Chromatin remodeling complexes: architects influencing breast cancer progression
Octavianus Giovani, Grace S. Eckersley, Hayden R. Jones, Sankari Nagarajan

TL;DR
This paper explores how chromatin remodeling complexes influence breast cancer progression and their potential as therapeutic targets.
Contribution
The paper highlights the role of chromatin remodeling complexes in breast cancer and their potential for targeted therapies.
Findings
Chromatin remodeling complexes are divided into four subfamilies and influence breast cancer progression.
Targeted therapies using ATPase inhibitors or proteasome-based degraders show promise in controlling tumor growth.
Further research is needed to understand the mechanisms of these complexes for therapeutic strategies.
Abstract
As one of the most common types of cancer, breast cancer strongly contributes to the increase in morbidity and mortality worldwide. Alterations in the genetic and epigenetic landscape contribute to the complexity and heterogeneity of the disease, making its understanding and prognosis more challenging. Chromatin remodeling complexes are implicated as essential factors driving the progression and aggressiveness of breast cancer by permitting chromatin dynamics to promote or suppress transcription. Based on their structure and biochemical properties, chromatin remodeling complexes are divided into four subfamilies: SWI/SNF, ISWI, CHD and INO80. Due to their involvement in breast cancer progression, these complexes present potential therapeutic targets, either through direct or indirect approaches. Several promising efforts have been made to develop targeted therapies against chromatin…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
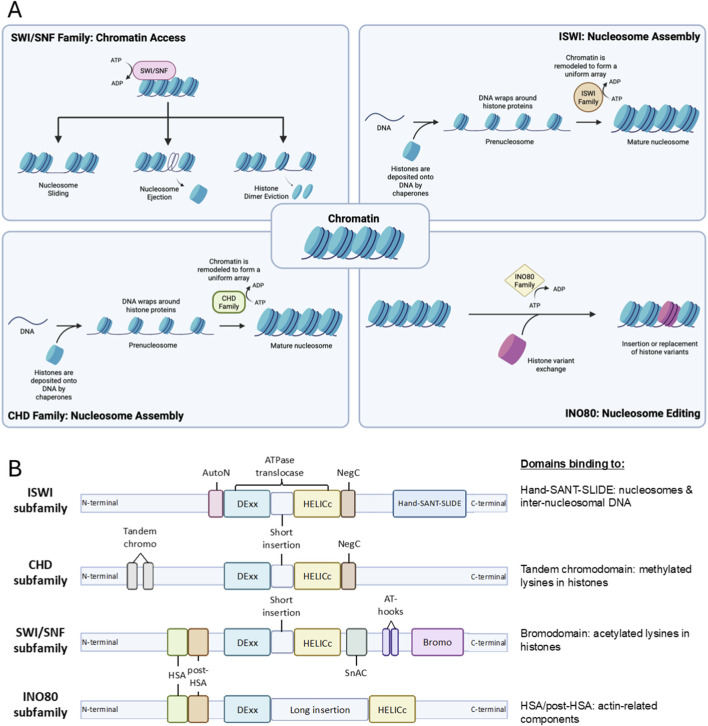 FIGURE 1
FIGURE 1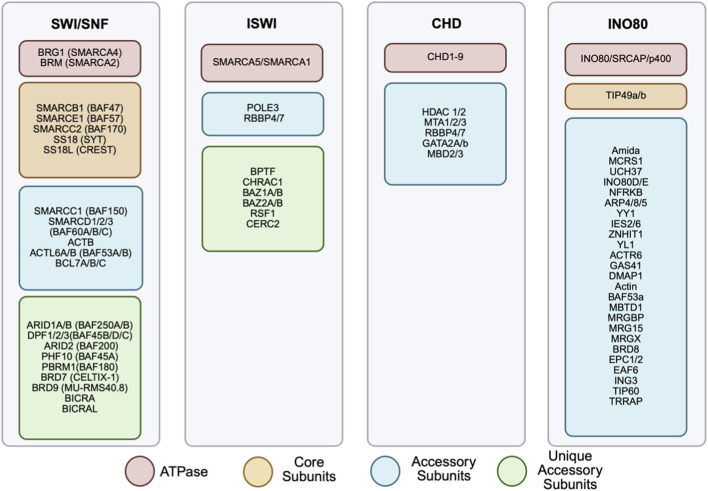 FIGURE 2
FIGURE 2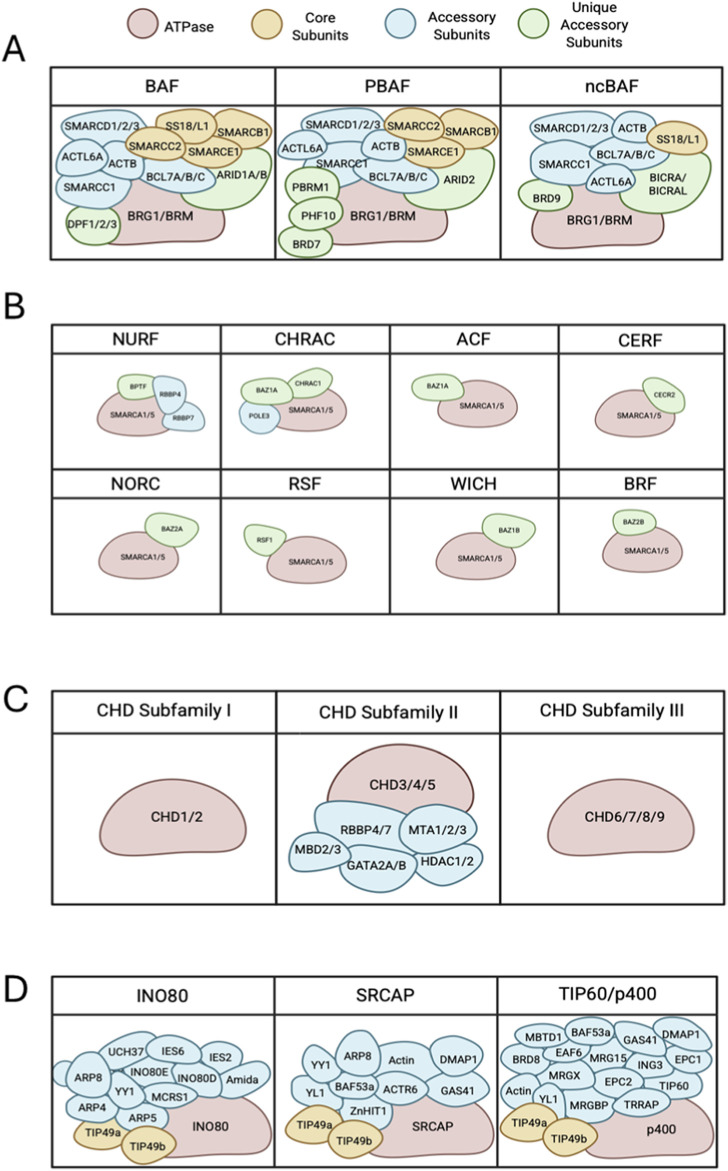 FIGURE 3
FIGURE 3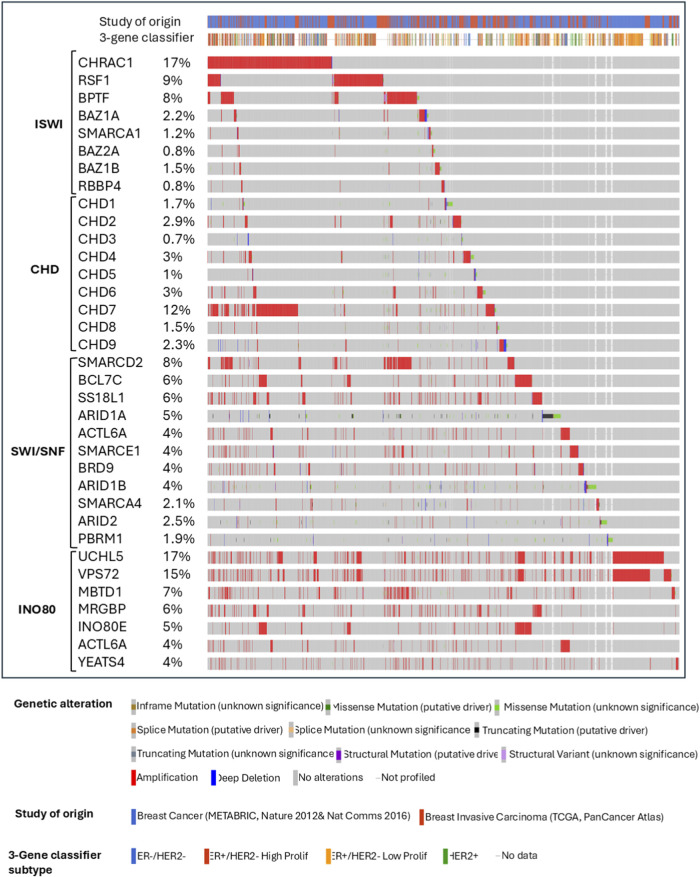 FIGURE 4
FIGURE 4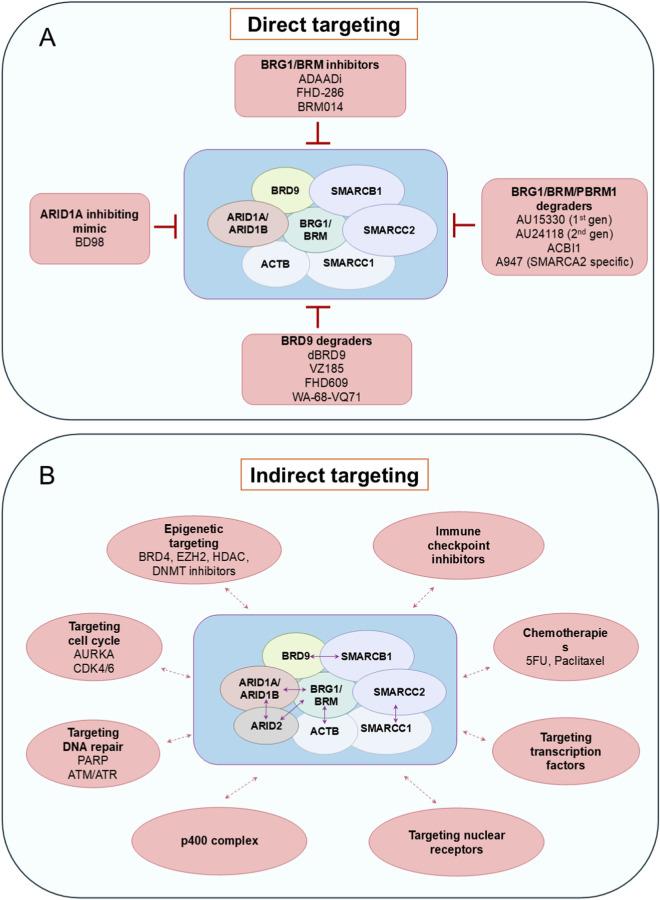 FIGURE 5
FIGURE 5| Complex | Subunit | Key results | Proposed functional classification | References |
|---|---|---|---|---|
| SWI/SNF | BRG1/BRM | Functional studies: | Oncogenic |
|
| ARID1A | Functional studies: | Tumor suppressive |
| |
| ARID1B | Functional studies: | Oncogenic |
| |
| PBRM1 | Functional studies: | Tumor suppressive |
| |
| ARID2 | Non-functional studies: | Tumor suppressive |
| |
| ISWI | SMARCA5 | Functional studies: | Oncogenic |
|
| SMARCA1 | Functional studies: | Oncogenic |
| |
| BAZ1A | Functional studies: | Oncogenic |
| |
| BPTF | Functional studies: | Oncogenic |
| |
| CECR2 | Functional studies: | Oncogenic |
| |
| CHD | CHD1 | Functional studies: | Oncogenic |
|
| CHD3 | Non-functional studies: | Tumor suppressive |
| |
| CHD4 | Functional studies: | Oncogenic |
| |
| CHD5 | Non-functional study: | Tumor suppressive |
| |
| CHD7 | Non-functional studies: | Oncogenic |
| |
| CHD9 | Non-functional studies: | Tumor suppressive |
| |
| INO80 | INO80 | Non functional studies: | Tumor suppressive |
|
| SRCAP | Functional studies: | Oncogenic |
|
- —Cancer Research UK10.13039/501100000289
- —Breast Cancer Now10.13039/501100007913
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromatin Remodeling and Cancer · Genomics and Chromatin Dynamics · Protein Degradation and Inhibitors
Introduction
1
Breast cancer is one of the most common cancers in the world, with its morbidity and mortality rates increasing over time (Sung et al., 2021; Siegel et al., 2023). While the burden of other cancers, such as lung cancer and melanoma, is predicted to decrease, breast cancer is expected to create more impact on patients and healthcare systems by 2040 (Arnold et al., 2022). In the UK, there were around 58,000 new cases and 12,000 deaths related to breast cancer in 2022, presenting it as the most common type of cancer (Ferlay et al., 2024). Furthermore, there were a total of 2.3 million new cases and 685,000 deaths associated with breast cancer globally in 2020. These numbers are predicted to climb to over 3 million new cases and 1 million deaths annually by 2040 (Arnold et al., 2022).
