a gain-of-function approach. We integrated the putative symbiotic PHA synthesis operon from pha-1 (Figs. 2A and 2E) into the genome of RmP110 with its native pSymA. Strikingly, RmP110 with pha-1 alone resulted in a more complete collapse of symbiosis than that associated with pSymA-HM006 (Fig. 4A). Shoot dry weight production was similar to uninoculated control plants (Fig. 4A), with few pink nodules (Fig. 4E) and barely detectable acetylene reduction activity (Supplemental Figure S10). TEM analysis showed that nodules were sparsely populated with bacteroids which showed signs of degradation by the host plant (37) (Supplemental Figure S11).
To rationalize the discordance between the partially reduced symbiotic efficiency conferred by the complete HM006 pSymA, and the near non-functional symbiosis rendered by the pha gene cluster alone, we hypothesized that other genetic determinants on HM006 pSymA alleviate the inhibition of symbiosis caused by the pha cluster. In search of such determinants, we identified other strains bearing phaC homologues by BLAST (Supplemental Table S11) and compared the content of the local genetic region surrounding phaC (Fig. 4C). These included the well-studied pea symbiont Rhizobium johnstonii (formerly Rhizobium leguminosarum) Rlv3841 as well as S. meliloti USDA1963. We found that a set of carbon metabolism genes previously characterized in A. tumefaciens C58 as involved in N-acylhomoserine lactone catabolism (attKLM) were consistently collocated with the pha cluster in other strains (38) (Fig. 4C), and we considered they could be responsible for alleviating the inhibition of symbiosis caused by the pha cluster alone when they were transferred with HM006 pSymA. We cloned the catabolism gene set and introduced them to the wild type that contained the pha gene cluster. Introduction of the catabolism genes was sufficient to restore functional symbiosis (Fix + phenotype) to the wildtype background characterized by increased shoot dry weight and pink nodule number (Figs. 4D, 4E, and 4F). Consistent with these data, Deletion 8 of HM006 pSymA which lacked these genes was more symbiotically impaired than other deletions (Fig. 2B).
Evidence for a novel symbiotic poly(gamma)hydroxybutyrate cycle.
We considered that the recovery of symbiosis mediated by the catabolism genes may be the result of a PHA cycle, wherein the pha synthesis genes produce a polymer, whereas the att catabolism genes are able to break it down. Based on the annotation of the function of homologues to the att genes in A. tumefaciens (Fig. 5A), we predicted a novel pathway for polymer synthesis and catabolism based on the intermediate gamma-hydroxybutyrate (Fig. 5B). The proposed pathway places the synthesis genes in a rational scheme for polymer production based on their annotations; oxidation of succinate semialdehyde to gamma-hydroxybutyrate by the iron-containing dehydrogenase, the addition of an acetyl-CoA group by the CoA transferase generating GHB-CoA (with acetyl-CoA substrate generated by acetyl-CoA synthetase), and polymerization of GHB-CoA monomers via the PhaE/PhaC PHB synthase. The breakdown of the P4HB product of the proposed synthesis pathway then fits with the characterized activities of homologues to AttL and AttK, wherein they break down GHB to succinate, fueling the TCA cycle and bacteroid nitrogen-fixation. In this scheme, the lactonase AttM could conceivably resolve monomers from a GHB polymer with a similar hydration reaction to the cleaving of the lactone ring in gamma-hydroxybutyrolactone (38). To provide evidence for the proposed pathway, we performed metabolomics of root nodules formed by rhizobia bearing or lacking these pathways, looking for the presence and abundance of pathway intermediates. Consistent with the pathway scheme above, we found significant increase of both the predicted pathway intermediates, SSA and GHB (Fig. 5C, Supplemental Figure S12).
Conversion of succinate semialdehyde to gamma-hydroxybutyrate by an iron-containing dehydrogenase in bacteroids is the mechanism for symbiotic collapse in M. sativa.
