From Shallow to Full Wrapping: Geometry and Deformability Dictate Lipid Vesicle Internalization
Stijn van der Ham, Alexander Brown, Halim Kusumaatmaja, Hanumantha Rao Vutukuri

TL;DR
This study explores how the shape and flexibility of lipid vesicles affect their engulfment by other membranes, important for processes like drug delivery and cell transport.
Contribution
The research introduces a mechanical model and experimental system to predict and control vesicle engulfment based on geometry and deformability.
Findings
Vesicle engulfment is governed by geometry when size exceeds the bendocapillary length.
Deformability controls transitions between different wrapping states near the bendocapillary length.
Light-responsive lipids enable on-demand switching between wrapping states.
Abstract
The deformability and adhesion of vesicles critically influence their engulfment by lipid membranes, a process that is central to endocytosis, viral entry, drug delivery, and intercellular transport. We developed a versatile experimental system of giant unilamellar vesicles (GUVs) that interact via depletion-induced adhesion. Combining experiments with continuum simulations, we construct a state diagram identifying conditions for the endo- and exocytic engulfment of small GUVs by larger ones. Leveraging full 3D confocal reconstructions of vesicle curvature, we experimentally quantify the bendocapillary length, a scale governing the competition between membrane bending and adhesion. When the vesicle size exceeds this length, wrapping is governed by geometry. In contrast, near this scale, deformability controls transitions between shallow, deep, and fully wrapped states, suppressing full…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1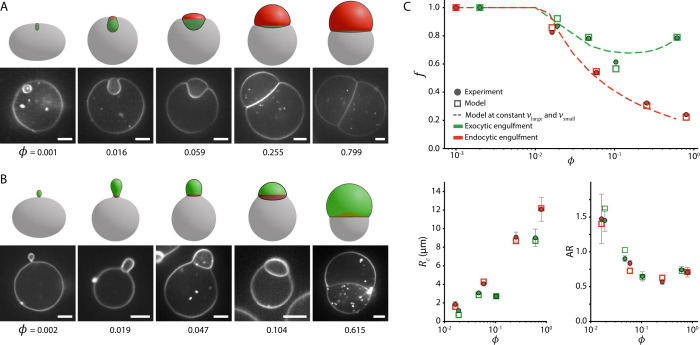 2
2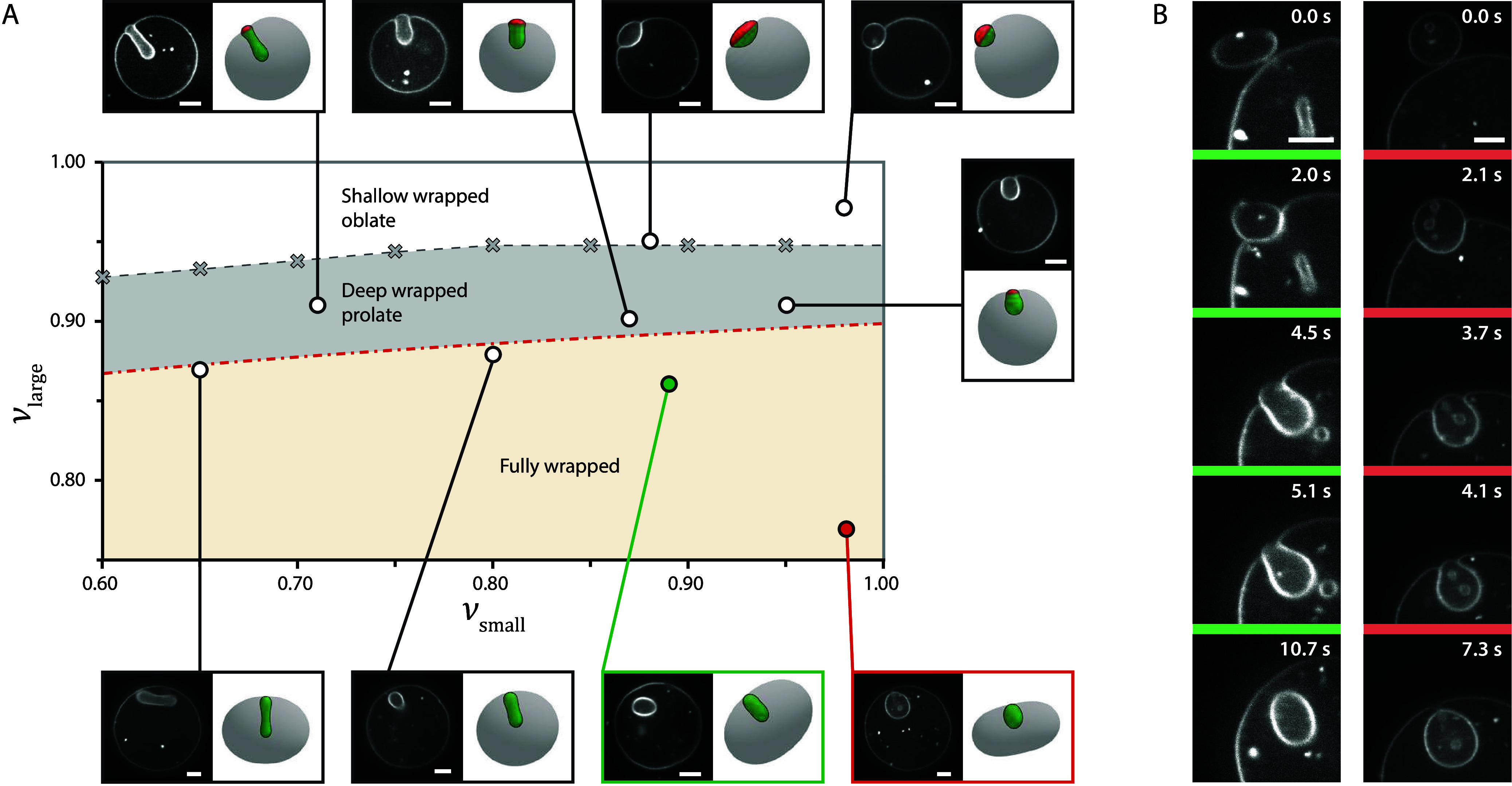 3
3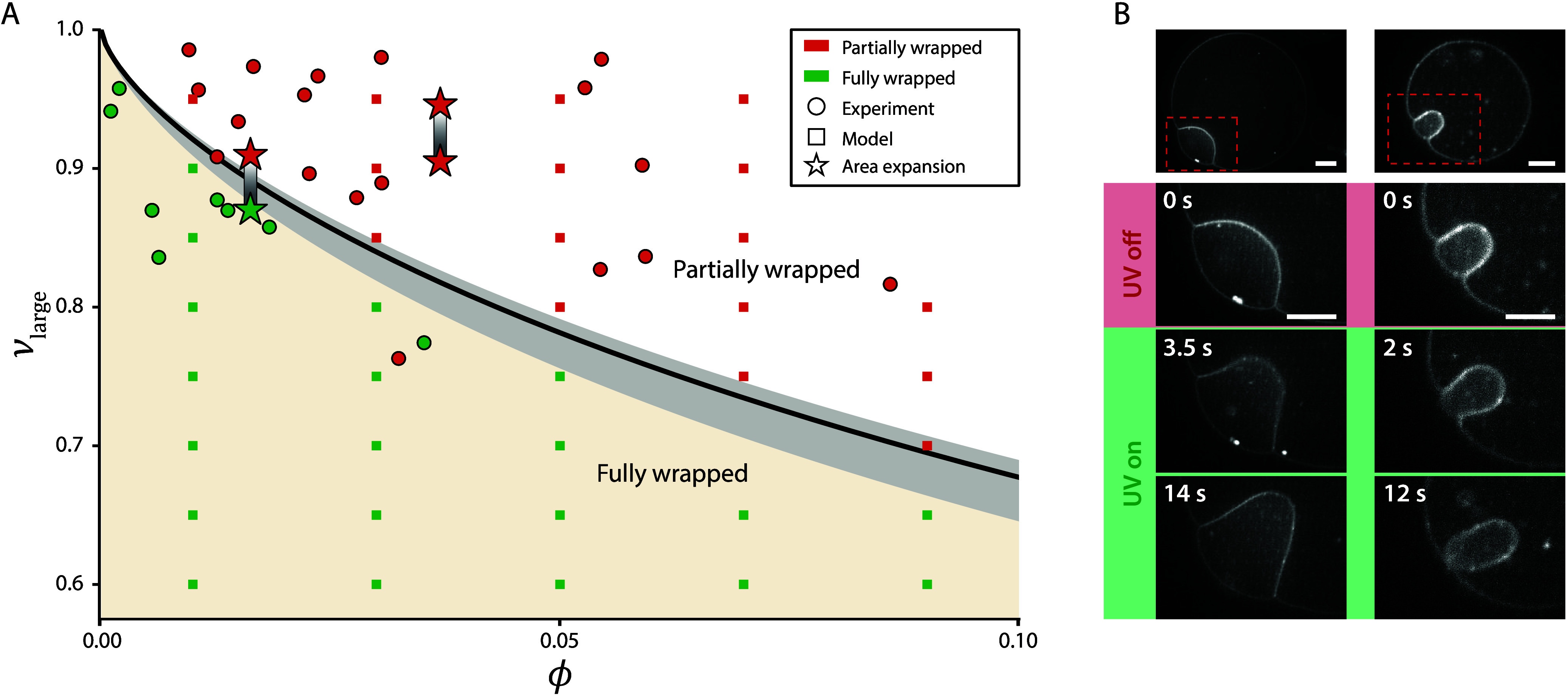 4