Breast cancer subtypes
2
Breast cancer is a disease with high heterogeneity, multiple subtypes, and a variety of risk factors, which altogether challenge the diagnosis and treatment of patients, as the different subtypes require different types of therapy. Aggressive breast cancers are associated with metastasis, treatment relapse, and high mortality, hence possessing lower survival rates (Arnold et al., 2022; Afifi and Barrero, 2023). The aggressiveness of breast cancer can be predicted by its subtype, which is based on the expression of the estrogen receptor (ER), progesterone receptor (PR), and the human EGF receptor 2 (HER2). The combination of expression of these three receptors can be used to group breast cancer tumours into four main molecular subtypes: luminal A, luminal B, HER2-enriched, and triple-negative breast cancer (TNBC).
Luminal A cancers show expression of ER and PR (ER+/PR+) but lack HER2 (HER2-) and typically have low numbers of cells exhibiting proliferative markers (e.g., Ki67). They have the most favourable prognosis, as they respond well to ER-targeting therapies. Luminal B cancers are also ER+ and PR+, but they can be either HER2+ or HER2-. They often present with larger, faster-growing tumours with increased Ki67 detection and a slightly worse prognosis than luminal A. HER2 cancers are ER- and PR- but HER2+. They have high rates of proliferation, and patients with these cancers have markedly increased survival rates since the advent of HER2-targeting therapies. TNBC tumours are ER-, PR- and HER2-. They are the most aggressive breast cancers, with poor survival, early metastasis, and limited treatment options (Afifi and Barrero, 2023; Lee et al., 2023).
With the growing knowledge from sequencing technologies, it becomes apparent that in addition to genetic alterations in key proliferative genes (ER, HER2), dysregulation in the epigenetic landscape plays a significant role in driving breast cancer aggressiveness. One type of protein complexes that has been implicated in the epigenetic dysregulation are chromatin remodeling complexes. These complexes are crucial in mediating gene transcription, and alterations in the genes encoding these complex proteins may contribute to oncogenesis and cancer progression several cancers, such as prostate, hepatocellular, pancreatic, colorectal, and oral squamous cell carcinoma (Fang et al., 2011; Pancione et al., 2013; Kumar et al., 2016; Ordonez-Rubiano et al., 2024). This review aims to elaborate in-depth on the functions of the chromatin remodeling complexes, explore their roles in driving progression and aggressiveness in breast cancer, and discuss how inhibitors of chromatin-modifying proteins have shown promise as an alternative treatment for breast cancer (Table 1).
Chromatin remodeling complexes
3
Gene expression is vital for cells to function and survive. However, the intricacy of chromatin architecture and the position of nucleosomes along the DNA strands limit the accessibility of DNA for transcription factor binding and therefore control gene expression. The chromatin remodeling complexes are comprised of enzymes and proteins that regulate chromatin accessibility by opening or closing the chromatin to execute their functions on replication, gene expression, repair, and recombination (Kadoch and Crabtree, 2015). These complexes function by establishing the proper density and spacing of nucleosomes, which leads to the packaging and unpackaging of chromatin during the aforementioned processes (Clapier and Cairns, 2009; Clapier et al., 2017). Hence, based on their function, remodelers can be classified into three groups: nucleosome assembly and organization, chromatin access, and nucleosome editing (Figure 1A) (Flaus et al., 2006; Clapier et al., 2017).
Schematic depiction of chromatin remodeling complexes. (A) General functions of chromatin remodeling complexes, which include nucleosome maturation assembly and spacing to form mature nucleosome by ISWI and CHD complexes, promoting chromatin acssibility by enabling chromatin repositioning through nucleosome sliding, nucleosome ejection and histone dimer eviction by SWI/SNF complex, and nucleosome editing via insertion or canonical and variant histone performed by INO80 complex. (B) Structure of each subfamily proteins with their domains. Each possesses split ATPase domains composed of Dexx and HELICc segments, with different domains unique to each subfamily. HSS and Bromo domains near the C-terminals of ISWI and SWI/SNF subfamilies bind to nucleosomes and histones of interest. Tandem chromo and HSA/post-HSA domain adjacent to N-terminals of CHD and INO80 subfamilies bind to methylated lysines in histone and actin-related components, respectively.
Most remodeling complexes use ATP hydrolysis to alter the histone-DNA contacts. They are more extensively studied and share several properties that enable the engagement, selection, and remodeling of the nucleosome. These properties include the affinity for nucleosomes compared to the DNA elements, domains that recognize covalent histone modifications, catalytic subunits with ATPase domains to break histone-DNA contacts, domains that regulate the ATPase domain, and domains for interaction with other chromatin or transcription factors for selective action on particular nucleosomes at specific sites (Clapier and Cairns, 2009; Clapier et al., 2017).
This review will focus on the members of ATP-dependent chromatin remodeling complexes comprising SWItch/Sucrose Non-Fermentable (SWI/SNF), Imitation SWItch (ISWI), Chromodomain Helicase DNA-binding protein (CHD), and INOsitol-requiring mutant 80 (INO80) remodeler families (Figure 2). Despite their similar properties, they possess differences in ATPase structure and complex constituencies (Figure 1B). For example, the ATPase domains of all remodeler families are divided into DExx and HELICc segments, but the INO80 family has a unique longer insertion domain between the two segments. Other distinguishing features are the differences in the combinations of flanking domains within the remodeler families, such as bromodomain for the SWI/SNF family, Helicase and SWI-3 adaptor 2 Nuclear receptor co-repressor Transcription factor IIIB (SANT)-associated (HSA) domain for the SWI/SNF and INO80 families, HAND-SANT-SLIDE (HSS) module for the ISWI family and tandem chromodomains for the CHD family. These domains are evolutionarily conserved in protein composition and function (Clapier and Cairns, 2009).
Subunits composition of chromatin remodeling complexes. The four chromatin remodeling complexes: SWI/SNF, ISWI, CHD and INO80, are represented with their respective subunits listed. Each family is made up of a selection of subunits including ATPase (red), core subunits (yellow), accessory subunits (blue) and unique accessory units (green). Different combinations of each subunit can result in differing complex actions. Several of these subunits have been implicated in cancer.
SWI/SNF subfamily
3.1
SWI/SNF remodelers can slide nucleosomes along DNA, evict nucleosome components and eject whole nucleosomes, rendering the chromatin more accessible to proteins and RNA. This exposes binding sites for transcription factors, coactivators or repressors as well as DNA repair and recombination factors (Boeger et al., 2004; Clapier and Cairns, 2009; Clapier et al., 2017).
Structure and composition
3.1.1
The SWI/SNF family is composed of a central catalytic subunit with 8–14 associated subunits. The catalytic subunit possesses an ATPase-translocase domain formed by two RecA-like lobes that flank a small, conserved insertion, surrounded by an N-terminal HSA domain that binds to actin or actin-related proteins (ARP), post-HSA domain, AT-hooks, and C-terminal bromodomain (Figure 1B) (Mohrmann and Verrijzer, 2005; Kasten et al., 2011; Schubert et al., 2013; Clapier et al., 2017). There are three SWI/SNF complexes in humans: BRG1-associated Factors (BAF, SWI/SNF-A), Polybromo-associated BAF (PBAF, SWI/SNF-B), and the recently defined non-canonical BAF (ncBAF) (Xue et al., 2000; Mohrmann and Verrijzer, 2005; Mani et al., 2017). Each functional complex can only possess one catalytic subunit, either Brahma [BRM/SWI/SNF related, matrix-associated, actin-dependent regulator of chromatin, subfamily A, member 2 (SMARCA2)] or Brahma-Related Gene (BRG1/SMARCA4). The rest of the complex is formed by core subunits, which are essential for remodeling activity, and accessory subunits that target or regulate the complex activity (Figures 2, 3A).
Subunit and domain arrangement of chromatin remodeling complex subfamilies. (A) All SWI/SNF complexes share BRG1 or BRM as ATPase subunits (red) that complex with various core subunits (yellow), accessory subunits (blue) and several unique accessory subunits (green) forming BAF, PBAF and ncBAF. (B) ISWI subfamilies are comprised of SMARCA5 or SMARCA1 as the core ATPase subunits (red), and accessory proteins (blue) and unique accessory subunits (green), making up to 16 complexes. (C) Chromatin remodelers in the CHD family can be divided into 3 subfamilies. Subfamily 1 and 3 consist of the core CHD ATPase. CHD subfamily 2 reflects a larger complex of multiple accessory subunits (blue). (D) Remodelers of INO80 subfamilies have several proteins that bind to scaffold domains including an ATPase (red), core subunits (yellow) and accessory subunits (blue).
The core subunits include SMARCB1 (SNF5), SMARCE1, SMARCC2, Synovial Sarcoma Translocation Gene on Chromosome 18 (SS18), and SS18-Like 1 (SS18L1). Some accessory subunits are common in major subfamilies, such as SMARCC1, SMARCD1/2/3, β-actin, Actin Like 6A/B (ACTL6A/B), and B-Cell Lymphoma 7 Protein Family Member A/B/C (BCL7A/B/C). The remaining accessory subunits act as signature unique subunits, which define the three SWI/SNF complexes. These specific subunits include AT-Rich Interactive Domain-Containing Protein 1A/1B (ARID1A/ARID1B) and Doubled PHD Fingers Family 1/2/3 (DPF1/2/3) for BAF; ARID2, PHD Finger Protein 10 (PHF10), Polybromo 1 (PBRM1), and Bromodomain Containing 7 (BRD7) for PBAF; and BRD9, BRD4 Interacting Chromatin Remodeling Complex Associated Protein (BICRA), and BICRA-Like (BICRAL) for ncBAF (Mohrmann and Verrijzer, 2005; Mani et al., 2017). The core subunits of SWI/SNF can be combined with different specific subunits, which results in tissue- and developmental-specific remodeler subtypes (Ho and Crabtree, 2010; Clapier et al., 2017). BAF subunits bind strongly on enhancer regions, while PBAF and ncBAF subunits are mostly enriched on promoters (Nakayama et al., 2017; Gatchalian et al., 2018; Schick et al., 2019; Wang et al., 2019; Mittal and Roberts, 2020).
Role in breast cancer
3.1.2
Among the four remodeling complexes, the SWI/SNF complex is the most studied, with the strongest link in various cancers (Varela et al., 2011; Wilson and Roberts, 2011). In general, the SWI/SNF complex is thought to be a tumor suppressor (Jones et al., 2022). Around 20% of all cancers possess mutations in the genes encoding SWI/SNF subunits (Kadoch et al., 2013). The mammalian SWI/SNF complex comprises subunits that require each other for redundant/non-redundant genomic functions. Hence, any changes in the concentration of the subunits may lead to aberrant gene and transcription activity, which can eventually expedite cancer progression (Wu et al., 2017; Cui et al., 2019; Jones et al., 2022).