To gain further insight into the role of individual genes in the pathway, we constructed knockout mutations within genes from each step of the predicted pathway (ghbD, phaC2, ghbT, and acs), with double knockouts created for the duplicated genes ghbD and phaC2. Strikingly, only a double knockout within the iron-containing dehydrogenase ghbD (the predicted first step of the synthesis pathway) was able to recover symbiosis (Figs. 6A and 6B). These data suggested that the first step of the predicted pathway, which we predicted to direct carbon from SSA away from succinate in a reductant consuming reaction, rather than polymer synthesis per se, was responsible for symbiotic collapse. To ensure these data were not an artifact of working in the HM006 pSymA hybrid background, we also constructed double-knockouts of the iron-containing dehydrogenase in HM006 and showed a similar recovery of symbiosis with M. sativa (Figs. 6C and 6D), coincident with a reduction in carbon polymer synthesis observed by flow cytometry (Fig. 6E, Supplemental Figure S13).
Since GhbD, the iron-containing dehydrogenase, appeared the critical element of symbiotic collapse, we wished to verify its hypothesized activity in the predicted P4HB synthesis pathway (Fig. 5B). We purified the iron-containing dehydrogenase and demonstrated its activity with SSA as a substrate (Supplemental Figures S14 and S15). Purified GhbD showed a Km of 0.1129 or 0.7708 and a Vmax of 1.891 or 2.469 in an NADH/NADPH linked spectrophotometric assay when incubated with SSA and NADH or NADPH, respectively, indicating a higher affinity for NADH as a cofactor (Figs. 7A and 7B). The loss-of-function data combined with the verified activity of GhbD suggested that the primary mechanism for symbiotic collapse is the biochemical conversion of SSA to GHB in bacteroids rather than polymer synthesis, since phaC double mutants retained symbiotic collapse phenotype (Figs. 6A and 6B). To further support this presumption, we expressed the dehydrogenase in the naïve wildtype background, without the downstream portions of the operon predicted to be involved in GHB polymerization (Figs. 7D to 7F). We found this construct was still sufficient for the collapse of M. sativa symbiosis previously observed by transferring the whole operon (Fig. 4). Based on the confirmed biochemical activity of GhbD (Figs. 7A and 7B), we further predicted we could rescue the symbiotic phenotype caused by ghbD expression by overexpressing attK from the catabolism gene set which in Agrobacterium is involved in the conversion of SSA into succinate in a reductant generating reaction (38) (Fig. 7C). Such an activity would compete with the conversion of SSA to GHB in a reductant-consuming reaction by GhbD (Fig. 7C). Indeed, we found that including the attK gene downstream of ghbD in the gain-of-function construct significantly restored symbiosis between S. meliloti and M. sativa (Figs. 7D to 7F).
Discussion
Replicon transplantation experiments led to the finding that the synthesis of GHB-based hydrocarbon polymers is linked to the collapse of symbiotic effectiveness in M. sativa nodules leading to a reduction in partner quality. Several differences are notable between the carbon polymer production genes discovered here (Fig. 4H) and the well-characterized polyhydroxybutyrate synthesis genes (phbABC). First, annotations of genes in the operon (e.g., the inclusion of an iron-dependent dehydrogenase) are inconsistent with those in the well-characterized PHB synthesis pathway in rhizobia (40). Second, the high level of expression of these genes during symbiosis contrasts with the typically described pattern of PHB synthesis in S. meliloti, where PHB is produced in the free-living state and during infection thread invasion but not during symbiotic nitrogen fixation (36, 41). The inclusions in bacteroids observed in electron micrographs did not show the typical bright granules due to a lack of staining by lead citrate as is often observed for PHB (35). Finally, the observation of GHB accumulation in M. sativa nodules occupied by rhizobia containing the new pha synthesis genes (Fig. 5C), and the verification of GhbD as a dehydrogenase that reduces SSA to GHB (Fig. 7) further highlight the unique nature of this pathway. Altogether, these data support its existence as a novel symbiotic PHA synthesis pathway in rhizobia. We further supply evidence that the att catabolism genes impact the symbiotic phenotype associated with symbiotic PHA synthesis and propose that the pha and att clusters act together in a symbiotic P4HB cycle (Fig. 5B). A likely source of SSA, the substrate for the first step in the proposed synthesis pathway, is gamma-aminobutyrate (GABA) catabolism, which is provided to bacteroids from plant cells and catabolised to SSA via highly redundant transamination reactions (42–44).