4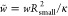 5
5- —Engineering and Physical Sciences Research Council10.13039/501100000266
- —Engineering and Physical Sciences Research Council10.13039/501100000266
- —Nederlandse Organisatie voor Wetenschappelijk Onderzoek10.13039/501100003246
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLipid Membrane Structure and Behavior · Cellular transport and secretion · Supramolecular Self-Assembly in Materials
Vesicle engulfment by lipid membranes is a fundamental mechanism by which cells internalize material, enabling cargo exchange across cellular membranes and playing essential roles in intercellular communication, transport, and immune responses.? In biological systems, extracellular vesicles (EVs) transport molecular cargo between cells, while viruses exploit similar mechanisms to gain entry into host cells. ?−? ? ? In therapeutic contexts, engineered liposomes mimic these mechanisms for targeted drug delivery. ?−? ? While some vesicles cross membranes via direct fusion, many follow wrapping-based pathways such as endocytosis or phagocytosis. ?,?,? Understanding the physical principles underlying these processes is essential for interpreting cellular behavior and developing advanced vesicle-based technologies.
Most studies to date have focused on the engulfment of rigid particles, ?−? ? ? ? ? ? ? where membrane deformation occurs around a fixed particle shape. In contrast, vesicles are deformable, allowing for mutual reshaping during engulfment. Theoretical studies on the engulfment of soft objects by membranes predict that deformability can stabilize partially wrapped states,? inhibit full engulfment,? and drive mutual shape remodelling.? These objects can be distinguished by their deformation modes. For instance, in condensate droplets, soft particles, and lipid vesicles, these modes are respectively determined by droplet surface tension, particle elasticity, and vesicle bending deformation. ?