Contrary to the tumor suppressive role of the SWI/SNF complex, the ATPase subunits of the SWI/SNF complex, BRM and BRG1, are highly expressed in primary breast cancer compared to normal breast tissue and are vital for cell proliferation (Table 1) (Bai et al., 2013; Wu et al., 2015). BRG1 is known to play various essential roles, but it can act either as a tumor suppressor or a tumor promoter in different types of breast cancer (Wu, 2012; Wu et al., 2015; Li K. et al., 2024). As a tumor suppressor, BRG1 can interact with another tumor suppressor, BRCA1, to stimulate the transcription of TP53 or regulate TP53 directly (Bai et al., 2013; Sobczak et al., 2019; Sobczak et al., 2020). On the other hand, BRG1 can also promote cell proliferation by associating with E2F transcription factors on gene promoter sites (Li K. et al., 2024). In addition, BRG1 is required as a coactivator in ER signaling. BRG1 is recruited in an estrogen-dependent manner to enhancer DNA regions containing estrogen-responsive elements (EREs), which show ERα binding and possess active marks such as histone acetylation (DiRenzo et al., 2000). The chromatin remodeling mediated by BRG1 and the activity of histone acetyltransferases such as CBP, p300, and p300/CBP-associated factor (PCAF) on these enhancers lead to nuclear hormone receptor-dependent transcriptional activation (Sobczak et al., 2020). Furthermore, BRG1 is vital in supporting proliferation by promoting fatty acid synthesis in TNBCs (Wu et al., 2016a; Li K. et al., 2024). On the other hand, BRM loss promotes proliferation and drives the transformation of normal epithelial cells (Cohet et al., 2010). A study by Yang et al. further confirmed that the expression level of BRM is inversely correlated with breast cancer malignancy. This occurs due to the epigenetic activation of Claudins, a family of tight junction proteins, which further suppresses the migration and invasion of breast cancer cells (Yang et al., 2019). However, various knockout and knockdown studies on BRG1 and BRM further confirmed its role in maintaining cellular proliferation, in which the absence or reduced expression of BRG1 or BRM significantly decreases the proliferation rate of breast cancer cells in both in vivo and in vitro models (Bai et al., 2013; Wu et al., 2015). The effect of BRG1 or BRM perturbations on cell proliferation might be mediated through independent mechanisms, as the knockdown of each gene reduced cell viability independently, while combined knockdown resulted in an additive effect on cell proliferation.
SMARCD2, BCL7C and SS18L1 are amplified in 6%–8% of breast cancer patients from the TCGA and METABRIC breast cancer patient cohorts (Figure 4) (Curtis et al., 2012; Weinstein et al., 2013). Immunohistochemistry (IHC) analysis by Tropee et al. in breast cancer patient samples further signifies the possibility of SMARCD3’s role in tumour suppression through its association with cell cycle regulators (Tropée et al., 2021). While SMARCD2 is known to play a crucial role in regulating chromatin accessibility and is amplified in many patients (Figure 4), evidence regarding its role in breast cancer remains scarce and needs further exploration.
Summary plot of breast cancer study cohorts showing the most altered ISWI, CHD, SWI/SNF, and INO80 complex genes from the chromatin remodeling complexes in patients from METABRIC and TCGA invasive breast cancer studies from cBioPortal.
Besides the core subunits, significant aberration of the SWI/SNF complex activity may be caused by mutations in auxiliary subunits. ARID1A and ARID1B in the BAF subfamily are frequently mutated in breast cancer (Li K. et al., 2024). The majority of ARID1A mutations are heterozygous deletions, but they show biallelic inactivation of the gene due to epigenetic mechanisms, potentially leading to complete protein loss in IHC analyses in patients (Priestley et al., 2019). ARID1A and ARID1B may influence breast cancer by promoting various cellular functions related to tumor suppression, hence controlling aggressiveness and therapeutic response (Li K. et al., 2024). Moreover, approximately 58% of the ARID1A and ARID1B mutations are known to occur in their intrinsically disordered regions (IDRs) (Patil et al., 2023). These IDRs are heterogeneous conformational ensembles that engage in non-specific interaction compared to other folded domains (Konrat, 2014; Piovesan et al., 2021). Functionally, IDRs are considered to be responsible for influencing the dynamics of chromatin-bound proteins, and creating transcription hubs or phase-separated dense condensates, leading to transcriptional regulation (Boija et al., 2018; Sabari et al., 2018; Wei et al., 2020). Hence, overcoming the challenges in targeting IDRs may present a promising therapeutic strategy for treating breast cancers.
ARID1A maintains various vital cellular functions, including cell cycle control, preserving genomic integrity during double strand break repair processes, preventing telomere lengthening (Li and Lee, 2023). ARID1A is also known to participate directly in DNA repair by promoting DNA end resection (Davó-Martínez et al., 2023). ARID1A is known to perform its role by mobilizing nucleosomes to create nucleosome-depleted regions, activating the binding of transcription factors, and thus enabling gene transcription. On the other hand, ARID1A associates with nuclear hormone receptors on enhancers to control the expression of estrogen target genes and cell cycle regulators (Nagarajan et al., 2020). Other studies also demonstrate that ARID1A is critical in mediating chromatin accessibility on promoters and enhancers and thus regulates their activity (Alver et al., 2017; Mathur et al., 2017). Therefore, loss-of-function mutation in ARID1A may compromise the expression of many genes involved in tumor suppression, differentiation, Epithelial to Mesenchymal Transition (EMT), and lineage specificity, which subsequently leads to cancer progression. This is supported by an in vitro migration assay study, which found that ARID1A effectively inhibits cell migration in multiple breast cancer cell lines (Guo et al., 2018). In addition, ARID1A loss-of-function is also correlated with the presence of activating mutations in the gene of Phosphatidylinositol-4,5-Bisphosphate 3-Kinase Catalytic Subunit Alpha (PIK3CA), loss of Phosphatase and Tensin Homolog (PTEN) expression, and loss of p53 function (Yamamoto et al., 2012; Zang et al., 2012; Bosse et al., 2013; Allo et al., 2014). A clinicopathological analysis also revealed that 78% of TNBCs exhibit low ARID1A expression, and breast cancer with low ARID1A mRNA expression is associated with advanced tumors, increased p53 expression, and high Ki-67 expression (Zhang et al., 2012).
Interestingly, in metastatic, drug-relapsed or endocrine-resistant ER-positive breast cancer patients and primary lobular breast cancer patients, who are generally resistant to ER-targeted therapies, ARID1A inactivating mutations are more frequently observed, suggesting that ARID1A loss is sufficient to mediate therapy resistance (Desmedt et al., 2016; Yates et al., 2017; Razavi et al., 2018; Nagarajan et al., 2020; Xu et al., 2020; Schwartz et al., 2021). Consistently, ARID1A is known to control therapeutic response in breast cancer cell lines by influencing the innate proliferative potential, as shown by Nagarajan et al. (2020). In this study, ER+ breast cancer cells with ARID1A loss showed increased tumor growth in both untreated and 4-hydroxytamoxifen (a selective estrogen receptor modulator)-treated cells compared to ARID1A wild-type. Using Chromatin Immunoprecipitation assay with sequencing (ChIP-seq) analyses, it was observed that ARID1A binding decreases with estrogen and increases after 4-hydroxytamoxifen treatment, and its loss led to upregulation in the expression of tamoxifen-response genes, suggesting ARID1A as a corepressor and tumor suppressor. The effect on endocrine resistance can also be explained by the increased chromatin accessibility and decreased binding of histone deacetylase 1 (HDAC1) following the loss of ARID1A, which supposedly promotes the transcriptional process. The resulting hyperacetylation on histone H4K5, 8 and 12 residues (but not H3K27) due to HDAC1 loss leads to increased binding of a Bromodomain and Extra-Terminal domain (BET) containing protein, BRD4, which influences subsequent BRD4-dependent transcription (Nagarajan et al., 2020). Interestingly, a lineage-specific pioneer factor Forkhead Box A1 (FOXA1) promotes ARID1A binding on a subset of enhancers, while estrogen stimulation and its subsequent ER activation regulate ARID1A binding (Nagarajan et al., 2020). The findings from this work were concordant with a similar study by Xu et al. on ARID1A knockout in breast cancer cells deprived of estrogen, which showed increased proliferation compared to controls, suggesting a role in estrogen-independent proliferation (Xu et al., 2020). Moreover, ARID1A depletion in vitro leads to resistance towards Fulvestrant (a selective estrogen receptor degrader). This process is modulated through the reprogramming of chromatin accessibility and chromatin landscape of the breast cancer cells. In contrast to Nagarajan et al., Xu et al. identified that the loss of ARID1A induces a reduction in H3K27ac levels, which act as a strong marker for active enhancers and promoters, in several sites that have lost chromatin accessibility. Moreover, ARID1A loss disrupts the occupancy of several transcription factors, including master regulators of ER-dependent transcription and the key determinants of luminal phenotypes, such as ER, FOXA1, and GATA3, while at the same time increasing the binding of TEA Domain Transcription Factor 4 (TEAD4), which is more enriched in the basal phenotype. TEAD4 silencing is associated with the sensitivity of ARID1A knockout cells to fulvestrant. Furthermore, gene set enrichment analysis (GSEA) by Xu et al. upon ARID1A knockout reported a significant activation of basal-like transcription, as reflected by the increased activity of stemness-related gene expression such as keratins KRT6, KRT15, KRT5, CD44, and Tumor Protein P63 (TP63). Altogether, the findings from Xu et al., indicated a cellular plasticity of luminal to basal transition upon ARID1A loss, making the cells resistant to endocrine therapies.
ARID1B is the homologous counterpart of ARID1A with similar function. Hence, *ARID1A-*mutated tumours may depend on ARID1B for proliferation, suggesting a synthetic lethality combination between ARID1A and ARID1B and thus providing an opportunity for personalised targeting of cancer growth (Nagl et al., 2007; Helming et al., 2014). Given its function in regulating the cell cycle, the above studies showed that ARID1B is an essential subunit of the SWI/SNF complex, with a broad role in promoting proliferation and cell cycle progression. Various evidence illustrates that inactivating mutations in ARID1B enable the clonal selective advantage in cancer cells, especially in breast cancer with drug relapse (Stephens et al., 2012). ARID1B also acts as a prognostic biomarker whose expression is associated with disease-free survival in breast cancer patients (Cui et al., 2019; Li K. et al., 2024). On the other hand, ARID1B is highly expressed in TNBC compared to other molecular types of breast cancer. Its high expression is closely related to the clinicopathological profile of the patients, such as age, tumor size, histological grade, and nuclear polymorphism, and serves as an independent predictive marker (Shao et al., 2015).
Similar to the BAF subfamily, the mutations in auxiliary subunits in the PBAF subfamily, such as PBRM1 and ARID2, are frequently observed in breast cancers (Li K. et al., 2024). PBRM1 is a tumor suppressor gene that binds to the p21 promoter and upregulates its baseline and signal-dependent transcription, subsequently inhibiting tumor development in breast cancer cells (Xia et al., 2008). PBRM1 interaction with p21 protein is also implicated in various functions, including DNA damage response, cell cycle regulation, cellular differentiation, and maintenance of genomic stability (Hopson and Thompson, 2017; Hagiwara et al., 2021; Yamashita et al., 2023). However, PBRM1 is known to promote resistance to T-cell-dependent killing in preclinical cancer models, hence the possibility of affecting the anti-tumor immune response (Miao et al., 2018; Pan et al., 2018). Several studies identified PBRM1 as a prognostic indicator in breast cancer patients. PBRM1 expression is strongly correlated with clinical stage and lymph node status and serves as a significant indicator for overall survival and recurrence-free survival in breast cancer patients. Low expression of PBRM1 in breast cancer tissues is associated with an unfavorable outcome (Mo et al., 2015).