It has been suggested that the production of reduced carbon storage polymers such as PHB may negatively impact the symbiosis by diverting resources away from nitrogen fixation (7). If the M. sativa phenotypes we document in this study was the result of diversion of resources from symbiosis via PHA synthesis, they would be by far the most dramatic observation of this phenomenon to date. To investigate the plausibility of this, we utilized an available host/symbiont metabolic model (VINE (45)) to investigate the degree to which the reduction in M. sativa partner quality could be explained by resource consumption (either reductant or carbon) by symbiotic PHA synthesis (Supplemental Fig. 16). While the modelling was consistent with resource consumption negatively impacting nitrogen fixation, the magnitude of reduction in shoot dry weight accumulation predicted by the model was significantly less than observed in M. sativa experimentally. We thus conclude that the observed symbiotic collapse cannot be explain simply due to diversion of reductant or carbon away from nitrogen fixation.
Instead, we hypothesize that the accumulation of GHB intermediates during PHA polymer synthesis in M. sativa nodules is toxic to Sinorhizobium bacteroids, resulting in a loss of nitrogen fixation capacity. This is supported by multiple observations. First, only mutations in the pha loci that abolish ghbD function enable the recovery of symbiosis in M. sativa (Fig. 6A). Similarly, expressing GhbD alone (converting SSA to GHB) in a naïve background resulted in a substantial reduction in symbiosis in M. truncatula, which was partially rescued by co-expressing AttL which diverts SSA to succinate (Supplemental Figure S18). Both of these observations suggest that synthesis of the pathway intermediates (rather than the polymer itself) was sufficient to collapse the symbiosis. Lastly, even though large PHA granules accumulate in bacteroids within both M. sativa and M. truncatula nodules, a substantial increase in GHB and SSA intermediates was only observed in M. sativa nodules (Supplemental Figure S17), consistent with our observation of a strong symbiotic phenotype only in this host.
The production of reduced carbon polymers such as PHB have been implicated in rhizobium “cheating”, in which the rhizobium benefits (storage of carbon for use once released from the nodule) at the expense of the host (reduced rate of nitrogen fixation) (7). However, it is not clear whether carbon polymer synthesis is a cheating behavior. Although PHB mutants showed increased nitrogen fixation in some rhizobia (46), limited effects were observed in others (35, 41, 46, 47). In fact, PHB production and other lipogenesis represent important reductant stores during symbiotic nitrogen fixation and therefore may facilitate rather than limit nitrogen-fixation capacity (48). In this study, removing the pha gene sets from HM006 pSymA did not significantly impact partner quality with M. truncatula (Supplemental Figure S6), even though the bacteroids within M. truncatula nodules did accumulate large PHA granules (Figs. 3B and 3D, Supplemental Figures S7, S8 and S9), suggesting that the polymer synthesis was not at the expense of the host. While P4HB synthesis in bacteriods came at a large cost to M. sativa, the proposed toxicity of the intermediates to the bacteriods and the complete collapse of the symbiosis likely means that P4HB synthesis hurt, rather than benefited, S. meliloti. Collectively, these results suggest that at least on the tested plants, P4HB synthesis is unlikely to represent a cheating behavior.
Presumably, the pha clusters confer some benefit to the rhizobia and/or the host to maintain them in rhizobium populations despite the potential for limiting partner quality on some hosts. PHA clusters have been theorized to benefit the symbiosis by providing a source of carbon to fuel SNF during carbon-limited scenarios (49), and the cycle we propose here would be particularly well suited to this purpose by engaging with central metabolism through dicarboxylates (unlike the canonical PHB catabolism pathway (50)), which must allow SNF in the nodule to drive ammonium excretion to the plant (51). PHAs accumulated in nodules could also benefit rhizobia outside of the nodule where they often encounter carbon-limited soil environments (36, 51). However, as bacteriods are terminally differentiated in Medicago nodules and thus cannot regrow in the soil, it may be that PHAs accumulated in bacteriods do not benefit free-living rhizobia. Alternatively, selection for maintenance of the pha clusters may be independent of the symbiosis, while their expression in bacteriods is non-beneficial and the result of inappropriate activation following detection of plant-produced signals.