−? ? ? In parallel, experimental studies on nanoparticles demonstrated that particle elasticity affects cellular uptake and circulation time. ?,? Vesicle–vesicle adhesion studies have further shown that contact morphologies between similarly sized vesicles result from a balance between bending elasticity, membrane tension, and adhesion strength. ?−? ? ? ? However, a systematic experimental investigation into the role of deformability in lipid vesicle engulfment remains elusive.
To address this gap, we develop a model system using giant unilamellar vesicles (GUVs),? in which small vesicles interact with larger ones via depletion-driven adhesion. We combine experiments and theory to examine how adhesion and membrane bending govern engulfment and characterize this using the bendocapillary length, a characteristic scale at which bending and adhesion energies balance. When the small vesicle is much larger than this length, wrapping is primarily governed by the geometry. In contrast, when the vesicle size is comparable to this scale, the deformability strongly influences transitions between partially and fully wrapped states.
Experimental
Realization of Vesicle–Vesicle Engulfment
To experimentally investigate vesicle engulfment, we use a biomimetic system where large GUVs engulf smaller ones, mimicking cellular vesicle uptake (Movie S1). GUVs of varying sizes were prepared using the droplet transfer method? see (Supporting Information Section 1). Adhesion between vesicles was induced by suspending them in a solution containing polyacrylamide (0.25 wt %), a nonadsorbing polymer that gives rise to depletion-driven adhesion (see Supporting Information Section 1). This yields an adhesion energy E ad = wA c proportional to the contact area A c, where w is the adhesion strength per unit area. ?,?,?,?