Despite sharing the same AT-rich interaction domain and some structural similarities, ARID2 is not a homolog of BAF subunits ARID1A and ARID1B (Li K. et al., 2024). A decrease in ARID2 expression is frequently found in non-luminal breast cancer subtypes but ARID2 is a predictor of poor survival in ER-positive breast cancer patients (Zhang et al., 2021). Furthermore, a genomic analysis in matched primary and metastatic breast cancer tissues by Yates et al. found that ARID1A, ARID1B, and ARID2 are often inactivated in recurrent breast cancer. This finding is of significant interest since these genes are commonly wild-type in primary breast cancer and imply selective treatment-induced clonal evolution of aggressive cells with these mutations (Yates et al., 2017). Further investigation on the role of ARID2 in breast cancer would be of great interest.
While SWI/SNF is overall considered a tumor suppressor, some members of the subfamily possess dual roles as tumor suppressors and oncogenes in certain cancers (Table 1) (Sen et al., 2019; Fontana et al., 2023), but the mechanism underlying this context-specific function is unknown.
ISWI subfamily
3.2
ISWI subfamily is closely related to the SWI/SNF and is mainly responsible for nucleosome assembly and organization following DNA replication and transcription at locations where the nucleosomes are ejected (Clapier et al., 2017). Upon chromatin disassembly (e.g., in the replication process), histone H3-H4 tetramers and H2A-H2B dimers were brought to adjacent DNA by histone chaperones. ISWI remodelers then assist in forming prenucleosomes with DNA-histone combinations and their maturation into nucleosomes with DNA-histone octamers (Torigoe et al., 2011; Gurard-Levin et al., 2014; Fei et al., 2015; Clapier et al., 2017). Subsequently, by placing them at a fixed and appropriate distance, the ISWI remodeling complex promotes the formation of nucleosomes arranged on beads-on-a-string structures (Ito et al., 1997; Varga-Weisz et al., 1997; Corona et al., 1999; Clapier et al., 2017).
Structure and composition
3.2.1
Structurally, the ATPase domain of the ISWI subfamily is separated by a small insertion sequence and flanked by two domains, the autoinhibitory N-terminal (AutoN) and the negative regulator of coupling (NegC), which function as the regulators of ATPase activity (Figure 1B) (Corona and Tamkun, 2004; Yadon and Tsukiyama, 2011; Clapier and Cairns, 2012). The C-terminal HSS domain of ISWI remodelers binds to the unmodified histone H3 tail and the linker DNA, flanking the nucleosome (Grüne et al., 2003; Boyer et al., 2004; Dang and Bartholomew, 2007; Clapier et al., 2017).
ATP-utilising chromatin assembly and remodeling factor (ACF) complex catalyzes the relaxation of the chromatin, aiding histone deposition, and the chromatin accessibility complex (CHRAC) allows nucleosome sliding and assembly on the regions with nonhomologous end-joining repair of double-strand breaks (Kukimoto et al., 2004; Lan et al., 2010). While these subfamilies regulate chromatin accessibility and thus repress transcription, an ISWI subset named the NURF complex may, in contrast, promote transcription (Xiao et al., 2001; Hochheimer et al., 2002; Clapier and Cairns, 2009). These dual roles may be associated with their differences in the activity of mechanistic sliding. Hence, the ISWI subfamily may act either as an activator or repressor during transcription, depending on the availability of other factors (Narlikar et al., 2002; Erdel et al., 2010).
In total, there are 16 ISWI subfamily complexes identified in mammals. Each complex of the ISWI subfamily is composed of one ATPase subunit, either SMARCA5 or SMARCA1, and other noncatalytic subunit(s) (Figures 2, 3B). For instance, the combination of the ATPase subunit with Bromodomain Adjacent to Zinc Finger Domain protein BAZ2B, will constitute either BRF1 (B-related factor 1) or BRF5 remodeling complexes, depending on whether the ATPase involved is SMARCA1 or SMARCA5, respectively. An exception is the NURF1 complex, which is composed of SMARCA1 with BPTF (Bromodomain PHD finger Transcription Factor) and additional RBBP4 and RBBP7 subunits, compared to the NURF5 (NUcleosome Remodeling Factor) counterpart, which only consists of the combination of SMARCA5 and BPTF.
Role in breast cancer
3.2.2
ISWI remodeling complexes have multifaceted functions as they are known to promote the transcription of oncogenes (Table 1), but some of their members are also implicated in tumor suppression. Genetic alterations of ISWI members are commonly reported in many types of cancers, and they also correlate with poor prognosis of breast cancer patients (Pérez-Pena et al., 2019; Li et al., 2021), as ISWI ATPases interact with numerous DNA-binding factors and cofactors involved in malignant cell transformation and progression (Mayes et al., 2017; Koedoot et al., 2019).
Previous studies have shown that a high level of BAZ1A in HER2+ breast tumors is associated with poor overall survival and relapse-free survival, and amplification of Bromodomain PHD Finger Transcription Factor (BPTF) in breast tumors is linked to short metastasis-free survival (Pérez-Pena et al., 2019; Li et al., 2021). Overexpression of SMARCA5 is also frequently seen in breast cancer, which is positively correlated with the stages of tumor, node and metastasis, and poor overall survival (Jin et al., 2015). SMARCA1 plays a vital role in maintaining cell survival and cell cycle progression, as inhibition of SMARCA1 leads to the upregulation of Apoptotic Protease Activating Factor 1 (APAF1) and thus increased activity of caspase 9 in primary breast cancers (Ye et al., 2009; Li et al., 2021). SMARCA5 may be responsible for the transition from G1 to S phase in cell proliferation and for accommodating the Matrix Metallopeptidase 2 (MMP2)-mediated invasion (Jin et al., 2015). However, according to a study by Dai et al., SMARCA1 downregulation is associated with poorer prognosis in breast cancer patients, which warrants further investigation into its role in breast cancer prognosis (Ye et al., 2009; Dai et al., 2022). Gene expression analysis by Li et al. showed that SMARCA1 is downregulated while SMARCA5 is upregulated in invasive breast carcinoma, suggesting opposing roles of these proteins (Li et al., 2021). However, other studies demonstrated increased expression of both proteins in the same context (Jin et al., 2015). Hence, a further study confirming their expression levels and functions in various subtypes of breast cancer is needed, as the balanced expression of SMARCA5 and SMARCA1 may be of interest for therapeutic purposes.
Another member of the ISWI subfamily implicated in breast cancer progression is the Remodeling and Spacing Factor (RSF) complex, which consists of a chaperone nuclear protein and SMARCA5 or SMARCA1. The overexpression and amplification of RSF1 are reported in many cancers, including breast cancer (Figure 4), and are associated with poor overall survival, advanced clinical features, and drug resistance (Li et al., 2021). However, RSF1’s role in malignant transformation is associated with p53 expression. In normal cells, RSF1 upregulation induces Ataxia Telangiectasia Mutant (ATM)/p53-dependent DNA damage response, eventually leading to growth arrest and apoptosis. Thus, mutation or inactivation of p53 compromises the growth-inhibitory effects of RSF1 and is paired with the overexpression of SMARCA5, eventually promoting cell proliferation (Sheu et al., 2010; Kshirsagar et al., 2012). This mechanism might provide a selective advantage for p53-mutated cells, allowing RSF1 to act as a driver gene in this context (Sehdev et al., 2010; Sheu et al., 2013; Li et al., 2021).
The components of ACF and CHRAC complexes, including BAZ1A, CHRAC1, and POLE3, are also upregulated in breast cancers (Li et al., 2021). However, BAZ1A is associated with cellular senescence by regulating Mothers Against Decapentaplegic homolog 3 (SMAD3) and p21 in tumor cells (Ito et al., 1999; Li et al., 2019). As for CHRAC complexes, the CHRAC1 gene is amplified on chromosome 8q24.3 and it is confirmed to be a driver gene that promotes the proliferation and clonal survival of breast cancer cells (Mahmood et al., 2014). Like its homologous counterpart, BAZ1B is overexpressed and amplified in breast cancer (Figure 4). It promotes the expression of the Cytochrome P450 Family 19 Subfamily A Member 1 (CYP19A1) gene, which encodes the aromatase enzyme that produce estrogens from androgens (Lundqvist et al., 2018; Li et al., 2021). Aromatase inhibitors, such as Letrozole, Anastrozole, and Exemestane, reversibly bind to aromatase to prevent androgens from binding (Ghodsi and Hemmateenejad, 2016; Bhatia and Thareja, 2024). However, the upregulation of BAZ1B may lead to the deregulated expression of CYP19A1 and ER-α coding gene, ESR1, reducing the efficacy of aromatase inhibitors. Interestingly, vitamin D analogs can be introduced to dissociate BAZ1B from the CYP19A1 promoter and to improve the efficacy of aromatase inhibitors. This instance highlights the feasibility of targeting BAZ1B indirectly via vitamin D analog modulation (Lundqvist et al., 2013; Lundqvist et al., 2018).
Another documented ISWI subfamily member implicated in breast cancer progression is the NURF complex with BPTF and RBBP4/7 subunits. BPTF is known to be frequently amplified in breast cancer (Figure 4) (Li et al., 2021). High BPTF copy number is significantly associated with advanced tumor grade in ER+ and TN breast cancers. BPTF expression may also be responsible for promoting proliferation and controlling apoptosis, as evidenced in an in vivo TNBC study (Bezrookove et al., 2022). According to the study by Li et al., the expression levels of several ISWI subfamily members, including NURF complex, are strongly associated with immune checkpoint activation and/or tumor-infiltrating immune cell ratio. In breast cancer, the BPTF/NURF complex may promote the immune escape of cancer cells (Landry et al., 2011). Knockout of BPTF in mouse breast cancer models promotes the expression of immunoproteasome subunits Psmb8 and Psmb9 and antigen transporters Tap1 and Tap2, therefore resulting in enhanced antigenicity and T-cell cytotoxicity. Additionally, BPTF-deficient tumor cells may exhibit increased perforin, granzyme, and IFN-g due to enhanced T-cell cytotoxicity, which may induce Janus Kinase/Signal Transducers and Activators of Transcription (JAK/STAT) and Fas Cell Surface Death Receptor/Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (Fas/TRAIL) pathways (Mayes et al., 2016; Mayes et al., 2017). In breast cancer cell lines, BPTF can also inhibit the antitumor activity of NK cells by reducing the cell surface heparan sulfate proteoglycan and natural cytotoxicity receptor co-ligand abundance (Mayes et al., 2017).
Due to the involvement of RBBP4/7 in several multi-protein transcription complexes other than ISWI, there has been no exact conclusion as to whether RBBP4 and RBBP7 execute these functions in an ISWI or non-ISWI-dependent manner in breast cancer (Abbey et al., 2018; Moody et al., 2018; Glancy et al., 2021). However, in other cancers, such as gastric and liver, RBBP4 is recruited as a part of the NURF complex to promote transcription of certain factors [e.g., SRY-Box Transcription Factor 2 (SOX2) and Octamer-Binding Transcription Factor 4 (OCT4)] that drive tumor progression (Zhu et al., 2015; Ding et al., 2019; Cai et al., 2023). In breast cancer, RBBP4/7 are known to function as part of HDAC complexes. RBBP7 is documented for its role in metastasis by binding to EMT-related genes in several cancers (Li et al., 2010; Li et al., 2016; Wang J. et al., 2020; Li et al., 2021). Interestingly, the RBBP4-containing complex is also involved in drug resistance (Li et al., 2021). In TNBC, B-cell CLL/lymphoma-11A (BCL11A) is a transcription factor that contributes to maintaining the chemo-resistant breast cancer stem cell population through its interaction with RBBP4. Inhibition of RBBP4-BCL11A complex formation by BCL11A peptide inhibitor may decrease aldehyde dehydrogenase-positive breast cancer stem cells; hence, targeting interactions between RBBP4 and oncogenic transcription factors may provide opportunities for intervention (Moody et al., 2018).