Though the benefits conferred by the pha clusters to the microsymbiont and/or host remain to be elucidated, we expect that coevolution of these clusters in rhizobia with specific hosts can tune their activity to limit their negative impact on symbiosis, for example, through ensuring rapid flux through toxic pathway intermediates. Indeed, HM006 was isolated from M. truncatula nodules grown with soil from its native range (33, 52). When HM006 is present in M. truncatula, although the genes are highly expressed, we observes different gene expression patterns across the region relative to HM006 bacteroids in M. sativa nodules (Supplemental Figure S8), which may reflect the ability to fine tune the pathways through adaptation of transcriptional control. In addition, other genetic elements in the loci may participate in limiting the impact to specific hosts (Fig. 4C).
Partner quality differences in nature are widespread in many if not all rhizobium/legume systems (53–55). The replicon transplantation experiments described in this report revealed a correlation of partner quality phenotypes between M. sativa and the pSymA replicons of the tested strains. Thus pSymA (and perhaps the mobile symbiotic element in many rhizobia) likely represents the primary reservoir for accessary genes influencing partner quality in Sinorhizobium, although accessory genes on pSymB or additional plasmids can also influence symbiotic outcomes (21, 56). Given the much higher degree of genetic variation on extrachromosomal replicons relative to the chromosome (11, 22, 32, 57, 58), it is unsurprising these elements contain important determinants of the partner quality differences observed across different strains. Certainly, many mechanisms exist for the diversity in partner quality phenotypes observed between rhizobia and their different hosts, distributed through a reshuffling of genetic modules that engender metabolic flexibility but also can render individual partnerships inefficient (12, 59). This study suggests that along with diversity in symbiotic and immune signaling (60–63), metabolic incompatibility may be a key theme governing the diversity of these interactions we observe in nature.
Materials and Methods
Genetic Manipulations
Bacterial strains and plasmids used in this work are listed in Supplemental Table S1. Media used for routine growth and selection of microbes included LB for Escherichia coli, LBmc for S. meliloti, and YPD or SC defined media for Saccharomyces cereviciae. Antibiotics or supplements were added at routine concentrations when appropriate and details are summarized in the Supplemental Methods.
We routinely used a triparental mating protocol to transfer plasmids between donor and recipient strains by conjugation using the E. coli helper strain MT616 to drive mobilization (16). Transfer of extrachromosomal replicons between S. meliloti strains also utilized conjugation; either via an integrated oriT with triparental mating, or through the use of rctB in the donor strain to drive biparental conjugation (1). Deletions were made by FLP-FRT recombination (64), wherein FRT sites were integrated flanking a target region by homologous recombination, and subsequent excision of the region was catalyzed by introduction of FLP recombinase. Alternatively, region capture from S. meliloti involved a quadriparental mating with S. meliloti donor, E. coli helper, E. coli with a FLP-recombinase plasmid, and an E. coli recipient. FLP-FRT recombination was also used to introduce gene clusters into the genome by catalyzing recombination between an FRT site in a plasmid containing the cluster and a single FRT site in a landing pad in the genome (14). We used Φ M12 bacteriophage to transfer insertions between S. meliloti backgrounds using transduction (49). Selection of desired events involved the use of antibiotics as well as prototrophic selection. Further details of genetic manipulations are included in Supplemental Methods. Verification of genetic modifications was performed by PCR using primers in Supplemental Tables S4 to S7 or by whole genome sequencing.
Plasmid Construction
Plasmids for homologous recombination were routinely assembled using Level 1 (BsaI) Golden Gate cloning into suicide destination vectors that follow the BEVA architecture (66). Inserts were PCR amplified with primers found in Supplemental Table S3. Plasmids for gene cluster integration were assembled by yeast recombineering as previously described (66, 67). Primers for amplification of regions that were combined in the assembly are in Supplemental Table S8. Further details of plasmid construction are available in Supplemental Methods. Plasmids were verified by restriction digest or whole plasmid sequencing.