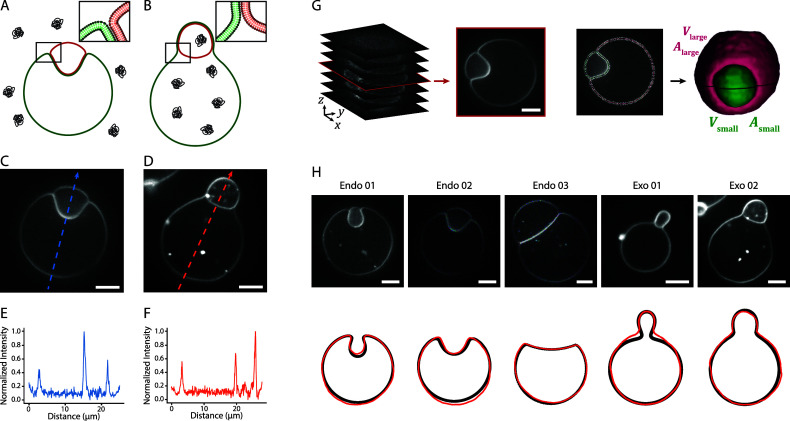
The direction of engulfment can be influenced by adding the polymer to either the outer or inner solution of the GUVs (FigureA,B). When the polymer is added to the outer solution, it creates depletion layers outside the GUVs, promoting adhesion to external vesicles and leading to endocytic engulfment (FigureC). Conversely, adding polymer to the inner solution generates depletion layers inside the GUVs, which can drive adhesion to internal vesicles and lead to exocytic engulfment (FigureD). The direction of engulfment is also reflected in the fluorescence signal of the GUV membranes (FigureC,D). Membrane overlap in the adhered region results in markedly increased fluorescence intensity, appearing as a central peak in the line profile for endocytic engulfment and as an outer peak for exocytic engulfment (FigureE,F).
A, B) Schematic representation of endocytic (A) and exocytic (B) engulfment of a small vesicle (red membrane) by a large vesicle (green membrane) where the polymer is added to the outer and inner solution, respectively. C, D) Confocal fluorescence microscopy images of the midplane of partial endocytic (C) and exocytic (D) vesicle engulfment. E, F) Fluorescence intensity profile along the dashed lines from panel C and D. G) 3D reconstruction of vesicles from confocal fluorescence z-stacks. Image stacks were segmented using the LimeSeg plugin in FIJI (ImageJ), , enabling quantitative extraction of vesicle volume (V), surface area (A), mean curvature (M), and shape. H) The top row shows confocal fluorescence microscopy images of five vesicle pairs that were used for the calibration of L exp. The bottom row shows the corresponding cross-sectional shape comparison between the experimental (red) and simulated (black) large vesicle morphology for L exp = 0.6 μm. Scale bars are 5 μm.
Modeling
the Vesicle–Vesicle Engulfment
To interpret the observed engulfment morphologies, we modeled vesicle–vesicle interactions using energy minimization within a continuum framework (see Supporting Information Section 1), balancing adhesion energy gain and bending energy cost. The model captures the experimental behavior using four dimensionless parameters: the volume ratio of the small and large vesicle, their reduced volumes, and the ratio of the small vesicle size to the bendocapillary length. The volume ratio ϕ is defined as the volume of the small vesicle, V small, divided by that of the large vesicle, V large:
The reduced volume ν_ i _ (i = small, large) quantifies the vesicle deformability and is defined as the ratio between the vesicle’s volume V _ i _ and the volume V _ i,sph_ of a sphere having the same surface area A _ i _:
A vesicle with ν = 1 behaves like a rigid spherical particle. As ν decreases, the excess membrane area increases, allowing the vesicle to adopt a broader range of nonspherical shapes, thus increasing its deformability.?
Finally, the ratio R small/L characterizes the balance between adhesion and membrane bending forces, where R small is given by
and L is defined as
with κ the bending rigidity of the membrane. For R small/L ≫ 1, adhesion and surface tension forces dominate, while for R small/L ≪ 1, bending rigidity dominates.
We determined ϕ, ν_large_, and ν_small_, by measuring the volume and surface area from 3D confocal microscopy reconstructions using a contour segmentation algorithm (FigureG and Supporting Information Section 1). The experimental bendocapillary length was determined by comparing vesicle curvature profiles with model predictions. For five vesicle pairs (FigureH), we replicated ϕ, ν_large_, and ν_small_ in simulations, while treating the bendocapillary length L sim as a tunable parameter. The L sim value for which the simulated vesicle’s curvature best matched the experimental curvature was identified and used to calculate
(details in Supporting Information Section 1). Averaging across five vesicle pairs yielded L exp = 0.6 ± 0.1 μm, which is used throughout our analysis. Cross-section comparisons between experiments and simulations are shown in FigureH, demonstrating close agreement.