Additionally, another ISWI family member, BRF1/5 complexes, are formed by the interaction of BAZ2B with SMARCA1/5 [!!! INVALID CITATION !!! (Oppikofer et al., 2017)]. BAZ2B is reported to be downregulated in primary breast cancer, but its role in cancer progression is yet to be elucidated (Li et al., 2021). CECR2, which is a subunit of the CECR2-containing Remodeling Factor (CERF) complex, is also implicated in breast cancer metastasis (Zhang M. et al., 2022). Upregulation of CECR2 in breast cancer metastasis is also attributed to the modulation of tumor immunity by promoting M2 macrophage polarization to create an immunosuppressive environment. This is achieved by forming a complex with p65 through its bromodomain, which can subsequently activate the expression of NF-kB target genes such as Colony Stimulating Factor-1 (CSF1) and C-X-C Motif Chemokine Ligand 1 (CXCL1) that mediate macrophage-mediated immune suppression upon metastasis (Zhang M. et al., 2022).
CHD subfamily
3.3
The remodelers of the CHD subfamily are structurally similar to those of the ISWI subfamily, except that they have the tandem chromodomains. CHD remodeling factors differ in function depending on the chromodomain diversity and perform all three general remodeling processes (Lusser et al., 2005; Denslow and Wade, 2007; Konev et al., 2007; Clapier and Cairns, 2009; Kunert and Brehm, 2009; Murawska and Brehm, 2011). However, they are mainly implicated in nucleosome assembly and repression of transcription primarily through involvement as part of the Mi-2/nucleosome remodeling and deacetylase (NuRD) complexes, which contain histone deacetylases (HDAC1/2) and methyl CpG-binding domain (MBD) proteins (Figure 2) (Denslow and Wade, 2007; Clapier and Cairns, 2009; Allen et al., 2013). Predominantly, members of the CHD subfamily are associated with tumor suppression, however, some members, such as CHD1 and 8, are reported to exhibit both tumor-suppressive and pro-oncogenic functions depending on the context (Tan et al., 2014; Yu et al., 2015).
Structure and composition
3.3.1
CHD family proteins are structurally similar to those of the ISWI family, except for the tandem chromodomains in the amino terminus and they only possess the NegC regulator, followed by DNA-binding domain (DBD) with only the SANT and SLIDE domain instead of the HSS domain (Figure 1B) (Tran et al., 2000; Kunert and Brehm, 2009; Hauk et al., 2010; Ryan et al., 2011). CHD proteins are further divided into three subfamilies, depending on the presence of their unique domains (Figure 3C) (Mills, 2017). For example, Subfamily I (CHD1, CHD2) possess SNF2 domains homologous to CHD1 proteins of other organisms (such as mouse Chd, which has AT-rich DNA binding domain), Subfamily II (CHD3, CHD4, CHD5) possess dual plant homeodomains (PHDs) which are known to bind to histone methylation, and Subfamily III (CHD6, CHD7, CHD8, CHD9) has domains such as Brahma and Kismet (BRK), PHD Zinc-finger like, SANT-like, CR, and DNA-binding domains (Delmas et al., 1993; Marfella and Imbalzano, 2007; Mills, 2017).
Role in breast cancer
3.3.2
Since the CHD proteins control fundamental biological processes by altering chromatin compaction and regulating access to DNA, they are crucial in regulating cancer progression (Table 1). Their genes are heavily altered in around 30 types of cancers (Dahiya et al., 2019). For example, CHD subfamily I affects cancer invasion, metastasis and overall survival, while subfamily II plays a central role in cancer progression by promoting EMT and metastasis (Table 1). Although subfamily III is not strongly associated with malignancy, its deregulation is reported to modulate cancer-related pathways and survival through its upstream factors (Mills, 2017; Dahiya et al., 2019).
CHD1 is a member of the CHD subfamily I that is most associated with breast cancer pathogenesis (Mills, 2017). Estrogen inhibits the expression of microRNAs that target and degrade CHD1 transcription, thereby promoting cancer cell proliferation (Tan et al., 2014; Mills, 2017). CHD1 also increases the proliferative effect of estrogen via interaction with c-MYC However, inactivating mutations in CHD1 are observed in breast cancers (Curtis et al., 2012; Weinstein et al., 2013). CHD2 has been implicated in tumor suppression, although this correlation still needs further exploration. Russo et al. proposed that CHD2 transcription induced by human chorionic gonadotropin during pregnancy may prevent breast cancer (Russo and Russo, 2012; Mills, 2017). The role of CHD2 in breast cancer progression might be more apparent in the presence of p53 heterozygosity, such as in Li-Fraumeni syndrome. This is based on the crucial function of Chd2 in maintaining development, as Chd2 is a candidate gene target of breast cancer genetic susceptibility gene Mtsm1 (Marfella et al., 2006; Koch et al., 2007; Mills, 2017).
CHD subfamily II acts as a modulator of cellular proliferation and cell cycle progression (Bagchi et al., 2007; Mills, 2017) and is implicated in various processes in breast cancers, including chemoresistance, EMT, metastasis, etc. (Wang et al., 2011; Wu et al., 2012; Mills, 2017). CHD5 is the most well-studied among the subfamily members for its role in breast cancer. CHD5 loss-of-function or inactivation due to promoter hypermethylation, deletions and/or mutations has been reported in breast cancer pathogenesis (Bagchi et al., 2007; Mulero-Navarro and Esteller, 2008; Wu et al., 2012). In mouse models, its tumor-suppressor function is achieved by activating the cell cycle inhibitor gene, Cyclin Dependent Kinase Inhibitor 2A (Cdkn2a), which encodes the two proteins p16Ink4a and p19Arf (Bagchi and Mills, 2008). Moreover, CHD5 represses the expression of WEE1, which is a mitotic checkpoint gene (Quan et al., 2014). Furthermore, Chd5, as part of the Trithorax complex, modulates the activity of the polycomb repressive group complex (PcG) and inhibits the expression of the oncoprotein subunit Bmi (Paul et al., 2013; Xie et al., 2015). CHD5, as a component of the NURD complex, also works in parallel with other NURD complex components containing histone deacetylase activity, which may further explain its role in tumor suppression (Quan et al., 2014; Quan and Yusufzai, 2014; Kolla et al., 2015). In mouse models, Chd5 functions functions as a dose-dependent tumor suppressor, as its heterozygous loss induces immortalization and tumorigenesis, while having three copies leads to senescence, apoptosis, and perinatal lethality (Bagchi et al., 2007; Mills, 2017). Overall, these findings suggest that CHD5 expression might correlate directly with breast cancer survival. (Wu et al., 2012).
Akin to CHD5, CHD3 and CHD4 are components of the NURD complex, whose roles are implicated in transcription, proliferation, and DNA damage repair (O’Shaughnessy and Hendrich, 2013; Mills, 2017). While CHD3 is considered as an oncogene and CHD4 a tumor suppressor, recent evidence demonstrates the role of CHD4 as an oncogene in breast cancers, as its amplifications are more common (Figure 4). CHD4 regulates downstream pathways for cellular proliferation and aggressiveness through transcriptional mechanisms while maintaining genomic stability through non-transcriptional pathways (Hou et al., 2017; Luo et al., 2018; Ou-Yang et al., 2019; Wang Y. et al., 2020). Silencing CHD4 causes an arrest in the G0 phase, significantly reducing DNA synthesis and upregulation of p21^WAF1,^ which is a cell cycle inhibitor (D'Alesio et al., 2016). Cells deficient in CHD4 show an increase in p21 expression in BRCA-proficient cell lines via inhibition of HDAC1 recruitment on the p21 promoter (Hou et al., 2017). Depletion of CHD4 significantly decreases cell proliferation and migration in HER2-positive and triple-negative breast cancer cell lines, leading to a reduction in tumor mass in luminal B and HER2-ortholog-activated triple-negative PDX and transgenic mouse models (D'Alesio et al., 2016; D'Alesio et al., 2019). The regulation of cell proliferation in HER2-positive breast cancer cell line with CHD4 depletion is explained by the observed increase in HER2 Tyr-1248 phosphorylation and subsequent inhibition of the downstream HER2/PI3K/AKT/ERK signaling cascade. CHD4 positively correlates with cell motility and mortality, and its silencing triggers hyperacetylation of histone H3 on the E-cadherin promoter, further reducing migration and invasiveness in TNBC and non-TNBC cell lines (Luo et al., 2018). Furthermore, CHD4 regulates these aggressive properties through direct control on the expression of β1-integrin and p21 in TNBCs (Hou et al., 2017; Luo et al., 2018; Ou-Yang et al., 2019). Upon knockdown, reduction in the expression of EMT markers such as Vimentin, β-catenin and SNAI1 was observed (Ou-Yang et al., 2019).
Moreover, CHD4 loss may promote dysregulation in autophagy, which is one of the underlying causes of cancer (Wang et al., 2013; Wei et al., 2015; D'Alesio et al., 2019). Autophagy is marked by changes in microtubule-associated protein 1A/1B-light chain 3 (LC3) levels, a protein involved in autophagosomal membrane formation and degradation (Kabeya et al., 2000; Kabeya et al., 2004). The ratio of cytosolic LC3-I level and membrane-bound LC3-II level can be used to estimate autophagic activity (Tanida et al., 2008). During the process of autophagy, an autophagy adaptor protein, p62, binds to LC3 on the autophagosome and to ubiquitinated proteins, promoting subsequent degradation (Lippai and Lőw, 2014; Liu et al., 2016). CHD4 depletion causes accumulation of p62 and an increase in the LC3 II/I ratio, suggesting a block in late-stage autophagy in HER2-positive breast cancer cells, further contributing to growth arrest in cancer cells upon CHD4 loss (D’Alesio et al., 2017; D'Alesio et al., 2019).
Even though CHD subfamily III is shown to be dysregulated in cancer, it is more implicated in neurological and developmental disorders (Ronan et al., 2013). CHD8 is a member of subfamily III with the strongest link to breast cancer. CHD8 mediates estrogen and cyclin D1-mediated recruitment of E2F1 to the promoter of cyclin E2, facilitating the proliferative effect of estrogen (Caldon et al., 2009). However, CHD8 is one of the top tumor suppressor genes, mutated in breast cancer (Figure 4) (Pongor et al., 2015). Interestingly, CHD8 is known to influence progestin-dependent gene regulation, and it also interacts with the SWI/SNF complex, where depletion of its ATPase subunits inhibits CHD8 recruitment (Ceballos-Chávez et al., 2015). CHD7 is the most amplified CHD protein, represented in around 11% breast cancer patients (Figure 4) and its amplifications are more prevalent in aggressive breast cancer subtypes, correlating with high tumor grade and poor prognosis. This association may support the findings from CHD7 knockdown models in triple-negative breast cancer cell lines showing inhibition of cell proliferation and decreased gene expression of several CHD7 targets, including the NRAS oncogene (Chu et al., 2017). CHD3 and CHD9 are the most deleted CHD genes in breast cancer (Figure 4), with 60% and 55% of breast cancer patients showing heterozygous loss, respectively (Chu et al., 2017). However, their role in cancer progression is still obscure to date.