Symbiotic Assays
Plant materials used in this work included Medicago sativa cv. Iroquois (alfalfa) and Medicago truncatula A17 (barrel medic). Plant assays were conducted in Leonard jar assembles with a 1:1 (wt:wt) mixture of sand and vermiculite with 250 mL of Jensen’s medium (68). Four to six seedlings were added to each Leonard jar and inoculated two days after with 10 mL of sterile water containing 100 μL of dense overnight culture. M. sativa seeds were surface sterilized for five minutes with 95% ethanol and for 20 minutes with 2.5% sodium hypochlorite. Seeds were then rinsed with sterile water for one hour, with water replaced every 15 minutes. M. sativa seeds were placed on 1.5% water agar, placed in the dark at room temperature, and allowed two days to germinate. M. truncatula seeds were first scarified with sulfuric acid for 10 minutes. After scarifying, seeds were rinsed 10 times with sterile water and then proceeded to be surface sterilized with 2.5% sodium hypochlorite for two minutes. Seeds were rinsed another 10 times with sterile water. M. truncatula seeds were placed on 1.5% water agar and placed in the dark at 4°C for three days. The seeds were then moved to room temperature and allowed two more days to germinate. Four to six seedlings were added to each Leonard jar and inoculated two days after with 10 mL of sterile water containing 100 μL of dense overnight culture.
Plants were grown in Conviron Gen1000/2000 growth chambers programmed with a day cycle of 18 hours at 21°C with maximum light followed by a night cycle of six hours at 17°C with no light. Plants were allowed 42 days post-inoculation to grow. After the 42-day period, shoots were cut from roots and placed in a drying oven for one week, after which they were weighed for shoot dry weight. Fix^+^ (pink) nodules were picked from roots, counted, and weighed immediately for average nodule fresh weight.
Collected nodules were placed in an air-tight container with desiccant packets and let dry for a week, after which they were weighed for nodule dry weight. In some experiments HP6890 gas chromatograph was used to perform gas chromatography for acetylene reduction assays as previously described (69). This assay was performed immediately after shoots were detached from the root system. A summary of all plant data collected is included in a Supplemental Data file.
Flow Cytometry
Nodules pooled from four plants from one pot were macerated in 750 μL of phosphate-ascorbate buffer (PAB) using a mortar and pestle. An additional 750 μL of PAB was used to rinse and collect any additional bacteroids from the mortar. After vortexing briefly, samples were spun at 200g for 5 minutes to spin down residual plant debris. The resulting supernatant was then pelleted and resuspended in 100 μL of fixative (30% molecular-grade ethanol). After 15 minutes of incubation, samples were pelleted to remove the fixative and resuspended in 500 μL of PBS.
A portion of each sample was pooled to create the three control samples: unstained, DAPI-only, and Nile Red only. Prior to staining, samples were diluted 1:10 in PBS. 1 μL of 10% Nile Red in DMSO and 1 μL of DAPI (1 mg/mL in PBS) were added to samples and allowed 15 minutes to incubate in darkness. Following staining, samples were again diluted 1:10 in PBS, giving a final 1:100 sample dilution. Cells were analyzed on a Beckman Cytoflex S flow cytometer equipped with violet, yellow, red, and blue lasers. The 561 nm yellow laser was used to excite Nile Red, and fluorescence was detected using the 610/20 bandpass filter. DAPI was excited using the 405nm violet laser and detected with the 450/45 bandpass filter. DAPI-positive bacteroids were gated and analyzed for Nile Red fluorescence intensity. Details on the gating schemes are described in Supplemental Figures S19 and S20.
Electron Microscopy
Specimens for transmission electron microscopy were fixed in 2.5% glutaraldehyde in sodium phosphate buffer, postfixed in 2% osmium tetroxide, dehydrated through a graded acetone series including saturated uranyl acetate in 70% acetone, and embedded in Spurr’s resin polymerized at 60°C for 24 hours. Embedded samples were sectioned at 70–90 nm thickness on an RMC MT XL ultramicrotome. Ultrathin sections were collected on copper grids with a formvar-carbon supporting film, then stained with Reynolds’ lead citrate. Observation and imaging were performed on a JEOL JEM-1400 electron microscope operating at 120 kV and equipped with an AMT NanoSprint 15L bottom-mount camera.