Although L exp should be approximately constant for a given polymer concentration, the vesicle size R small,exp varies, causing changes in R small,exp/L exp. Therefore, for each vesicle pair, we replicate R small/L in simulations, enabling direct comparison to experiments. This framework allows us to systematically investigate the role of the four dimensionless parameters in vesicle engulfment.
Endo-
and Exocytic Engulfment of Vesicles with Varying Volume Ratio
Combining experiments and continuum simulations, we begin by studying how the direction of engulfment influences vesicle morphology (FigureA,B). We explore this behavior by studying vesicle pairs across a range of volume ratios (ϕ = 0.001–0.799), while keeping their reduced volumes approximately constant. Because of experimental variability, we limited our analysis to vesicle pairs with reduced volumes between 0.87 and 0.97 for both large and small vesicles. We employ the same polymer concentration of 0.25 wt % polyacrylamide, which should lead to a constant adhesion strength. However, due to vesicle size variation, the ratio R small/L ranges from 1.8 to 20.
Endo- (A) and exocytic (B) vesicle engulfment as a function of volume ratio ϕ. Confocal fluorescence microscopy images representing equilibrium conformations at various ϕ (indicated below the images). All vesicles had a reduced volume between 0.87 and 0.97 and were recorded at the same adhesion strength (0.25 wt % polyacrylamide). The ratio R small/L ranges from 1.8 to 20. For endocytic engulfment (A), the polymer was present in the outer solution, whereas for exocytic engulfment (B), the polymer was present in the inner solution. The corresponding morphologies, replicated by the model, are shown above the microscopy images. The large vesicle is represented in gray, the small vesicle in red, and the contact area is highlighted in green. Scale bars are 5 μm. C) Quantitative comparison between experiments and simulations for the vesicles shown in panels A (red) and B (green). In all plots, circles denote experimental data and squares denote simulated counterparts. Top panel: wrapping fraction f as a function of ϕ. Dashed lines indicate simulated f-values for endocytic (νlarge = 0.9 and νsmall = 0.94) and exocytic (νlarge = 0.92 and νsmall = 0.92) engulfment, with R small/L = 5 held constant. Bottom panels: Contact-line radius R c and small vesicle aspect ratio AR as a function of ϕ for the partially wrapped vesicles. Error bars represent the standard deviation in the experimental data, originating from the variation around the symmetry axis.
During interaction, the membranes’ flexibility drives both vesicles to remodel each other, minimizing the system’s energy. In endocytic engulfment (FigureA), the large vesicle forms an invagination at the contact site that deepens with decreasing ϕ. Conversely, in exocytic engulfment (FigureB), a protrusion forms around the smaller vesicle. In both cases, the small vesicle undergoes shape changes as it becomes increasingly wrapped, transitioning from an oblate form in the shallow-wrapped state to a prolate geometry in the deep wrapped state. As the wrapping fraction approaches 1, a narrow catenoidal membrane neck forms, creating either an inward-facing bud (endocytic) or an outward-facing bud (exocytic) that fully encapsulates the smaller vesicle.
In the fully wrapped state (leftmost panels of FigureA,B), small discrepancies appear between experiments and simulations. Experimentally, large vesicles are slightly flattened, whereas simulations predict a more elongated, prolate shape. This deviation stems from gravity due to a small density mismatch between the inner and outer solutions (osmotically matched sucrose versus glucose), which causes size-dependent flattening of large vesicles.? We further quantified this effect in Supporting Information Section 2.
To further validate our observations, we compared experimental vesicle shapes with model predictions using three independent geometric observables: the wrapping fraction f = A c/A small, the contact line radius R c, and the small-vesicle aspect ratio AR (FigureC; see Supporting Information Section 3 and Figure S5). The open symbols correspond to simulations matched to the measured dimensionless variables (ϕ, ν_large_, ν_small_, R small/L, where L exp = 0.6 μm), while the dashed lines show representative trends obtained for fixed parameter sets. Across both endo- and exocytic regimes, the model and experiments show good quantitative agreement in all three observables, supporting the validity of the mechanical framework.