INO80 subfamily
3.4
The most prominent function of the INO80 subfamily lies in histone editing, in which a particular histone in a nucleosome is removed in a replication-independent manner and replaced with either a canonical or variant histone (Figure 1) (Clapier et al., 2017). INO80 complex regulates chromatin structure through mobilizing mononucleosomes in an ATP-dependent manner in two different ways (Bao and Shen, 2007; Chen et al., 2013). The first is by catalyzing the sliding of nucleosomes, while the second is by catalyzing the replacement of histone H2A-H2B dimers with H2AZ-H2B dimers in nucleosomes (Shen et al., 2000; Jin et al., 2005; Papamichos-Chronakis et al., 2011; Chen et al., 2013). While H2AZ is associated with active transcription due to its role in the recruitment of transcription factors, especially on enhancers, its removal from promoters is also required for gene activation (Santisteban et al., 2000; Dhillon et al., 2006; Brahma et al., 2017). It is also implicated in homologous recombination following DNA damage and the prevention of non-coding transcription (van Attikum et al., 2004; Conaway and Conaway, 2009; Xue et al., 2015; Brahma et al., 2017). The functions of two other complexes, however, are slightly different than the INO80 complex, in which the SRCAP complex replaces canonical H2A-H2B dimers with the H2A.Z variant similar to the INO80 complex, while the p400/TIP60 complex replaces H3.1 histone with variant H3.3 (Mizuguchi et al., 2004; Papamichos-Chronakis et al., 2011; Pradhan et al., 2016; Clapier et al., 2017).
Structure and composition
3.4.1
One prominent feature of INO80 subfamily is that these proteins possess a long insertion of more than 1,000 amino acids that splits the ATPase domain, which binds a hetero-hexameric ring of helicase-related (AAA-ATPase) ruvB-like proteins (Rvb1/TIP49 (TATA-binding-protein interacting protein 49) and Rvb2/TIP48 in humans). Another property shared by the members of this subfamily is the presence of the HSA domain which recruits actin and ARP (Figures 1B, 2, 3D) (Szerlong et al., 2008; Clapier et al., 2017). The three-member complexes of this subfamily are the INO80, SNF2-Related CREB-Binding Protein (CBP) Activator Protein (SRCAP) complex, and the p400/TIP60 complexes (Figure 3D). Each subfamily complex has integral protein(s) that act as the scaffold for other subunits to interact with (Mayes et al., 2014; Dijkwel and Tremethick, 2022). The INO80 complex scaffold is structurally divided into three modules that each interact with a domain of the primary INO80 scaffold (Chen L. et al., 2011). This includes one N-terminal interacting module consisting of nuclear factors related to kB (NFRkB), Amida, Microspherule protein 1 (MCRS1**), Ubiquitin C-Terminal Hydrolase (UCH37), INO80D, and INO80E; the HSA-interacting module that is composed of actin-related proteins ARP4, ARP8, and Kruppel family zinc finger transcription factor Yin Yang 1 (YY1)**; and SNF2-ATPase interacting module which contains TIP49a, TIP49b, Inositol-eighty subunit 2 (IES2) (Chen et al., 2013). The SRCAP complex is composed of SRCAP as the primary scaffold that interacts with two modules, the ARP module and the motor module. The motor module of the SRCAP complex consists of YL1, ACTin Related protein 6 (ACTR6), TIP49a/b hexamers, and Zinc finger HIT-type containing 1 (ZnHIT), while the regulatory ARP module comprises b-actin, BAF53a, DNA Methyltransferase 1 Associated Protein 1 (DMAP1), and Glioma Amplified Sequence 41 (GAS41) (Krogan et al., 2003; Kobor et al., 2004; Mizuguchi et al., 2004).
The ARP module is shared between the SRCAP and TIP60/p400 complex, while the motor module of the TIP60/p400 complex possesses only YL1 and TIP49a/b hexamers as its subunit (Jacquet et al., 2016; Feng et al., 2018). The TIP60/p400 complex has Malignant Brain Tumor Domain-containing protein 1 (MBTD1) as an additional subunit for its ARP module and three other modules, including the Trimer Independent of Nucleosomal Acetyltransferase of H4 (NuA4) for Transcription Interactions with Nucleosomes (TINTIN) module for histone marker reading function, histone acetyltransferase (HAT) module for catalytic function, and Transformation/Transcription Domain Associated Protein (TRRAP) module for transcription activator binding function. The TINTIN module consists of several subunits, such as MRG domain Binding Protein (MRGBP), Mortality Factor 4 Related Gene on chromosomes 15 (MRG15), and BRD8. In contrast, the Histone acetyltransferase (HAT) module is composed of Enhancer of Polycomb Homolog 1 (EPC1), TIP60, INhibitor of Growth family member 3 (ING3), and Esa1-Associated Factor 6 (EAF6) (Jacquet et al., 2016). The TRRAP module is composed of only TRRAP as a large, single subunit that is tethered to the SANT domain of P400 (Auger et al., 2008).
Role in breast cancer
3.4.2
Although there are very few documented studies on the INO80 subfamily in the context of breast cancer, there has been a growing interest in researching this chromatin remodeling complex due to its unique function in editing nucleosome composition and histone specialization, which changes nucleosome stability (Clapier, 2021).
INO80 complex aberrations are associated with cancer progression, as their binding to enhancers is crucial in mediating oncogenic gene expression (Min et al., 2013; Wang et al., 2014; Runge et al., 2018; Thang et al., 2023). In the TCGA breast cancer atlas cohort, the subunits of the INO80 complex are frequently amplified, accounting for alterations present in around 5% of overall breast cancer cases (Figure 4) (Thang et al., 2023). However, when characterized under PAM50-based subtyping, INO80 expression is significantly downregulated in basal-type breast cancer (Thang et al., 2023) Importantly, INO80 functions as a mediator of dynamic replacement of an active enhancer-based histone variant, H2A.Z, and thus as a critical regulator of enhancers close to E2 target genes such as GREB1 and TFF1, contributing to breast cancer progression (García-Pedrero et al., 2006; Papamichos-Chronakis et al., 2011; Segala et al., 2016; Thang et al., 2023). Thang et al. further explained the relationship between INO80 and ER through Weighted Gene Co-expression Network Analysis (WGCNA), which indicated the existence of networks between INO80 and a subset of luminal breast cancer biomarkers, including ESR1, FOXA1, and other ER target genes such as GATA3, TFF1, and AR (Tozlu et al., 2006; Chen Y. et al., 2011; Hurtado et al., 2011; Thang et al., 2023). However, lower expression of INO80 is associated with reduced overall survival rate, distant metastasis-free survival, and recurrence-free survival in breast cancer patients (Thang et al., 2023). Low INO80 copy number status is also associated with an increased risk in TNBC patients compared to luminal and HER2+ patients. While INO80 affects the ductal morphogenesis and differentiation of the mammary gland, knockout of INO80 alone is not enough to cause cancerous changes in mouse mammary gland models through mechanisms yet to be understood.
Ubiquitin Carboxyl-terminal Hydrolase 35 (UCH37), encoded by UCHL5, is heavily amplified (Figure 4) and is associated with poor outcomes in breast cancer patients (Curtis et al., 2012; Weinstein et al., 2013). UCH37 is a deubiquitinase oncoprotein in the INO80 chromatin remodeling complex (VanderLinden et al., 2015). This protein can activate deubiquitination of E2 promoter binding factor F1 (E2F1), which subsequently enhances the transcription and increases the proliferative activity of E2F1 target genes (Mahanic et al., 2015). Moreover, UCHL5 amplification is associated with higher activity of the TGF-β signaling pathway through the formation of Smad ubiquitination regulatory factor 2 (Smurf2) – Smad7 complex. This affects the expression of subsequent TGF-β downstream targets such as MMP-2 and PAI-1 that are crucial in regulating tumor migration and invasion (Wicks et al., 2005; Wicks et al., 2006; Cutts et al., 2011).
Besides INO80, other subfamily members, including SRCAP and p400/Tip60 complexes, also regulate H2A.Z deposition into chromatin (Ruhl et al., 2006; Svotelis et al., 2010). Moreover, the p400/Tip60 complex is known to indirectly affect cell proliferation by regulating the expression of several transcription factors, such as E2F1, p53, KAI1, and Myc (Taubert et al., 2004; Kim et al., 2005; Samuelson et al., 2005; Nagl et al., 2007; Svotelis et al., 2010). Another study by Cao et al. on a long non-coding RNA, LINC00665 in breast cancer cell lines shows that it can inhibit miRNA- 641, which affects SRCAP translation by binding to its 3′UTR region. This loss of SRCAP inhibition results in proliferation and invasion of breast cancer cells, suggesting its role as an oncogene (Cao et al., 2022). This suggests that the regulation of p400/Tip60 and SRCAP complexes is related to breast cancer progression, but their exact function needs further exploration. YL1, encoded by the vacuolar protein sorting 72 homologs (VPS72) gene, is a subunit shared by the SRCAP and TIP60/p400 complex and is frequently amplified in many cancers, including breast (Figure 4) and melanoma. Given the prevalent alteration in breast cancer, this prompts further research on the possibility of utilizing VPS72 expression as a prognostic indicator in breast cancer.
Potential therapeutic approaches
4
Targeting SWI/SNF family members
4.1
As described in this review, chromatin remodelers are vital to several essential biological pathways and, therefore, pose a challenge to treat without off-target effects. Due to their significant involvement in breast cancers, their subunits can act as attractive therapeutic targets, however, they are poorly investigated in breast cancers and their association with other chromatin-associated complexes may complicate the effects. Several efforts have been made to exploit the vulnerabilities of each chromatin remodeling subfamily in other cancers, which are explained below as potential therapeutic approaches to target breast cancers. Strategies targeting these complexes can generally be divided into two categories: direct and indirect targeting (Figures 5A,B) (Zhang and Li, 2022).
Targeting SWI/SNF complex using direct and indirect strategies. (A) Inhibitors and degraders (PROTACs) which were developed to target SWI/SNF function. gen–generation. (B) Synthetic lethality interactions with SWI/SNF complex subfamilies. Blue double-headed arrows represent synthetic lethality interactions within the complex subfamilies. Red double-headed arrows represent synthetic lethality interactions with other complexes or pathways.
Direct targeting approaches
4.1.1
Targeted therapies can be directed against the exact aberrant chromatin remodelers that are altered in various types of cancer (Zhang and Li, 2022). These may work by inhibiting the ATPase catalytic subunit of the remodeling complex (for example, competitive ATP inhibitors or allosteric agents) or by disrupting the protein-protein interactions of the catalytic subunits. Presence of ATPase domains in the chromatin remodeling proteins is therapeutically beneficial due to their druggable enzymatic pockets, offering highly selective inhibitors.
For example, the ATPase activity of SWI/SNF members BRG1 and BRM can be blocked by dual allosteric small-molecule inhibitors, BRM014 or orally bioavailable inhibitor FHD-286 (Figure 5A) in BRG1-deficient lung cancer, acute myeloid leukaemia and uveal melanoma models (Papillon et al., 2018; Rago et al., 2020; Vaswani et al., 2025). Additionally, inhibitors of the SWI/SNF bromodomain, like PFI-3, have little effect as a single agent, but they can sensitise cancer cells to DNA damage by doxorubicin (Lee et al., 2021). Furthermore, Active DNA-dependent ATPase A Domain inhibitor (ADAADi) was the first SWI/SNF catalytic activity inhibitor, discovered from the byproduct of the activity of aminoglycoside phosphotransferases (Figure 5A). This exerts its function as a competitive inhibitor against DNA-dependent ATPase domains of SWI/SNF (Muthuswami et al., 2000; Dutta et al., 2012). However, its specificity towards SWI/SNF ATPase is unclear, and although it was able to sensitize breast cancer cells to chemotherapy agents, variable responses were observed in different cell lines (Rakesh et al., 2021; Dreier et al., 2024).