GUS Staining and Microscopic Observations
The promoter regions of the pha clusters, nifH, and nodA were cloned via Golden Gate Cloning. Plasmids and strains used are listed in Supplemental Table S1. Primers used for promoter region amplification are listed in Supplemental Table S3. The nodules were fixed in 100 mM sodium-phosphate buffer with 0.1% Triton X-100 and then in 1.5% glutaraldehyde in 100 mM phosphate buffer under vacuum for 30 minutes and 90 minutes. After that, the nodules were embedded in 3% (w/v) low-melting agarose and sectioned into 100-μm slices using vibrating microtome VF-210–0Z (Precisionary, USA). The slices were then vacuum immersed in GUS Staining Solution for 30 min. The solution contains 2 mM 5-bromo-4-chloro-3-indoxyl-β-d-glucuronide cyclohexyl ammonium salt (X-Gluc), 0.2% Triton X-100, and 2 mM each of K3[Fe(CN)6] and K4[Fe(CN)6] in 50 mM sodium phosphate buffer (pH 7.2). After incubating at 37°C for 2 hours, the stained nodule slices were observed using a BioTek Cytation 5 Cell Imaging Multimode Reader (Agilent, USA).
Enzyme Assays
A NADPH/NADH colorimetric assay was used to determine the Km and Vmax values of GhbD reacting with succinate semialdehyde (SSA). Individual 100 μL reactions comprised of purified GhbD (1 μg), NADPH (1 mM) or NADH (1 mM), SSA (0, 0.125, 0.25, 0.5, 1, 2.5, 5, or 10 mM), and reaction buffer (50 mM Na_3_PO_4_ pH 7.0, 200 mM NaCl, 10 mM MgCl_2_). The 340 nm absorbance value of every reaction occurring at 28°C was measured in 1-minute intervals for 11 minutes starting immediately after the addition of GhbD. Absorbance values were read using a BioTek Cytation 5 Cell Imaging Multimode Reader by Agilent. Simple linear regression tests were completed with absorbance values over time for specific substrate concentrations. The resulting slopes, with a R^2^ value > 0.95, were used to determine the rate of reaction per mg of GhbD present. These rates, at different substrate concentrations, were used in the Michaelis-Menten equation to determine the Km and Vmax values for GhbD with SSA and NADPH/NADH. GraphPad Prism Version 10.4.1 was used to perform simple linear regression and non-linear regression tests.
Metabolomics
Metabolomics analysis of plant tissue samples was performed using a 4-channel chemical isotope labeling (CIL) LC-MS method (70). Normalization and labeling kits were obtained from Nova Medical Testing Inc. (Edmonton, AB, Canada). Each tissue sample was homogenized using a bead beater with steel beads, followed by the addition of ice-cold methanol/water (4:1, v/v) for metabolite extraction. The resulting extracts were normalized and labeled according to the protocols provided with the CIL kits.
LC-MS analysis was carried out using a Thermo Vanquish UHPLC system (ThermoFisher, Waltham, MA, US) coupled to a Bruker Impact II QTOF mass spectrometer (Bremen, Germany). Chromatographic separation was performed on an Agilent Eclipse Plus C18 reversed-phase column (150 × 2.1 mm, 1.8 μm particle size) maintained at 40°C. The mobile phases consisted of 0.1% (v/v) formic acid in water (A) and 0.1% (v/v) formic acid in acetonitrile (B). The gradient program was as follows: 0 min, 25% B; 10 min, 99% B; 15 min, 99% B; 15.1 min, 25% B; 18 min, 25% B. The flow rate was 400 μL/min. Mass spectra were acquired at 1 Hz over a mass range of m/z 220–1000. LC-MS data were processed using DataAnalysis (Bruker Daltonics) and IsoMS Pro (Nova Medical Testing Inc.). Manual verification of the key compound gamma-hydroxybutyrate (GHB) was performed using an authentic standard on an alternative instrumental platform. Specifically, MS/MS spectra and retention time (RT) data from the samples were compared to those of the standard, both acquired on a Dionex 3000 LC system coupled with Orbitrap Q Exactive HF mass spectrometer (ThermoFisher, Waltham, MA, US), using the same LC method and column. The MS/MS spectra of GHB in the samples, collected at multiple collision energies, matched those of the authentic standard, confirming the compound’s identity in the sample.