Endocytic Vesicle
Engulfment and the Role of Vesicle Reduced Volume
While our system enables the study of both endo- and exocytic engulfment (Figure), we focus on endocytic uptake to explore how vesicle deformability influences the wrapping morphology. We map vesicle–vesicle shapes as a function of ν_small_ and ν_large_ at an approximately constant volume ratio ϕ (FigureA). Here, ν_small_ sets the deformability: even slight deviations from ν_small_ = 1 enable the small vesicle to adapt to the curvature imposed by the large vesicle. In contrast, ν_large_ reflects the excess membrane area available for wrapping and primarily governs the transitions among shallow, deep, and fully wrapped states. Within each regime, ν_small_ shapes the morphology. In the shallow wrapped state, small vesicles with a lower reduced volume appear more oblate, lying flatter against the large vesicle, whereas in the deep wrapped state they transition toward a more prolate form oriented perpendicular to the membrane (FigureA). In the fully wrapped state, the small vesicle recovers the morphology of a free vesicle with the same reduced volume but enclosed by the larger vesicle’s membrane. In this regime, minor differences between experiments and simulations arise from size-dependent gravitational flattening (Supporting Information, Section 2).
State diagram of endocytic vesicle engulfment as a function of νlarge and νsmall. A) Vesicle-vesicle morphologies as a function of νlarge (y-axis) and νsmall (x-axis). Confocal fluorescence microscopy images represent typical equilibrium conformations of vesicle pairs for various reduced volumes. The corresponding morphologies that were replicated by the model are shown alongside the microscopy images. All vesicle pairs had a volume ratio between 0.013 and 0.037 and were recorded at the same adhesion strength (0.25 wt % polyacrylamide). The ratio R small/L ranges from 3 to 7.3. The shaded areas represent shallow wrapped oblate vesicles (white), deep wrapped prolate vesicles (gray), and fully wrapped vesicles (sand). The transition between the shallow and deep wrapped states (gray dashed line) was obtained from simulations (gray crosses). The transition to the fully wrapped state (red dash-dotted line) was calculated with eq using ϕ = 0.016. B) Time series captured with confocal fluorescence microscopy, illustrating the morphology of two vesicles (green: νsmall = 0.89 and red: νsmall = 0.98 from panel A) as they transition from the free (f = 0) to the fully engulfed state (f = 1). Scale bars are 5 μm.
The occurence of shallow-wrapped oblates and deep-wrapped prolates reflects the orientational stability predicted for rigid ellipsoids. ?,? However, unlike rigid ellipsoids or rods, which reorient during wrapping, ?−? ?,? deformable vesicles can continuously reshape themselves, transitioning from oblate to prolate as the wrapping fraction increases. ?,? This dynamic adaptation is further illustrated by the continuous shape adaptations observed during the transition of a small vesicle from the free to fully wrapped state, as shown in FigureB and Movies S2 and S3 for two different values of ν_small_, corresponding to the vesicle pairs in FigureA marked with green (ν_small_ = 0.89) and red (ν_small_ = 0.98).
Geometrically Constrained Engulfment
To further characterize the onset of full engulfment, we examine how this transition depends on the vesicle geometry. In the R small/L ≫ 1 regime, the full engulfment transition is determined primarily by the system’s geometry: ϕ, ν_large_, and, to a lesser extent, ν_small_. As the small vesicle becomes wrapped, part of the large vesicle’s excess membrane area is consumed, increasing its sphericity, while the remaining membrane must enclose the combined volume of both vesicles. The effective reduced volume of the large vesicle after wrapping a small vesicle is thus: ?,?
which can be rewritten in terms of the dimensionless parameters as (see Supporting Information Section 4):
The value of ν_γ_ quantifies whether full wrapping is possible: when ν_γ_ ≤ 1, the small vesicle can be fully engulfed, whereas when ν_γ_ > 1, the large vesicle lacks sufficient excess membrane area to fully engulf the small vesicle, resulting in partial wrapping. Interestingly, eq shows that the engulfment transition depends only weakly on ν_small_, since it is scaled by ϕ^2/3^, which is small in our study (ϕ < 0.1). This weak dependence highlights that in this regime the small vesicle deformability plays a limited role.
We constructed a state diagram mapping the wrapping outcome as a function of ϕ and ν_large_ (FigureA), combining experiments, simulations, and the theoretical transition from eq. The narrow shaded region representing ν_small_ = 0.7 to 1 confirms the weak influence of the small vesicle deformability. Experimental data spanning R small/L = 1.8 to 11 align well with the predicted transition, indicating that even modest increases beyond R small/L = 1 are sufficient to reach the geometry-dominated regime. Simulations at R small/L = 10 closely match the analytical prediction, supporting this conclusion.