Importantly, subunits other than the catalytic and bromodomains of the chromatin remodeling complex, especially the SWI/SNF complex, possess virtually flat functional interfaces, due to the lack of hydrophobic ligand binding pockets, hence exhibiting a weak possibility for ligand interaction (Sammak and Zinzalla, 2015; Dang et al., 2017). This makes them undruggable with conventional approaches (Ye et al., 2019). Hence, a targeted protein degradation strategy has been pursued recently to answer this problem. This consists of a proteolysis-targeting chimera (PROTAC) with an ubiquitin-proteasome system (UPS) that precisely targets a protein by bringing an E3 ubiquitin ligase to the protein of interest and induces its ubiquitylation and degradation (Schneider et al., 2021; Békés et al., 2022). Some examples of targeted protein degraders are BRD9-directed degrader (dBRD9); VZ185, which targets BRD9 and BRD7; AU15330, AU24118, and ACBI1, which degrade SMARCA4, SMARCA2 and PBRM1 and A947, as a SMARCA2-selective degrader (Figure 5A) (Remillard et al., 2017; Zoppi et al., 2018; Farnaby et al., 2019). PROTAC-based inhibitors against SMARCA4, SMARCA2, and PBRM1 exert genomic effects by disrupting SWI/SNF interactions with chromatin (Singh et al., 2007; Filippakopoulos et al., 2012). The relevant example for this class is AU-15330, which degrades by linking the bromodomain ligand of BRG1/BRM/PBRM1 to the ligand of von Hippel-Lindau (VHL) E3 ubiquitin ligase. AU15330 efficiently targets nuclear receptor-positive cancers, such as AR-dependent prostate cancers, by reducing DNA accessibility at enhancer elements, disrupting enhancer-promoter loops, and ultimately suppressing oncogenic gene expression (Xiao et al., 2022).
Indirect targeting approaches
4.1.2
Due to the complexities of direct targeting of SWI/SNF components, sometimes indirect targeting is a valid option. Indirect targeting therapies take advantage of mutations already present in cancer cells, rendering them sensitive to the inhibition of other proteins through a mechanism known as synthetic lethality. Cancers with loss-of-function mutations in tumor suppressors may benefit more from indirect targeted therapies, where synthetic lethal interactions can be exploited (Figure 5B) (Nijman, 2011; Wanior et al., 2021). The most common example of these interactions is provided by the combination of mutually exclusive gene paralogues, such as in cancers with ARID1A mutation that require ARID1B for survival or tumors with BRG1 deficiency, which are sensitive to SMARCA2 depletion (Kaelin, 2009; Helming et al., 2014; Hoffman et al., 2014). Hence, selective degradation of the SWI/SNF ATPase SMARCA2 by PROTACs YDR1 or YD54 was found to provide better sensitivity in SMARCA4 mutant xenografted lung cancer cells (Kotagiri et al., 2025). Other known synthetic lethal interactions are BRD9/SMARCB1, SMARCC1/SMARCC2, BRG1/ACTB, BRG1/ARID2 and BRG1/BRM/ARID1A with p400 complex (Figure 5B) (Michel et al., 2018; Schick et al., 2019; Martin et al., 2023). Synthetic lethality can be induced in several ways, such as by inhibiting the DNA damage repair process in case of SWI/SNF mutant tumors. For example, ARID1A-deficient cancers respond to PARP inhibitors due to ARID1A’s role in DNA repair; ATR/ATM inhibitors are synergistic in cancers with ARID1A-deficiency or BAF-inhibition, and cancers with PBRM1, BRD7, or BRD9 inactivation are sensitive to replication stress and can be treated with PARP and ATR inhibitors (Shen et al., 2015; Williamson et al., 2016; Hu et al., 2019; Chory et al., 2020; Zhou et al., 2020; Chabanon et al., 2021). A similar response can also be seen by targeting proliferation-related factors. For instance, CDK4/6 inhibitors can sensitise BRG1-defective cancer cells, especially in small cell carcinoma of the ovary hypercalcemic type (SCCOHT) and Non-small cell lung cancer (NSCLC) due to the downregulation of cyclin D1 (Xue et al., 2019a; Xue et al., 2019b). Similarly, Aurora Kinase A (AURKA) promotes cell cycle progression and can be targeted in ARID1A-deficient colorectal cancer cells (Wu et al., 2018).
Other indirect targeted therapies include epigenetic therapies that target post-translational modifications of histones and immunotherapy-based approaches such as immune checkpoint blockade therapies. Epigenetic drugs like histone deacetylase (HDAC), DNA methyltransferase (DNMT), and enhancer of zest homolog 2 (EZH2) inhibitors are effective in SCCOHT with BRG1 loss-of-function, while pan-HDAC inhibitors are studied for their efficacy in treating *ARID1A-*mutated tumors (Fukumoto et al., 2018; Auguste et al., 2020). Mutations in ARID1A/ARID1B/ARID2 render cancers more prone to immune checkpoint blockade therapies (Zhu et al., 2022). The combination of HDAC6 inhibitor and anti-PD-L1 therapy in ARID1A-mutated ovarian cancers showed promising antitumor effects due to improved cytotoxic T-cell activity (Fukumoto et al., 2019). Moreover, PBRM1, which plays a role in anti-tumor immune response, might be a valuable and prospective therapy target.
Despite the tumour suppressive role of ARID1A and loss of BRG1 activity in ARID1A-perturbed ER+ breast cancers (Nagarajan et al., 2020), BRG1 and BRM may present as efficient targets in other subtypes due to their crucial roles in promoting progression and mediating resistance to chemotherapy. Previous research shows that the knockout of BRG1 increases the sensitivity to chemotherapy drugs such as 5-FU and paclitaxel (Wu et al., 2016b). BRG1 depletion reduces the gene expression of a drug transporter, ATP-binding cassette (ABC). This eliminates transporter induction, leading to an increase in intracellular drug concentration and sensitivity to chemotherapy (Dubey et al., 2016; Wu et al., 2016b; Wu et al., 2017). Coherently, the inhibition of drug efflux due to the absence of BRG1 also decreases the ability of the cancer cells to eliminate DNA damage (Li K. et al., 2024). This highlights the importance of inhibiting BRG1 and BRM, particularly in cases such as TNBCs, which often rely on nonspecific cytotoxic drugs (Sobczak et al., 2020).
Furthermore, utilising SWI/SNF-based synthetic lethal approaches presents a severe drawback in tumours where other complex subunits present with tumor suppressive function (Narlikar et al., 2013; Zinzalla, 2016). More careful approaches are crucial to selectively target oncogenic function while leaving tumor suppression function unaffected in the SWI/SNF complex. These can include narrowing the therapeutic window or inhibiting specific protein interactions (Zinzalla, 2016). Either the intra-complex protein-protein interactions (PPI) with cytoplasmic signaling effectors or transcription factors can be inhibited. For example, the manipulation of the Armadillo repeats (ARM) domain of the ARID1B subunit may disrupt ARID1B interaction with other subunits in the SWI/SNF complex and induce synthetic lethality in ARID1A-deficient cancers (Takada et al., 2012; Yu et al., 2013). Another approach may involve inhibiting the interaction with oncogenic transcription factors (MYC, JUNB, Notch1, beta-catenin, STAT1/2/3, SMAD2/3, NF-kB, and TCF3) or tumor suppressors (pRB, p53, and BRCA1) (Zinzalla, 2016). On the other hand, identifying the exact subunit that mediates the interaction with a specific transcription factor provides another opportunity, but only a handful have been identified. ARID1A possesses the interacting protein motifs (LXXLL), which enable binding to nuclear hormone receptors. SMARCD1 and SMARCD3 have been shown to interact with transcription factors, such as p53, NF-kB, and SALL4. Hence, developing selective inhibitors to target these interactions is of great interest (Puri and Mercola, 2012; Zinzalla, 2016).
Targeting other chromatin remodelers
4.2
In addition to the SWI/SNF subfamily, bromodomains of the ISWI subfamilies can be targeted (Pérez-Salvia and Esteller, 2017). For example, by targeting the bromodomain, a small molecule inhibitor NVS-CECR2-1 can affect CECR2 binding to chromatin and induce apoptosis in several cancer cell lines (Pérez-Salvia and Esteller, 2017; Park et al., 2020). Considering the role of CECR2 in modulating the immune response in the breast cancer tumor microenvironment and promoting metastasis, utilizing this agent to achieve an immune-responsive tumor microenvironment might be a promising approach to counter metastatic breast cancer with upregulated CECR2. Since BPTF is known to be frequently altered in breast cancer, inhibition of the BPTF bromodomain by aryl urea-1 (AU1) is a promising strategy to target the NURF1/5 remodeling complex (Frey et al., 2017). AU1 causes G1 arrest by impeding chromatin accessibility, lowering c-MYC occupancy, and subsequently diminishing the proliferative capacity of cancer cells. Another prospective BPTF inhibitor is sanguinarine chloride, which showed a potent antiproliferative effect in pancreatic cell lines by downregulating c-Myc expression (Liang et al., 2023). Although BPTF is known for its extensive role in regulating immunity in tumor microenvironments, the extent to which AU1 inhibition affects the BPTF immune function remains to be elucidated. Another selective competitive inhibitor that targets BAZ2A/B bromodomains is GSK2801. While it has no significant tumor suppressive effect as a single agent, GSK2801 showed a strong synergistic activity along with the BET inhibitor JQ1 in several TNBC cell lines by targeting BRD2 (Bromodomain-containing protein 2), which is co-regulated along with BAZ2A/B (Bevill et al., 2019). GSK2801 displaces BRD2 at the promoters of genes regulated by ETS (E26 transformation-specific) transcription factors and at 45S ribosomal DNA promoters, which induces caspase-3 activity and PARP cleavage, leading to apoptosis of tumor cells in the 3D cultures (Bevill et al., 2019). Although the mutation frequency of BAZ2A/B is insignificant in breast cancer, this instance suggests the possibility of targeting the chromatin remodeling complexes with synergistic inhibitors to overcome the limitation of single-agent therapies.
Despite its significant alteration rate and role as a driver gene for breast cancers, CHRAC1 does not possess a bromodomain and presents a significant challenge in developing small-molecule inhibitors. However, CHRAC1 is known to potentially interact with YAP in breast cancer and exerts its tumorigenesis effect by enhancing YAP target oncogenes, which are involved in the Hippo pathway (Li S. et al., 2024). Hence, targeting the interaction between CHRAC1 and YAP can provide an essential and novel approach to breast cancer treatment strategy. RSF1 is also faced with a similar issue due to the lack of bromodomain or catalytic domains, and further study on RSF1 interaction partners in breast cancers may potentially give therapeutic benefits.
Modulating the post-translational modifications is the most feasible approach for targeting CHD subfamilies, as no specific inhibitors have been discovered to date. Synthetic lethality between CHD1 and PTEN can also be employed, as seen in the PTEN-deficient xenograft model (Zhao et al., 2017). Since members of the CHD subfamily usually compose or interact with larger protein complexes, inhibiting other vital interactors, i.e., HDAC inhibitors, can also disrupt the function of CHD remodelers (Xue et al., 1998; Low et al., 2016; Smith et al., 2018). Moreover, CHD3 and CHD4 are known to participate in DNA repair, and the recruitment is mediated by PARP activity (Smith et al., 2018). Hence, PARP inhibitor AG-014699 and the suberohydroxamic acid as an HDAC inhibitor have been shown to inhibit CHD4 function and tumor growth in a mouse xenograft model of EpCAM^+^ hepatocellular carcinoma (Nio et al., 2015). Lastly, E3 ubiquitin ligases, such as SCFβ-TrCP E3 ligase, which targets CHD1 and FBXW7 E3 ligase for CHD6 protein, are being developed (Zhao et al., 2017; Zhang B. et al., 2022). Development of E3 ligases for CHD7 degradation may be beneficial for breast cancer treatment (Stark et al., 2006; Wagner et al., 2011; Beltrao et al., 2012; Povlsen et al., 2012; Akimov et al., 2018; Ordureau et al., 2020; Pedersen et al., 2021).
Despite being a potential target in many cancers, inhibition of INO80 complex subunits is poorly investigated. There are very few studies on inhibiting the INO80 and SRCAP complexes, for example, using Inositol hexaphosphate (IP6) against the INO80 complex (Shen et al., 2003; Willhoft et al., 2016).