DNA and RNA sequencing
Genome resequencing of hybrid strains and nodule RNA sequencing was performed by the Joint Genome Institute using Illumina NovaSeq technology. For RNA sequencing, bacteroids were isolated from nodules collected directly onto dry ice and flesh frozen in liquid nitrogen from one pot per replicate. RNA was extracted using a QIAgen RNeasy Plus Mini Kit, and underwent on-column DNase treatment. Libraries were prepared using Illumina’s Ribo-Zero rRNA removal kit and TruSeq Standard Total RNA HT sample prep kit. Raw fastq reads were filtered and trimmed according to the JGI QC pipeline, and mapped to the reference genome. Raw read counts were normalized to determine transcripts per million for each gene in the reference. Further details of DNA and RNA extraction, sequencing and bioinformatics analysis are included in Supplemental Methods. Raw data was deposited in NCBI SRA using accessions in Supplemental Table S13. Routine whole plasmid and whole genome resequencing to verify constructs and genetic manipulations was performed by Plasmidsaurus using Oxford Nanopore technology.
Comparative Genomics
Analysis of homology between genes within clusters of interest was performed using clinker (Min. alignment sequence identity 0.3) and visualized with clustermap.js (> 0.5 alignment sequence identity shown) on the CAGECAT web server https://cagecat.bioinformatics.nl/ (71). Whole genome alignments and delineation of locally colinear gene blocks were performed using the MAUVE (72) plugin in the Geneious software with default parameters (automatically calculate seed weight and minimum LCB score with MUSCLE3.6 gapped aligner).
Data Collection and Statistical Analysis
Plant experiments included uninoculated controls that were screened for the absence of nodules to confirm lack of contamination before any data collection. Individual replicates that makeup datapoints or were used for RNAseq or flow cytometry consisted of the average of all plants in one pot (six for M. sativa and four for M. truncatula). For statistical analysis we used a one-way ANOVA test with Dunnett’s multiple comparison test to establish significant differences between replicates. All data shown was significant as determined by one-way ANOVAs. In the figures, results from post hoc significance testing are indicated as *P < 0.0332, **P < 0.0021, ***P < 0.0002, ****P < 0.0001.
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Weisberg A. J., , Pangenome Evolution Reconciles Robustness and Instability of Rhizobial Symbiosis. m Bio 13, e 00074–22 (2022).35416699 10.1128/mbio.00074-22PMC 9239051 · doi ↗ · pubmed ↗
- 2Porter S. S., Simms E. L., Selection for Cheating Across Disparate Environments in the Legume-Rhizobium Mutualism. Ecology Letters 17, 1121–1129 (2014).25039752 10.1111/ele.12318 · doi ↗ · pubmed ↗
- 3Heath K. D., Stinchcombe J. R., Explaining Mutualism Variation: A New Evolutionary Paradox? Evolution 68, 309–317 (2014).24303853 10.1111/evo.12292 · doi ↗ · pubmed ↗
- 4Triplett E. W., Sadowsky M. J., Genetics of Competition for Nodulation of Legumes. Annu. Rev. Microbiol. 46, 399–428 (1992).1444262 10.1146/annurev.mi.46.100192.002151 · doi ↗ · pubmed ↗
- 5Burghardt L. T., di Cenzo G. C., The Evolutionary Ecology of Rhizobia: Multiple Facets of Competition Before, During, and After Symbiosis with Legumes. Current Opinion in Microbiology 72 (2023).
- 6Mendoza-Suárez M., Andersen S. U., Poole P. S., Sánchez-Cañizares C., Competition, Nodule Occupancy, and Persistence of Inoculant Strains: Key Factors in the Rhizobium-Legume Symbioses. Front. Plant Sci. 12, 690567 (2021).34489993 10.3389/fpls.2021.690567 PMC 8416774 · doi ↗ · pubmed ↗
- 7Denison R. F., Legume Sanctions and the Evolution of Symbiotic Cooperation by Rhizobia. The American Naturalist 156, 567–576 (2000).
- 8Westhoek A., , Conditional Sanctioning in a Legume–Rhizobium Mutualism. Proc. Natl. Acad. Sci. U.S.A. 118, e 2025760118 (2021).33941672 10.1073/pnas.2025760118 PMC 8126861 · doi ↗ · pubmed ↗