A) State diagram of the partially and fully wrapped state as a function of νlarge and ϕ. The black line gives the theoretical transition between the partially (red) and fully wrapped (green) state based on eq . The gray shaded area indicates the influence of νsmall on this transition. Based on the available experimental data, we show a lower and an upper bound of νsmall = 0.7 and 1.0, respectively, and the black line representing νsmall = 0.9. Experimental and simulated data points are plotted as circles and squares, respectively. For the simulated data, R small/L = 10 and νsmall = 0.9, while for the experimental data R small/L ranges from 1.8 to 11 and νsmall ranges from 0.65 to 0.99. Area expansion estimates corresponding to panel B are given by pairs of stars, where the top star corresponds to the vesicle pair before UV illumination, and the bottom star to the vesicle pair after UV illumination. B) Confocal fluorescence microscopy images of two vesicles containing azo-PC before (red) and during (green) UV exposure. The increase in membrane area during UV exposure drives a transition from shallow to deep wrapped (left column) and from deep to fully wrapped state (right column). Scale bars are 10 and 5 μm for the left and right column, respectively.
Equation and FigureA also predict that decreasing ν_large_ promotes full wrapping. To test this hypothesis, we incorporated photoresponsive azo-PC lipids into GUV membranes? (Supporting Information Section 1). Upon UV illumination, azo-PC isomerizes, increasing membrane area and lowering ν_large_.? This light-triggered modulation enabled active control of engulfment, driving transitions from shallow to deep, and from deep to full wrapping (Figure, Movies S4 and S5), consistent with recent reports on photoswitchable endocytosis of biomolecular condensates.?
Repeated UV illumination of the vesicles shown in FigureB induced reversible transitions between shallow and deep wrapping, whereas transitions to the fully wrapped state were not reversible (Figure S6). We attribute this asymmetry to the high energetic cost of reopening the membrane neck in combination with a relatively small area change (Supporting Information Section 1). However, further investigation of this asymmetry is warranted to fully understand the underlying mechanisms, as repeated illumination often led to the formation of narrow internal membrane tubes that consumed the membrane area and drove the vesicles toward more spherical shapes.
While the majority of the experimental data conform to the predicted transition, one data point near ϕ ≈ 0.035 remained partially wrapped despite satisfying ν_γ_ < 1. Comparison with a neighboring fully wrapped vesicle reveals the difference lies in their R small/L value: R small/L = 7.3 for the fully wrapped vesicle versus 3.0 for the partially wrapped one. This suggests that the outlier lies near the edge of the geometry-dominated regime, where additional factors such as bending energy may inhibit full engulfment.
Adhesion
Constrained Engulfment
Having established that vesicle geometry primarily dictates the engulfment transition in the R small/L ≫ 1 regime, we now consider the regime where R small/L ≈ 1. Here, the vesicle geometry is insufficient to predict the wrapping outcome. Although full engulfment can still occur when ν_γ_ < 1, the interplay between bending rigidity and adhesion introduces additional constraints. As a result, full wrapping may be suppressed by the deformability of the small vesicle (ν_small_).
Figure shows the full engulfment transition as a function of R small/L (or equivalently ) and ν_γ_. We fix ν_large_ = 0.75 and examine three values of ν_small_: 0.8, 0.95, and 0.99, while varying ν_γ_ by changing ϕ (see eq). The partial to full wrapping transition is calculated from the total energy curves as a function of f (Figure S7). The results show that full engulfment requires an increasing adhesion strength as ν_small_ decreases, illustrating that increased deformability inhibits complete wrapping. This behavior is consistent with prior findings for planar membranes and volume-unconstrained vesicles. ?,? Notably, the two experimental points discussed previously, both at ν_γ_ = 0.93 but with R small/L values of 7.3 and 3.0, fall into the fully and partially wrapped regimes, respectively, consistent with their observed wrapping states.
Partial to full wrapping transition as a function of R small/L (left axis), or equivalently w̃=wRsmall2/κ (right axis), and νγ. The transition is shown for constant νlarge = 0.75, while νsmall is varied: 0.8 (green), 0.95 (blue), and 0.99 (red). The vertical gray dashed line indicates the geometric cutoff at νγ = 1, beyond which full engulfment cannot occur.