Discussion
5
Direct targeting strategies
5.1
While the direct targeted therapies offer precision and selectivity in inhibiting the proteins of interest, the complexity of the chromatin remodeling complexes may raise problems with this approach. Furthermore, the specificity of direct targeted therapies in inhibiting specific proteins may also limit their broader applicability. Patients with mutations or aberrations on the target genes that provide synthetic lethality may benefit from the above therapies, however, identifying the aberrations occurring in breast cancer patients before initiating therapies is laborious and poses financial burdens for clinical settings.
Another pitfall is the capability of these therapies to achieve context-dependent effects in different cancers. Intratumoral heterogeneity coerces more complexity by establishing different molecular subtypes; for example, in small cell lung cancers (SCLC), these signatures are driven by transcription factor, which include achaete-scute family bHLH transcription factor 1 (ASCL1), neurogenic differentiation factor 1 (NeuroD1), POU domain class 2 transcription factor 3 (POU2F3), and yes-associated protein 1 (YAP1) (Borges et al., 1997; Huang et al., 2018; Rudin et al., 2019; Duplaquet et al., 2024; He et al., 2024). Among these subtypes, POU2F3-driven SCLC (SCLC-P) exhibits SWI/SNF dependency, especially towards SWI/SNF ATPases and ncBAF complex subunits, including BRG1, SMARCD1, and BRD9 (Huang et al., 2018; Duplaquet et al., 2024; He et al., 2024). This is evidenced by strong effects on proliferation and expression of POU2F3 and its coactivators upon BRD9 degradation (using FHD-609, WA-68-VQ71, or dBRD9A) on non-neuroendocrine POU2F3+ SCLC models, and pan-SWI/SNF ATPase inhibition or degradation (using FHD-286, BRM014 and AU-15330) on all POU2F3+ cells (Figure 5A) (Duplaquet et al., 2024). Further analyses showed that the reduction in POU2F3 levels is due to the impairment of the chromatin accessibility of the POU2F3 gene. This dependency serves as the basis for utilizing the SWI/SNF ATPase inhibitor as a therapeutic agent for SCLC-P. However, other subtypes of SCLC are resistant to SWI/SNF inhibition, underscoring the importance of molecular subtype identification in cancers with diverse intratumoral heterogeneity and context-specific association of transcription factors with epigenetic modulators (Huang et al., 2018; Duplaquet et al., 2024).
While PROTACs like AU15330 provide high selectivity over ATPase subunits of SWI/SNF complex, they may not provide a therapeutic advantage in cancers where BRG1 and ARID1A are tumor suppressors, as BRG1/BRM degradation may further compromise their tumor-suppressive ability and jeopardize the cancer cells by developing other mutations due to deregulated genomic instability. Moreover, several breast cancer cell lines, such as MCF7, MDA-MB-486, and MDA-MB-231, exhibit resistance to AU15330 treatment (Xiao et al., 2022). These issues may be associated with subtype heterogeneity and genetic diversity in cancer cell line models (Navin et al., 2010; Shah et al., 2012; Yang et al., 2018). Further studies exploring subtype-specific vulnerability against SWI/SNF inhibitor agents in various breast cancer in vivo models may give a better understanding of context-specific effects in breast cancer treatment. To aid this strategy, an advanced molecular profiling with analyses of differential gene expression and chromatin profiling can be employed to identify whether BRG1 or any other targeted proteins of interest predominantly support oncogenic or tumor-suppressing pathways. If it primarily supports oncogenic pathways, it is safe to assume that inhibiting BRG1 would help suppress breast cancer progression. Otherwise, the inhibition of the BRG1 might be deleterious, unless the patients are presented with BRG1 or ARID1A/ARID1B mutations, which require careful monitoring of gene alterations. Moreover, due to the broad expression of SWI/SNF subunits in cells, adverse side effects such as differentiation syndrome, hyperbilirubinemia, etc., were observed in patients treated with FHD-286 in phase 1 clinical trials (Dinardo et al., 2025). Hence, localizing the drug administration to cancerous tissue and minimizing exposure to normal cells by utilizing targeted delivery systems, such as with nanoparticles or antibody-based systems, can aid in reducing the adverse side effects. Combination with chemotherapeutic agents would also provide a better outcome by making the proliferating cells more vulnerable to these targeted therapies.
In addition, inadequate pharmacological properties for clinical use may also cause concerns. For example, AU24118 is more beneficial than AU15330 in treating SCLC-P xenografts NCI-H1048 and NCI-H211 due to its oral bioavailability and enhanced pharmacokinetic properties (He et al., 2024). AU24118 also downregulates POU2F3 and its coactivator, resulting in the reduction of tumor volume and weight. Other agents, such as FHD-609 and FHD-286, which act as BRD9 degraders and BRG1 inhibitors, respectively, have shown a promising increase in survival rate after 35 days of continuous treatment in the same SCLC-P xenografts (Duplaquet et al., 2024). Furthermore, these agents can be used with conventional chemotherapies for SCLC, such as cisplatin and etoposide, and have shown additive and synergistic anti-tumor effects without posing toxicity in multiple SCLC-P models (Duplaquet et al., 2024; He et al., 2024). Hence, exploring the efficacy of these improved clinical-grade inhibitors across different models and molecular contexts is of interest in addressing the intratumoral heterogeneity and resistance mechanisms in breast cancers.
While the premise of blocking BRG1 and BRM might seem prospective, ATPase-binding sites are very dynamic since they modulate intermolecular interactions during the remodeling cycle. The high conservation of ATPase domains and allosteric modulation of the pocket conformation by protein binding partners complicates drug design (Racki et al., 2014). While PROTAC is a beneficial strategy, the disruption of SWI/SNF subcomplexes through bromodomain degradation may create residual complexes without catalytic activity. These residual complexes can still activate deregulated transcription factor networks and drive oncogenic gene expression (Pan et al., 2019; Cantley et al., 2022). Lastly, the discovery of BD98, a macrolactam inhibitor, mimics the effect of ARID1A/B loss (Chory et al., 2020; Dreier et al., 2024). This agent can be best combined with ATR inhibitors for breast cancers, which can improve the efficacy of adaptive T-cell therapy by preventing differentiation (Guo et al., 2022). However, there are limited studies on this agent, and inhibitors for ATPase domains and bromodomains are still the most common approach (Dreier et al., 2024).
Despite their feasibility and potential, several challenges related to prolonged SWI/SNF inhibitor exposure demand further consideration for clinical use. This includes exploring the side effects of long-term administration, the chance of predisposing patients to secondary malignancies, the impact on SWI/SNF-maintained adult stem cells, and investigating the efficacy of combination therapies. Moreover, studies on potential drug resistance issues and biomarkers for monitoring and predicting prognosis still need to be explored (Dreier et al., 2024).
Indirect targeting strategies
5.2
While indirect strategies offer a more effective approach due to exemplar and well-studied synthetic lethality approaches that sensitise the cancer cells, context-specific effects must also be considered. For example, HDAC inhibition might not be a favorable therapeutic option for ARID1A-mutated ER+ breast cancers, as ARID1A-loss-induced endocrine resistance is mediated by the loss of HDAC1 activity (Nagarajan et al., 2020). Interestingly, the same study showed that the hyperacetylation and reprogrammed binding of BRD4 due to HDAC1 loss in these models can be manipulated as a therapeutic strategy using BET/HAT inhibitors. Unfortunately, BET inhibitors are not selective to BRD4 alone, and the pharmacokinetic profiles of many BET inhibitors failed tremendously in the initial phases of clinical trials due to dose-limiting toxicities such as thrombocytopenia (Doroshow et al., 2017; Sun et al., 2021; Trojer, 2022). Other strategies, such as controlled dosage of selective BRD4 targeting using PROTACs in combination with chemo- or endocrine therapies, may provide some potential benefits. Furthermore, novel vulnerabilities that align with the defined genetic defects need to be identified by applying genomic screens in an unbiased manner.
Although inhibiting the interactions of lineage-specific transcription factors with chromatin remodelers can provide ideal and specific targeting against cancers, it is significantly more difficult due to the undruggable nature of IDR regions, which interact with transcription factors (Sammak and Zinzalla, 2015). Hence, developing alternative strategies like interfering peptides against IDRs may present promising therapeutic approaches for treating breast cancers (Patil et al., 2023; Xie et al., 2025).
Conclusion and future perspectives
6
All subfamilies of chromatin remodeling complexes influence and regulate breast cancer aggressiveness to different extents. While there is a growing body of literature supporting their role in cancer, further studies using genetic and molecular profiling of breast tumours can delineate their tumor suppressor or coactivator activity and context-specific functions. This can reveal new avenues for developing effective therapeutic targets for breast cancer. Targeting subunits or other complexes that show synthetic lethal interactions may be feasible, which can tailor the treatment to deregulated cancer cells, overcome adverse side effects, and target resistance and relapse in breast cancer. In addition, the direct or indirect therapies targeting chromatin remodeling complexes should be tested in combination with existing systemic and immune checkpoint therapies, as this strategy might provide synergistic actions and potential benefits to cancers with genetic alterations on these complex subunits. Further exploration of transcription factor-specific associations is warranted to develop therapies in a subtype-specific manner and to establish precision medicine in relevance to complex-specific mutations. Research avenues must also explore the possibility of exploiting the chromatin remodeling complexes as measurable endpoint biomarkers for predicting therapeutic response and prognosis in breast cancer.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abbey M. Trush V. Gibson E. Vedadi M. (2018). Targeting human retinoblastoma binding protein 4 (RBBP 4) and 7 (RBBP 7). bio Rxiv. 10.1101/303537 · doi ↗
- 2Afifi N. Barrero C. A. (2023). Understanding breast cancer aggressiveness and its implications in diagnosis and treatment. MDPI 12 (4), 1375.10.3390/jcm 12041375 PMC 995937236835911 · doi ↗ · pubmed ↗
- 3Akimov V. Barrio-Hernandez I. Hansen S. V. F. Hallenborg P. Pedersen A. K. Bekker-Jensen D. B. (2018). Ubi Site approach for comprehensive mapping of lysine and N-terminal ubiquitination sites. Nat. Struct. Mol. Biol. 25 (7), 631–640. 10.1038/s 41594-018-0084-y 29967540 · doi ↗ · pubmed ↗
- 4Allen H. F. Wade P. A. Kutateladze T. G. (2013). The Nu RD architecture. Cell Mol. Life Sci. 70 (19), 3513–3524. 10.1007/s 00018-012-1256-2 23340908 PMC 3652912 · doi ↗ · pubmed ↗
- 5Allo G. Bernardini M. Q. Wu R.-C. Shih I.-M. Kalloger S. Pollett A. (2014). ARID 1A loss correlates with mismatch repair deficiency and intact p 53 expression in high-grade endometrial carcinomas. Mod. Pathol. 27 (2), 255–261. 10.1038/modpathol.2013.144 23887303 PMC 4603563 · doi ↗ · pubmed ↗
- 6Alver B. H. Kim K. H. Lu P. Wang X. Manchester H. E. Wang W. (2017). The SWI/SNF chromatin remodelling complex is required for maintenance of lineage specific enhancers. Nat. Commun. 8 (1), 14648. 10.1038/ncomms 14648 28262751 PMC 5343482 · doi ↗ · pubmed ↗
- 7Arnold M. Morgan E. Rumgay H. Mafra A. Singh D. Laversanne M. (2022). Current and future burden of breast cancer: global statistics for 2020 and 2040. Breast 66, 15–23. 10.1016/j.breast.2022.08.010 36084384 PMC 9465273 · doi ↗ · pubmed ↗
- 8Auger A. Galarneau L. Altaf M. Nourani A. Doyon Y. Utley R. T. (2008). Eaf 1 is the platform for Nu A 4 molecular assembly that evolutionarily links chromatin acetylation to ATP-dependent exchange of histone H 2A variants. Mol. Cell. Biol. 28 (7), 2257–2270. 10.1128/MCB.01755-07 18212047 PMC 2268442 · doi ↗ · pubmed ↗