For all ν_small_, the transition curves show a plateau at low ν_γ_, followed by a steep decline as ν_γ_ → 1. In the absence of spontaneous curvature, the bending energy is scale-independent. Consequently, the bending energy preengulfment depends only on the vesicle shape and is independent of ϕ and ν_γ_. In contrast, as ν_γ_ increases, the large vesicle becomes more spherical after engulfing the small vesicle, reducing the bending energy cost and lowering the adhesion required for full wrapping. This can be observed in the results shown in Figure S7. The effect is strongest near ν_γ_ = 1, where the bending energy gain is largest, and is consistent with previous theoretical studies.? Figure S7 further compares the influence of ν_small_ on this effect. For ν_small_ = 0.8, the wrapping energy curves change little with ν_γ_, giving a gradual decrease in the adhesion threshold as ν_γ_ → 1. For ν_small_ = 0.99, the curves vary strongly with ν_γ_, producing a sharper decrease in the adhesion threshold.
Conclusions
In this study, we introduced a biomimetic system of large GUVs engulfing smaller ones to investigate how vesicle deformability affects vesicle–vesicle uptake. Adhesion was mediated by depletion interactions, providing a simplified yet biologically relevant analog of cellular internalization. By selectively placing the depletant inside or outside the GUVs, we could control the directionality of engulfment, observing both endocytic and exocytic pathways in the same system.
Our experimental and theoretical analyses show that engulfment is governed by the interplay of adhesion and bending energies, characterized by the bendocapillary length. When R small/L > 1, wrapping is geometry-dominated: whether the small vesicle is partially or fully engulfed depends primarily on the large vesicle’s excess membrane area, quantified by ϕ, ν_large_, and, to a lesser extent, ν_small_. In contrast, when R small/L ≈ 1, the transition also depends on the deformability of the small vesicle, quantified by ν_small_. More deformable vesicles (lower ν_small_) tend to resist complete wrapping, requiring stronger adhesion to achieve full engulfment.
While the complexity of cellular uptake cannot be captured by a single characteristic length, the concept of a bendocapillary length provides physical insight into how adhesion and membrane bending govern the engulfment of deformable vesicles. For biological membranes with bending rigidities of few tens of k B T, and EVs ranging from tens of nanometres to several micrometres in size,? adhesion energies spanning weak nonspecific interactions to strong receptor–ligand binding (10^–8^ to 10^–4^ J m^–2^)? yield bendocapillary lengths of 10–1000 nm, comparable to EV dimensions. This suggests that many biological systems operate in a regime where deformability might influence uptake.
This study investigates ν_small_ values ranging from ∼0.6 to ∼1, which is comparable to those observed in EVs? and pleiomorphic viruses such as influenza.? Within this range, we find that vesicle deformability significantly influences engulfment, emphasizing both its biological relevance and the design principles it offers for soft drug delivery carriers.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1György B.SzabóT. G.Pásztói M.Pál Z.Misják P.Aradi B.LászlóV.Pállinger E.Pap E.Kittel A.Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles Cell. Mol. Life Sci.201168266710.1007/s 00018-011-0689-321560073 PMC 3142546 · doi ↗ · pubmed ↗
- 2Arandjelovic S.Ravichandran K. S.Phagocytosis of apoptotic cells in homeostasis Nature Immunology 20151690710.1038/ni.325326287597 PMC 4826466 · doi ↗ · pubmed ↗
- 3Cohen F. S.How viruses invade cells Biophys. J.2016110102810.1016/j.bpj.2016.02.00626958878 PMC 4788752 · doi ↗ · pubmed ↗
- 4Marsh M.Helenius A.Virus entry: open sesame Cell 200612472910.1016/j.cell.2006.02.00716497584 PMC 7112260 · doi ↗ · pubmed ↗
- 5Allen T. M.Cullis P. R.Liposomal drug delivery systems: from concept to clinical applications Adv. Drug Delivery Rev.2013653610.1016/j.addr.2012.09.03723036225 · doi ↗ · pubmed ↗
- 6Guimarães D.Cavaco-Paulo A.Nogueira E.Design of liposomes as drug delivery system for therapeutic applications Int. J. Pharm.202160112057110.1016/j.ijpharm.2021.12057133812967 · doi ↗ · pubmed ↗
- 7Bareford L. M.Swaan P. W.Endocytic mechanisms for targeted drug delivery Adv. Drug Delivery Rev.20075974810.1016/j.addr.2007.06.008PMC 200032917659804 · doi ↗ · pubmed ↗
- 8Joshi B. S.de Beer M. A.Giepmans B. N.Zuhorn I. S.Endocytosis of extracellular vesicles and release of their cargo from endosomes ACS Nano 202014444410.1021/acsnano.9b 1003332282185 PMC 7199215 · doi ↗ · pubmed ↗